








doi: 10.15389/agrobiology.2012.5.64eng
УДК 633.49:632/3:631.522/.524:577.2
INTERSPECIFIC AND INTRASPECIFIC DIVERSITY ON RESISTANCE TO Y VIRUS IN POTATO
E.V. Rogozina, O.Yu. Shuvalov, O.Yu. Antonova, T.A. Gavrilenko
The specimens of cultivated and wild species of potato from collection of N.I. Vavilov Institute of Plant Industry (VIR) were estimated on their resistance to Y virus with the use of phytopathological and molecular-genetic methods. The authors revealed the inter- and intraspecific variability on their resistance to Y virus and also the diversity of plant’s response in cultivated and wild species of potato on viral infection. Several samples of potato genotypes with available diagnostic components RYSC3-320 and GP122-EcoRV-406 are selected and characterized on their resistance to potato virus Y, which are the source of forms, valuable for breeding.
Keywords: potatoes, wild and cultivated species, potato virus Y, DNA markers.
Immunity of cultural plants is shown by many works of N.I. Vavilov's scientific heritage. He studied various infectious diseases in “a wide range of plant objects”, and considered that “the nature of viruses – causes of infection – is quite specific and distinct from other parasites” (1). The 1930ies were the earliest years of virology science. Tobacco mosaic virus, or TMV, was a new type of plant pathogens discovered at the end of XIX century (2), potato leaf roll virus (PLRV) and Y-virus (PVY) – in early XX century (3, 4).
Vavilov’s article “Plant Immunity to Viral Diseases” (published 1937) was based upon the study of the world plant collection of VIR (All-Russian Research Institute of Plant Industry) and available reports of foreign researchers. This work highlights frequent occurrence of viral diseases in vegetatively propagated crops, geographical regularities in the severity of plant infestation, and the influence of external conditions on symptoms of viral diseases. Viral diseases of plants were assumed to have one major feature – “the absence of any connections between phylogenic level of a host plant and its response to damaging by a virus”, which contrasted to known facts about parasitic fungi. In the above-cited work N.I. Vavilov also noted that the damage from viral disease had become a serious problem in the maintenance of a large variety of wild and cultivated potato species collected in the Cordillera; he wrote that the most distant phylogenetic forms and types of potatoes were susceptible to viruses, and many cultivated varieties were affected by the most common mosaic virus (1).
Potato virus Y, or PVY, is one of the most important plant viruses affecting potato; in epiphitotic years, it can cause the loss of about 80% yield of potato (5-7).
In this work the authors continued to study VIR collection of potato in respect to PVY resistance, particularly inter- and intraspecific diversity of this trait in a number of cultivated and wild potato forms by means of phytopathological and molecular genetic methods.
Technique. The material for study were samples of wild and cultivated potato species from VIR collection related to the North American group (series Demissa, Pinnatisecta, Longipedicellata) and the South American group (series Tuberosa, Maglia, Acaulia, Megistacroloba). Totally there were examined 416 collection samples, including 309 samples of cultivated species: Solanum ajanhurii — 8, S. chaucha — 27, S. curtilobum — 10, S. juzepczukii — 13, S. phureja — 44, S. stenotomum — 74, S. tuberosumsubsp. tuberosum — 38, and S. tuberosum subsp. andigenum — 95, as well as 107 samples of wild species: S. acaule — 12, S. albicans — 1, S. brevicaule — 5, S. bukasovii — 2, S. canasense — 1, S. demissum — 8, S. gourlayi — 12, S. hondelmannii — 7, S. leptophyes — 4, S. maglia — 3, S. megistacrolobum — 2, S. multidissectum — 7, S. oplocense — 5, S. pinnatisectum — 6, S. sparsipilum — 7, S. spegazzinii — 13, S. stoloniferum — 8, S. tarnii — 1, and S. vernei — 3. The names of series and species were adopted from J. Hawkes (J. Hawkes, 1990). The material was presented by individual genotypes-clones (3-5 plants of each genotype) or seedlings (20-30 plants per sample).
