








УДК 633.11:581.14:[581.132+581.116]:58.02
ИНТЕНСИВНОСТЬ ФОТОСИНТЕЗА И ТРАНСПИРАЦИИ В ОНТОГЕНЕЗЕ ЯРОВОЙ ПШЕНИЦЫ В СВЯЗИ С ОСВЕЩЕННОСТЬЮ И ТЕМПЕРАТУРОЙ
В.А. ДАВЫДОВ
В течение периода вегетации измеряли величину интенсивности фотосинтеза (ИФ) и транспирации (ИТ) яровой пшеницы сорта Скала и оценивали значения коэффициентов линейной корреляции между этими показателями, а также между ними и метеорологическими факторами (освещенность и температура воздуха). Наиболее высокая ИФ и ИТ в расчете на 1 г сухой массы отмечена на стадии начала кущения, в расчете на побег — в фазу цветения, когда за светлое время суток побег фиксирует около 360 мг СО2. За вегетационный период главный побег транспирирует примерно 700-800 мл воды и ассимилирует 20-25 г СО2, около 30 % которого овеществляется в биомассе побега и 5-7 % — в зерне. В первую половину вегетации выявлена тесная связь между указанными процессами и факторами среды, к концу вегетации — ос-лабление корреляции. Сходная тенденция обнаружена в паре признаков ИФ—температура воздуха. Для ИТ взаимосвязь c освещенностью и температурой была неоднозначной.
Ключевые слова: Tritiсum aestivum, фотосинтез, транспирация, освещенность, температура, корреляция, продукционный процесс.
Изучение закономерностей накопления биомассы и формирования урожая неизбежно приводит к пониманию, что факторы среды могут существенно влиять на интенсивность и характер физиолого-биохи-мических процессов у растений, в значительной мере определяя конечную продуктивность (1). Зависимость фотосинтеза и транспирации от освещенности и температуры общеизвестна, но в то же время количественные оценки этой корреляции исследованы явно недостаточно.
Зная особенности ассимиляционной деятельности побега пшеницы, можно выявить периоды, существенные для реализации биологической и хозяйственной продуктивности растений.
В представляемой работе была поставлена задача в разные фазы онтогенеза побега пшеницы измерить интенсивность фотосинтеза (ИФ) и транспирации (ИТ), на основании величины коэффициента линейной корреляции оценить эффективность использования лучистой энергии при фиксации СО2 и выявить взаимосвязь между ИФ и ИТ, температурой воздуха и ИТ у целого растения и его органов (флаговый лист и колос), а также определить эффективность фотосинтетической фиксации СО2, динамику нарастания биомассы и водорасходную величину.
Методика. Яровую пшеницу (Triticum aestivum L.) cорта Скала выращивали на полевом массиве площадью 50 м2 на темно-серой лесной почве в лесостепной зоне Иркутской области. Посев проводился по парам без внесения удобрений. Наблюдения выполняли в течение всего периода вегетации. В работе приводятся результаты, приуроченные к фазам кущения, выхода в трубку, цветения и молочной спелости.
Интенсивность фотосинтеза и дыхания определяли на инфракрасном газоанализаторе Инфралит-4 («Juncolor», Германия) при естественной концентрации СО2, используя набор цилиндрических кварцевых камер, которые одевали на побег или отдельный орган. Скорость прохождения воздушного потока через камеру для листа и колоса составляла 40, для целого побега — 100 л/ч. В верхней части камеры находился отградуированный датчик температуры с выходом на самописец. ИТ измеряли в том же потоке воздуха одновременно с ИФ по разнице с контролем при помощи гигрометра Волна-1 М (Россия) с погрешностью не более 10 %, освещенность над посевом — альбедометром М-69 (Россия), сигнал с которого регистрировался самописцем.
В указанные сроки также оценивали величину надземной и подземной части побегов (не менее чем у 20): после измерений ИФ и ИТ побег разделяли на органы, высушивали при температуре 100 °С и взвешивали.
Достоверность результатов оценивали по t-критерию Стьюдента с учетом стандартной ошибки (Sx). Корреляционный анализ проводили с использованием программы Excel 2000 при величине выборки показателей Р = 15.
Результаты. Первые измерения выполняли в начале фазы кущения проростка, когда сухая масса надземной части составляла 37 мг, корней — 12 мг. Как оказалось, величина ИФ к 800 равнялась 40 мг СО2/(г сухой массы · ч). Однако наибольшие значения ИФ отмечали в 1500 при освещенности 80 Вт/м2: показатель немногим превышал 50 мг СО2/(г сухой массы · ч). Затем значение ИФ изменялось в соответствии с освещенностью, и после 2200 прибор начал регистрировать выделение СО2, которое к 2400 составляло около 10 мг/(г сухой массы ·ч).
Аналогично изменялась интенсивность транспирации: если в дополуденное время величина ИТ не превышала 800 мг Н2О/(г сырой массы x ч), то к 1500 (пик интенсивности) ее величина составила 1100 мг Н2О/(г сырой массы x ч). То есть днем в жаркую солнечную погоду объем Н2О, транспирируемой проростком пшеницы за 1 ч, может превышать его собственную массу.
В то же время довольно высокие значения ИФ в расчете на 1 г сухой массы создают впечатление интенсивной ассимиляции СО2, что, однако, не совсем так. В действительности проросток, сухая надземная масса которого в этот период онтогенеза составляла всего 37 мг, за светлое время суток (условно за 16 ч) поглощал углекислоты в 27 раз меньше, чем в расчете на 1 г: общее количество поглощенного СО2 составляло всего около 16 мг при среднесуточном приросте сухой массы проростка 3,5±0,9 мг. Расход воды на транспирацию в указанный период составлял около 2 г в расчете на один проросток за световой день. В пересчете на 1 га посева при среднем числе проростков около 5 млн (2, 3) это примерно 80 кг СО2 и 10 т воды за световой день.
Выполненные измерения показали, что все изучаемые показатели (ИФ, ИТ, освещенность и температура воздуха) имеют сходную динамику, что подтвердилось высокими коэффициентами линейной корреляции. Наиболее тесную связь выявили между ИФ и ИТ проростка (r= 0,97), а также между ИФ и освещенностью (r = 0,86), более слабую — между ИФ побега и температурой воздуха (r = 0,47).
В фазу выхода в трубку сухая масса побега составляла 1284 мг (1108 мг — надземная часть, 176 мг — корни). В этот период также до 1000 происходило довольно быстрое возрастание интенсивности фотосинтеза, которая затем с небольшим спадом сохранялась до 1900 с последующим резким снижением, и уже к 2200 дыхание преобладало над фотосинтезом. Однако нужно отметить, что в этот период по сравнению с фазой кущения ИФ заметно упала, и ее значения в дневное время составили около 20 мг СО2/(г сухой массы · ч). Расчеты показывают, что за световой день побег пшеницы в указанных условиях ассимилировал немногим более 260 мг СО2 при среднесуточном приросте массы около 78,0±9,0 мг. Величина ИТ в расчете на побег существенно возросла, а в расчете на 1 г сырой биомассы снизилась почти в 2 раза по сравнению с таковой в предыдущий период онтогенеза. В фазу выхода в трубку за светлое время суток побег транспирировал примерно 11 г Н2О. Следовательно, в расчете на 1 га посева потребление СО2 и Н2О у растений существенно возросло и составило в эту фазу соответственно 1,5 и 50,0 т за световой день.
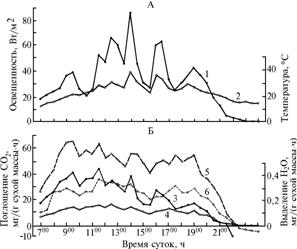 |
Рис. 1. Динамика метеорологических показателей (А), а также интенсивности фотосинтеза и транспирации (Б) в фазу цветения у растений яровой пшеницы сорта Скала: 1 — освещенность, 2 — температура воздуха, 3 — интенсивность транспирации флагового листа, 4-6 — интенсивность фотосинтеза соответственно побега, флагового листа и колоса (Иркутская обл.). |
Корреляция между интенсивностью физиологических процессов и метеорологическими факторами в эту фазу оказалась так-же довольно тесной, причем, например, сопряженность ИФ с температурой стала значительно выше (r = 0,71), чем в фазу кущения (r = 0,47).