The resistance to PVY was determined in the field conditions against a high infectious background for all samples of each group. The samples not affected by PVY in the field were further subject to artificial infection (mechanical inoculation and grafting) (8). The results were assessed visually by observation of symptoms of the disease, and by enzyme immunoassay (ELISA-test) with diagnostic kits (“Agdia Inc.”, USA). Causes of infection - the strains PVYo and PVYn – were provided by potato plants of varieties, respectively, Detskoselsky and Wilga (IHAR - The Plant Breeding and Acclimatization Institute, Radzików, Poland). Pathogenicity of these isolates was pre-tested on test-plants Nicotiana tabacum (cv Samsun). Clonal plants were tested in a 2-fold repetition by inoculation of 1-8 shoots (3 leaves per shoot were inoculated). The plants not infected through the mechanical inoculation were infected repeatedly by grafting on infected tobacco or tomato plants.
Molecular screening was performed using specific markers developed for MAS (marker-assisted selection) of PVY-resistant potato varieties. These markers are linked to genes providing extreme resistance to PVY: CAPS-marker GP122/EcoRV-406 for Ry-fsto gene (12th chromosome) (9, 10) and SCAR-marker RYSC3-320 for Ruadg gene (11th chromosome) (13, 14). The conditions of PCR and restriction corresponded to those recommended by developers of the primers (10-12). The products of amplification and restriction were separated by electrophoresis in horizontal 1,6% agarose gels in TBE buffer, stained with ethidium bromide and visualized under UV light.
Results. All tested potato forms expressed inter- and intraspecific variation of susceptibility to PVY in the conditions of high infectious background (Table 1). PVY infestation was quite frequent in cultivated species, much more in polyploid (89-93% samples affected) rather than in diploid S. phureja and S. stenotomum (23 and 50%). Many wild samples were affected by PVY as well: among the South American species of brevicaule - 26%, Acaulia - 67%, and in phylogenetically distant North American species the series Longipedicellata and Pinnatisecta – respectively, 37 and 67% (Table 1).
Various types of response to the infestation were manifested by the studied samples – resistance (or field resistance), tolerance to infection, hypersensitivity, and immunity (i.e. maximum degree of resistance).
| 1. Results of phytopathological study and molecular screening of cultivated and wild potato (genus Solanum L.) forms from VIR plant collection in respect to PVY virus infestation | ||||
Series, species |
Number of studied samples |
|||
total |
ELISA-tested / |
with diagnostic fragment/ |
||
RYSC3-320 |
GP122-EcoRV-406 |
|||
Species from South America |
||||
Cultivated |
||||
Series Tuberosa: |
|
|
|
|
S. chaucha |
27 |
14/13 (93) |
0 |
14/5 |
S. curtilobum |
10 |
10/9 (90) |
0 |
0 |
S. phureja |
44 |
31/7 (23) |
0 |
0 |
S. stenotomum |
74 |
55/25 (50) |
0 |
7/0 |
S. tuberosumsubsp. andigenum |
95 |
95/85 (89) |
2/0 |
6/2 |
S. tuberosumsubsp. tuberosum |
38 |
38/35 (92) |
0 |
0 |
Wild |
||||
Series Tuberosa: |
|
|
|
|
|
66 |
35/9 (26) |
0 |
12/0 |
Series Acaulia: |
|
|
|
|
S. acaule |
12 |
12/8 (67) |
0 |
0 |
Species from North America |
||||
Wild |
||||
Series Demissa: |
|
|
|
|
S. demisssum |
8 |
– |
6/– |
0 |
Series Longipedicellata: |
|
|
|
|
S. stoloniferum |
8 |
8/3 (37) |
5/0 |
3/0 |
Series Pinnatisecta: |
|
|
|
|
S. pinnatisectum |
6 |
6/4 (67) |
6/4 |
0 |
Note.ELISA was performed on plants without visible symptoms of PVYlesion; the table shows data of species represented by five and more samples. Dashes mean that the material wasn’t investigated. |
||||
Among the South American cultivated species (S. phureja, S. stenotomum, S. tuberosum subsp. andigenum, S. tuberosum subsp. tuberosum, S. curtilobum) and wild species (S. brevicaule, S. bukasovii, S. gourlayi, S . hondelmannii, S. leptophyes, S. multidissectum, S. sparsipilum, S. spegazzinii, S. albicans, S. acaule) field-resistant genotypes occurred with different rates. Several field-resistant genotypes of cultivated diploids S. phureja, S. stenotomum were not affected by the virus even through the mechanical inoculation. Among the North American wild species there were revealed field-resistant genotypes (samples S. stoloniferum and S. pinnatisectum), hypersensitive (S. stoloniferum), and immune (samples S. pinnatisectum, S. tarnii, S . stoloniferum).