В фазу цветения (рис. 1) высокую ИФ наблюдали у флагового листа (его масса равнялась 137 мг, а площадь составляла 80 % от максимальной), который, вероятно, обладал наибольшей ассимиляционной способностью (4, с. 43). В утреннее время ИФ достигала 65,7 мг СО2/(г сухой массы ·ч), или 9,0 мг на лист за 1 ч. Затем до 1900 величина ИФ постепенно снижалась, после 1900 произошло резкое падение и освещенности, и ИФ. Расчеты показали, что за световой день флаговый лист с сухой массой 137 мг ассимилирует около 100 мг СО2.
| Величина коэффициентов линейной корреляции (r) в парах признаков и факторов среды в течение вегетационного периода у растений яровой пшеницы сорта Скала (Иркутская обл.) | ||||
Пара признаков и факторов |
Начало |
Выход в |
Цветение |
Молочная |
ИФ побега—освещенность |
0,86*** |
0,84*** |
0,66*** |
0,40* |
ИФ побега—температура |
0,47** |
0,71*** |
0,54** |
0,07 |
ИФ побега—ИТ побега |
0,97*** |
0,86*** |
|
|
ИТ побега—температура |
0,41* |
0,87*** |
|
|
ИФ флагового листа—освещенность |
|
|
0,56** |
0,50** |
ИФ флагового листа—температура |
|
|
0,43* |
0,02 |
ИФ флагового листа—ИТ флагового листа |
|
|
0,79*** |
0,80*** |
ИТ флагового листа—температура |
|
|
0,24 |
–0,14 |
ИТ флагового листа—освещенность |
|
|
0,55** |
0,30 |
ИФ колоса—освещенность |
|
|
0,64*** |
0,54** |
ИФ колоса—температура |
|
|
0,53** |
0,11 |
ИФ колоса—ИФ флагового листа |
|
|
0,80*** |
0,92*** |
ИФ колоса—ИФ побега |
|
|
0,89*** |
0,94*** |
Освещенность—температура |
0,79*** |
0,53** |
0,82*** |
0,77*** |
П р и м е ч а н и е. ИФ и ИТ — соответственно интенсивность фотосинтеза и транспирации. |
||||
У колоса суточная динамика ИФ была сходной, а максимальная величина фиксации СО2 составила 36,6 мг СО2/(г сухой массы · ч), или 9,7 мг на колос за 1 ч. В эту фазу у колоса, сухая масса которого составляла 267 мг, величина ассимиляции за световой день, как и у флагового листа, была около 100 мг СО2.
Максимальную ИТ флагового листа — 490 мг Н2О/(г сырой массы · ч) — регистрировали около полудня с последующим снижением показателей. Количество Н2О, транспирируемой в расчете на флаговый лист за световой день, составило 5 г.
В эту фазу онтогенеза по сравнению с предыдущим периодом коэффициенты линейной корреляции несколько снизились, особенно в паре признаков ИФ побега—освещенность (r = 0,66), ИФ побега—температура (r = 0,54) и ИФ флагового листа—освещенность (r = 0,56) (см. табл.). В то же время достаточно тесная связь обнаружилась между ИФ и ИТ флагового листа (r = 0,79), ИФ колоса и ИФ флагового листа (r = 0,80), ИФ колоса и ИФ побега (r = 0,89). Слабой оказалась корреляция между ИТ флагового листа и температурой воздуха (r = 0,24), что может свидетельствовать о дефиците воды в листьях. Ассимиляционная активность всего побега оказалась значительной (фиксация 360 мг СО2 за световой день, из которых на долю флагового листа и колоса пришлось примерно 55 %). Величина суммарной транспирации у побега за световой день при солнечной погоде равнялась 13-14 г. В расчете на 1 га посева, учитывая побеги подгона, это составит около 70 т Н2О и не менее 2 т СО2.
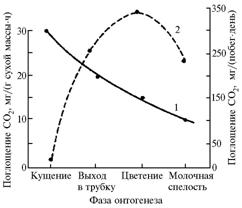 |
Рис. 2. Динамика интенсивности фотосинтеза у яровой пшеницы сорта Скала в разные фазы развития: 1 и 2 — среднедневная величина фиксации СО2 в расчете соответственно на 1 г сухой массы за 1 ч и на побег за световой день (Иркутская обл.). |
Среднесуточный прирост сухой массы растения в эту фазу был максимальным и составил 81,0±14,0 мг. Измерения, выполненные при сплошной облачности, выявили 1,5-2,5-кратное снижение ИФ и ИТ. Многочисленные наблюдения в разные годы на разных сортах показали, что у побега яровой пшеницы в начальной фазе колошения (цветения) наблюдается наибольшая интенсивность ассимиляции СО2, которая в зависимости от погодных условий сохраняется на протяжении 5-10 сут от начала колошения (чем суше и жарче, тем этот период короче), после чего величина ИФ начинает существенно снижаться (рис. 2).
Через 2 нед после цветения, то есть в начале фазы молочной спелости, когда на флаговых листьях некоторых растений проявились первые признаки отмирания, значения ИФ упали: наибольший показатель для флагового листа и колоса, регистрируемый в 1000 и 1300, составлял соответственно 43,5 и 19,6 мг СО2/(г сухой массы · ч). По сравнению с предыдущим периодом онтогенеза суммарная величина транспирации за световой день у флагового листа снизилась в 2 раза и составила 1,5 г. В начале фазы молочной спелости флаговый лист при сухой массе 145 мг в солнечную погоду за светлое время суток ассимилировал 70 мг СО2, колос при сухой массе 520 мг — 80 мг СО2 .
В этот период онтогенеза суммарная фиксация углекислоты всем побегом за светлое время суток составила 250 мг СО2, из которых около 60 % приходилось на флаговый лист и колос, а величина транспирации заметно упала и в хорошую солнечную погоду не превышала 6 г. В пересчете на 1 га посева в сутки количество ассимилируемой углекислоты оказалось примерно таким же, как в фазу выхода в трубку, — 1,5 т СО2, а расход воды значительно снизился и не превышал 30 т Н2О в сутки. Среднесуточный прирост массы побега в этот период составил 67,0±12,0 мг.
Эффективность фотосинтетической деятельности растений определяется отношением количества поглощенного углерода к ассимилирующей массе. Наши расчеты показали, что величина отношения массы ассимилируемого за день СО2 к надземной и общей сухой массе побега в начале фазы кущения была максимальной и равнялась соответственно 0,42 и 0,32, в фазу выхода в трубку составила соответственно 0,23 и 0,19, цветения — 0,16 и 0,14, а в фазу молочной спелости уменьшилась соответственно до 0,08 и 0,07. То есть в расчете на единицу сухой массы в периоды от кущения до начала молочной спелости этот показатель снизился в 4-5 раз.
Специфика поглощения СО2 в период роста и развития побега хорошо иллюстрируется динамикой коэффициентов линейной корреляции между ИФ и освещенностью (см. табл.): в фазу кущения r = 0,86 (наибольшее значение), в фазу выхода в трубку r= 0,84, в начале фазы колошения r = 0,66, в фазу молочной спелости r = 0,40, что, вероятно, объясняется не только снижением содержания хлорофилла в ассимиляционной ткани в течение вегетации (5), но и накоплением гетеротрофной массы клеток. В то же время максимальное значение коэффициента корреляции между ИФ и температурой регистрировали во второй половине июня в фазу выхода в трубку (r = 0,71), в то время как в конце мая в фазу кущения этот показатель был значительно ниже (r = 0,47). Отметим, что в фазу кущения средняя дневная температура воздуха равнялась 20 °С, в фазу выхода в трубку — оказалась примерно на 10 °С выше. Также следует подчеркнуть, что в течение всего периода вегетации величина коэффициента корреляции между ИФ и освещенностью была выше, чем между ИФ и температурой, что может свидетельствовать о более важной роли солнечной энергии и меньшей — температуры в жизнедеятельности растений яровой пшеницы в регионе наблюдений.
Значительно изменился в течение вегетации и транспирационный коэффициент как у побега, так и у отдельного листа. Так, если у проростка в фазу кущения отношение транспирируемой за день массы воды к надземной массе побега равнялось 50, то в фазу выхода в трубку — уменьшилось до 10. Аналогично у флагового листа этот показатель в фазу цветения был равен 15, в фазу молочной спелости — снизился до 10. У побега общий расход воды на транспирацию, как показывают расчеты, за июнь, июль и август составил около 700-800 мл. В расчете на 1 г зерна расход воды составил около 1100 мл, что хорошо согласуется с данными других авторов (6). В пересчете на 1 га производственного посева только главными побегами (5 млн шт.) транспирируется около 4000 т воды, а с учетом побегов кущения — значительно больше.
Подобные расчеты показывают, что за вегетационный период побег пшеницы ассимилирует 20-25 г СО2. Известно (7), что такое количество СО2 эквивалентно 14-17 г глюкозы, около 1/3 которой в конечном итоге овеществляется в биомассе побега и 5-7 % — в зерне. При этом, начиная с фазы цветения, на флаговый лист и колос приходится не менее 50 % всего ассимилированного побегом углерода. С начала колошения эта доля постоянно возрастает, так как нижние листья и стебель к этому времени желтеют и практически не фотосинтезируют. В пересчете на 1 га посева за вегетационный период только главные побеги поглощают 100 т СО2 (с учетом подгона — существенно больше). В настоящее время фотосинтетическую нагрузку на флаговый лист пытаются усилить за счет селекции на увеличение площади его поверхности и продолжительности жизнедеятельности (8, 9).