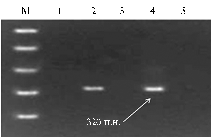
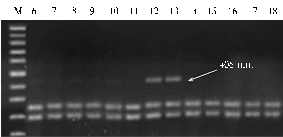
The data of phytopathological evaluation were compared with results of molecular screening by ELISA (Table 1, Fig., A, B). Among all studied South American species the diagnostic fragment RYSC3-320 was detected in only two samples (K-1742, K-15826) of tetraploid cultivated potato from Andes (S. tuberosum subsp. Andigenum), and among the samples of all studied North American species (S. pinnatisectum, S. tarnii, S. stoloniferum, S. demissum). Diagnostic fragment GP122-EcoRV-406 was detected in several genotypes of cultivated species S. stenotomum, S. chaucha, S. tuberosum subsp. andigenum, in wild South American species the group brevicaule, S. megistacrolobum, and in Mexican species S. stoloniferum (Table 1).
Comparison of data obtained by two different methods (phytopathological analysis and molecular screening for presence/absence of a diagnostic fragment) doesn’t give a clear answer about efficiency of the used markers (Table 1.).
| A |
B |
|
|
Electrophoretic spectra of PCR-amplification of markers linked to the genes providing maximum resistance to PYV virus in samples of potato varieties from VIR plant collection: A — SCAR-marker RYSC3-320 for gene Rуadg, B — CAPS-marker GP122/EcoRV-406 for gene Ry-fsto; 1 — S. tuberosumsubsp. andigenum (K-4500), 2 — S. tarnii (GLKS), 3 — S. chaucha(K-24688), 4 — S. demissum (K-23321), 5— S. chaucha(K-24678), 6— S. phureja(K-22218), 7 — S. phureja (K-2221), 8— S. curtilobum (K-13698), 9— S. phureja (K-22199), 10— S. phureja (K-21569), 11— S. phureja (K-5962), 12 — S. stoloniferum(K-3326), 13 — S. chaucha (K-24688), 14 — S. chaucha (K-24684), 15 — S. chaucha (K-24683), 16 — S. stenotomum (K-11023), 17 — S. stenotomum (K-11026), 18— S. brevicaule (K-5663); M – molecular weight marker GeneRuler™ 100 bp Plus DNA Ladder (“Fermentas“, Lithuania). |
|
2. Characterization of potato species Solanum stoloniferum and S. pinnatisectum from VIR plant collection in respect to their response to PVY virus infestation and the presence of R-genes for PVY resistance |
||||||
¹ in VIR catalogue |
Number of studied genotypes |
Response to PVY |
Number of genotypes with presence / absence (+/-) of diagnostic fragments |
|||
RYSC3-320 |
GP122-EcoRV-406 |
|||||
+ |
- |
+ |
- |
|||
S.stoloniferum |
||||||
K-3326 |
2 |
Immunity |
2 |
0 |
2 |
0 |
K-3360 |
7 |
Immunity |
7 |
0 |
3 |
4 |
K-3533 |
4 |
Immunity |
4 |
0 |
4 |
0 |
K-3554 |
4 |
Immunity |
3 |
1 |
0 |
4 |
K-21616 |
2 |
Immunity |
2 |
0 |
0 |
2 |
Total |
19 |
|
18 |
1 |
9 |
10 |
S. pinnatisectum |
||||||
K-4455 |
4 |
Immunity |
2 |
2 |
0 |
4 |
K-4459 |
11 |
Immunity |
11 |
0 |
0 |
11 |
1 |
Susceptibility |
1 |
0 |
0 |
1 |
|
K-15254 |
1 |
Field resistance |
1 |
0 |
0 |
1 |
5 |
Susceptibility |
5 |
0 |
0 |
5 |
|
K-19157 |
3 |
Immunity |
3 |
0 |
0 |
3 |
5 |
Susceptibility |
5 |
0 |
0 |
5 |
|
K-21955 |
11 |
Immunity |
10 |
1 |
0 |
11 |
K-23569 |
5 |
Immunity |
4 |
1 |
0 |
5 |
2 |
Susceptibility |
1 |
1 |
0 |
2 |
|
Total |
48 |
|
43 |
5 |
0 |
48 |
It should be noted that both DNA-markers are linked to R-genes controlling the maximum of resistance to PVY (immunity) in potato. Because immune forms have been detected only among the samples of Mexican species S. stoloniferum and S. pinnatisectum, this material was investigated in detail. The infestation was performed twice - artificial infestation by mechanical inoculation and grafting on PVY-infected tobacco plants, which has revealed 19 immune genotypes among the samples K-3326, K-3360, K-3533, K-3554, K-21 616 of S. stoloniferum (Table 2). S. pinnatisectum was represented by the samples K-4455, K-4459, K-19157, K-21955, K-23569, or totally 48 genotypes were tested; results: one – field resistant, 13 susceptible and 34 - immune. The mentioned genotypes were tested by ELISA for the presence of diagnostic components RYSC3-320 and GP122-EcoRV-406 (Table 2).