Итак, в процессе онтогенеза побега пшеницы выявилась тесная связь между фотосинтезом, транспирацией и факторами среды — освещенностью и температурой воздуха. Наибольшие показатели интенсивности фотосинтеза и транспирации в расчете на единицу массы отмечены на стадии кущения. Эффективность использования лучистой энергии при фотосинтезе в этот период развития также максимальная, к концу вегетации она существенно снижается. Наибольшую интенсивность фотосинтеза в расчете на побег регистрировали в начальной фазе колошения, когда в зависимости от погоды в течение 5-10 сут сохраняется самый высокий уровень ассимиляции СО2. В этот период онтогенеза при солнечной погоде за светлое время суток побег пшеницы с сухой надземной массой 2 г ассимилировал около 360 мг СО2. Вероятно, указанный период является ключевым для фиксации СО2 и запасания ассимилятов, часть которых сразу, а часть — в дальнейшем за счет реутилизации используется для роста и налива зерновки. Именно в этот период суточный прирост сухой массы побега был максимальным и составлял около 80 мг. Можно полагать, что если до цветения растение не испытывало значительных стрессовых климатических воздействий, то налив зерна и величина урожая в значительной мере определяются степенью соответствия экологических факторов физиологическим потребностям сорта в начальный период колошения.
Л И Т Е Р А Т У Р А
1. Н и ч и п о р о в и ч А.А. Фотосинтетическая деятельность растений как основа их продуктивности в биосфере и земледелии. В сб.: Фотосинтез и продукционный процесс. М., 1988: 5-28.
2. В о л ы н к и н а О.В. Влияние нормы высева пшеницы и обеспеченность ее азотом на урожай и качество. Зерновое хозяйство, 2004, 2: 26-27.
3. Ш а м с у т д и н о в а К.Г., Ш а й х у т д и н о в Ф.Ш., Г а й н у т д и н о в а Р.М. Влияние норм высева яровой пшеницы сорта Амир на урожайность и качество зерна на серой лесной почве Предкамья. Мат. науч. конф. агроэкономического факультета, посвященной 80-летию Казанской ГСХА «Достижения науки сельскохозяйственному производству». Казань, 2002: 58-61.
4. М о к р о н о с о в А.Т. Онтогенетический аспект фотосинтеза. М., 1981.
5. A r a u s J.L., L a b r a n a X., A r a u s J.L. е.а. Leaf photosynthesis and chloroplast senescence patterns in wheat flag leaves during grainfilling. Photosynthetika, 1991, 25, 1: 33-37.
6. К а л и н и н Н.И. Водопотребление и продуктивность засухоустойчивых сортов пшеницы. Докл. ВАСХНИЛ, 1983, 5: 4-6.
7. К у р с а н о в А.Л. Хлоропласт как датчик ассимилятов. В сб.: Фотосинтез и продукционный процесс. М., 1988: 54-69.
8. К у м а к о в В.А. Физиология формирования урожая яровой пшеницы и проблемы селекции. С.-х. биол., 1995, 5: 3-15.
9. H a f s i M., P f e i f f e r W.H., M o n n e v e u x P. Flag leaf senescence carbon content and isotop discrimination in durum wheat grown under semiarid conditions. Cereal. Res. Commun., 2003, 31(1-2): 161-168.
PHOTOSYNTHESIS AND TRANSPIRATION ACTIVITY IN ONTOGENESIS OF SPRING WHEAT IN CONNECTION WITH LIGHT AND TEMPERATURE
V.A. Davydov
During vegetation period the author determined the value of photosynthesis (PA) and transpiration (TA) activity in spring wheat of the Skala variety and estimated the coefficients of linear correlation between these processes and also between meteorological factors (light and air temperature). The most high PA and TA per 1 g of mass was during beginning of bushing out stage, counting on shoot—blooming phase, when for bright day the shoot fix about 360 mg of CO2. For vegetation period the main shoot transpires about 700-800 ml of water and assimilates 20-25 mg of CO2, about 30 % of which transforms to shoot and 5-7 % to corn biomass. During the first half of the vegetation the author finds close connection between indicated processes and factors of environment, towards the end of the vegetation the correlation was slacken. The similar tendency was revealed for the pair of PA—air temperature. For TA the correlation between light and temperature has a no single meaning.
Key words: spring wheat, photosynthesis, transpiration, illumination, temperature, correlation, productional process.
Сибирский институт физиологии и биохимии |
Поступила в редакцию |