The marker fragment RYSC3-320 has been detected in 18 of the 19 immune genotypes S. stoloniferum and in 30 of the 34 immune genotypes of S. pinnatisectum (Table 2). Diagnostic component of the marker GP122-EcoRV-406 has been found in 9 of the 19 immune genotypes of S. stoloniferum, and in none of the 34 immune genotypes of S. pinnatisectum (Table 2).
Thus, the authors have studied the resistance to PVY virus in a number of samples of cultivated potato species, in their wild relatives from the South American center of crop origin, and in phylogenetically distant species from the North American center of origin; these studied forms were different in the level of ploidy (di-, tri-, tetra-, penta-, hexaploid), type of ploidy (auto- and allopolyploids), genomic structure (species with A- and B-genome), and reproduction features (self-pollinated and cross-pollinated forms). The studied potato forms manifested inter- and intraspecific variation in resistance to PVY-virus, as well as different responses of plants to viral infection. These findings suggest the absence of any association between PVY-resistance of plants and their taxonomic classification, genomic composition, and geographical origin of the investigated material. Molecular genetic screening of a wide range of studied potato species has confirmed the point of view of N.I.Vavilov (1) that potato forms immune to viral diseases must be “rare”, and immunity of plants to viral diseases has certain specific features.
The literature data show the efficiency of using MAS-markers RYSC3 (13-15) and GP122 (16). However, in this research these markers were used for screening of particular splitting populations, not for a study of wide variety of species in collections. The obtained results show that these markers known to be developed for MAS of PVY-resistance in potato were ineffective for screening of a wide variety of species. For example, the diagnostic fragment RYSC3-320 has been detected in PVY-resistant S. tuberosum subsp. andigenum, in the majority of immune genotypes of S. stoloniferum, but at the same time – in both immune and susceptible genotypes of S. pinnatisectum. Diagnostic fragment GP122 has been identified only in a half of resistant genotypes of S. stoloniferum, though in several susceptible genotypes of cultivated species S. chaucha and S. tuberosum subsp. andigenum as well. Such facts can be explained by recombination in chromosomal regions with genes controlling PVY-immunity and the corresponding markers, or these species can have some resistance genes distinct from Ruadg (11th chromosome) and Ry-fsto (12th chromosome) providing PVY-immunity, as it was reported by some researchers (17, 18).
So, phytopathological and molecular genetic methods were used to study cultivated and wild potato species in respect to inter- and intraspecific variation in the resistance to PVY-virus, as well as various types of plants’ response to PVY infection. This work has an important practical result – the collection of potato genotypes individually estimated by resistance to PVY-virus and the presence of resistance genes associated with the markers GP122-EcoRV-406 and RYSC3-320 for Ry-fsto and Ruadg resistance genes, which can be used as a valuable source of forms for practical breeding.
REFERENCES
1. Vavilov N.I., Immunity of Plants to Infectious Diseases, in Tr. Vses. soveschaniya po izucheniyu ul’tramikrobov i fil’truyuschikhsya virusov (Papers of All-USSR Congress on Ultramicrobes and Filterable Viruses), Moscow-Leningrad, 1937.
2. Levin A.J. and Enquist L.W., History of Virology, in Fields Virology, Fields, B.N., Knipe D.M., and Howley P.M., Eds., Philadelphia, USA, 2007, p. 5.
3. Quanjer H.M., Van der Lek H.A., and Oortwijn Botjes J.G., On the Nature, Mode of Dissemination and Control of Phloem-Necrosis (Leaf-Roll) and Related Disease, Meded. R. Hoog, 1916, vol. 10, pp. 84-138.
4. Smith К.М., On the Composite Nature of Certain Potato Virus Diseases of the Mosaic Group as Revealed by the Use of Plant Indicators and Selective Methods of Transmission, Proc.the Royal Soc. of London, Series B, 1931, vol. 109, pp. 251-267.
5. Shelabina T.A., Ustoychivost’ k virusam rayonirovannykh sortov kartofelya i osobennosti zaschity ikh v Severo-Zapadnom Nechernozem’ye (Resistance of Potato Cultivars to Viruses and Features of Plant Protection in the North-West Region of Russia.), Cand. Sci. Dissertation, Leningrad, 1989.
6. Jeffries С., Potato. FAO/IPGRI Technical Guidelines for the Safe Movement of Germplasm, No 19, Rome: FAO&IPGRI, 1998.
7. Ivanyuk V., Monitoring of Phytopathogenic State in Belorussia, in Fitosanitarnoe ozdorovlenie ekosistemy (Phytosanitary Measures in Ecosystems: Compilation of Sci. Works), St. Petersburg, 2005, pp. 38-40.
8. Volovik A.S., Trofimets L.N., Dolyagin A.B., and Glez V.M., Metodika issledovaniy po zaschite kartofelya ot bolezney, vrediteley, sornyakov i immunitetu (Guidelines on Studies of Potato Diseases, Pests, Weeds and Immunity), Moscow, 1995.
9. Flis B., Hennig J., Strzelczyk-Zyta D. et al., The Ry-sto Gene from Solanum stoloniferum for Extreme Resistance to Potato Virus Y Maps to Potato Chromosome XII and Is Diagnosed by PCR Marker GP 122718 in PVY Resistant Potato Cultivates, Mol. Breed., 2005, vol. 15, pp. 95-101.
10. Valkonen J., Wiegmann K., Hämäläinen J. et al., Evidence for Utility of the Same PCR-Based Markers for Selection of Extreme Resistance to Potato Virus Y Controlled by Rysto of Solanum stoloniferum Derived from Different Sources, Ann. Appl. Biol., 2008, vol. 152, pp. 121-130.
11. Hamalainen J., Watanabe K., Valkonen J., et al., Mapping and Marker-Assisted Selection for a Gene for Extreme Resistance to Potato Virus Y, Theor. Appl. Genet., 1997, vol. 94, pp. 192-197.
12. Kasai K., Morikawa Y., Sorri V., et al., Development of SCAR Markers to the PVY Resistance Gene Ry adg Based on a Common Feature of Plant Disease Resistance Genes, Genome, 2000, vol. 43, pp. 1-8.
13. Whitworth J., Novy R., Hall D., Crosslin J., and Brow C., Characterization of Broad Spectrum Potato Virus Y Resistance in a Solanum tuberosum ssp. andigena-Derived Population and Select Breeding Clones Using Molecular Markers, Grafting, and Field Inoculations, Am. J. Pot. Res., 2009, vol. 86, pp. 286-296.
14. Sagredo B., Mathias M., Barrientos C., et al., Evaluation of a SCAR RYSC3 Marker of the Ryadg Gene to Select Resistant Genotypes to Potato Virus Y (PVY) in the INIA Potato Breeding Program, Chilean Journal of Agricultural Research, 2009, vol. 69, no. 3, pp. 305-315.
15. Andrade C., Pinto C., De Paula Ribeiro S., Peixouto L., and Vilela X., Potato Clones with Multiple Copies of the Ryadg Allele Conferring Resistance to PVY, Crop Breed. Appl. Biotechnol., 2009, vol. 9, pp. 286-292.
16. Heldák J., Bezo M., Štefúno V., and Galliko A., Selection of DNA Markers for Detection of Extreme Resistance to Potato Virus Y in Tetraploid Potato (Solanum tuberosum L.) F1 Progenies, Czech J. Genet. Plant Breed., 2007, vol. 4, pp. 125-134.
17. Brigneti G., Garcia-Mas J., and Baulcombe D., Molecular Mapping of the Potato Virus Y Resistance Gene Rysto in Potato, Theor. Appl. Genet., 1997, vol. 94, pp. 198-203.
18. Song Y., Hepting L., Schweizer G., et al., Mapping of Extreme Resistance to PVY (Rysto) on Chromosome XII Using Anther-Culture-Derived Primary Dihaploid Potato Lines, Theor. Appl. Genet., 2005, vol. 111, pp. 879-887.
N.I. Vavilov Research Institute of Plant Industry, RAAS, |
Received June 26, 2012 |
![]()




