doi: 10.15389/agrobiology.2022.6.1083rus
УДК 636.2:637.043:577.2
Исследование выполнено при финансовой поддержке Российского научного фонда (проект № 21-76-20046).
ИСПОЛЬЗОВАНИЕ ВЫСОКОПРОИЗВОДИТЕЛЬНОЙ ИНФРАКРАСНОЙ СПЕКТРОМЕТРИИ ПРИ ИЗУЧЕНИИ ПОЛНОГЕНОМНЫХ АССОЦИАЦИЙ С ЖИРНОКИСЛОТНЫМ И КОМПОНЕНТНЫМ СОСТАВОМ МОЛОКА У КОРОВ (Bos taurus)
А.А. СЕРМЯГИН✉, Л.П. ИГНАТЬЕВА, И.А. ЛАШНЕВА, А.А. КОСИЦИН, О.В. КОСИЦИНА, А.С. АБДЕЛЬМАНОВА,
Н.А. ЗИНОВЬЕВА
Количество жира в молоке коров относится к признакам, наиболее подверженным высокой изменчивости, и зависит как от условий среды (кормление, технология содержания), так и от генетических факторов (порода, генотип). Особый интерес представляет содержание жирных кислот (ЖК) как биомаркера контроля физиологического состояния животных и критерия оценки показателей качества сырого молока, в части его пригодности к переработке (выход сыра, масла и сливок). Соотношение ЖК в молоке по числу атомов углерода, а также длине цепи, степени ее насыщения различается между особями и на популяционном уровне. Поэтому изучение генетической и геномной изменчивости признаков молочной продуктивности для повышения эффективности управления отбором животных остается актуальной задачей. Цель наших исследований заключалась в поиске полногеномных ассоциаций и полиморфизмов в генах, детерминирующих жирнокислотный состав молока, на основе инфракрасной спектрометрии как одного из наиболее быстрых и точных экспресс-методов физико-химического анализа состава молока. Популяционно-генетические параметры и изменчивость содержания жирных кислот в молоке изучали на популяции голштинизированного черно-пестрого и голштинского скота 14 племенных стад из Московской области (2017-2018 годы). Всего для оценки суточных показателей молочной продуктивности использовали 36982 образца. Расчет коэффициентов наследуемости (h2) и корреляции (rg) показателей состава молока коров проводили на основе метода REML (residual maximumlikelihood) с использованием семейства программ BLUPF90. Поиск SNP проводили в выборке коров из экспериментального стада голштинизированного черно-пестрого скота (ПЗ «Ладожский» — филиал ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2020-2021 годы). Фракционный состав молока определяли с помощью автоматического анализатора MilkoScan 7 DC («FOSS», Дания), принцип действия которого основан на экспресс-оценке методом инфракрасной спектроскопии. При индивидуальной оценке была сформирована группа из 144 генотипированных с помощью биочипа Bovine GGP 150K («Neogen», США) коров с полным фенотипическим описанием спектра жирных кислот и компонентов молока. Контроль качества генотипирования (110884 SNP), анализ полногеномных ассоциаций (GWAS, genome-wide association study) и многомерное шкалирование (MDS, multidimensional scaling) выполняли с помощью программы Plink 1.9. Поиск генов по выявленным значимым полиморфизмам проводили в браузере Ensembl по сборке генома крупного рогатого скота Bos taurus UMD 3.1.1 (https://www.ncbi.nlm.nih.gov/assembly/). Аннотацию генов для определения локусов количественных признаков на хромосомах животных осуществляли по международной базе данных Animal QTLdb. Наследуемость показателей жирных кислот молока варьировала от низкой для полиненасыщенных ЖК (h2 = 0,018) до умеренных для средне- (h2 = 0,125), длинноцепочечных (h2 = 0,155)и миристиновой кислоты (h2 = 0,155), мононенасыщенных ЖК (h2 = 0,176) и олеиновой кислоты (h2 = 0,196). Методом многомерного шкалирования оценили генетическую структуру экспериментальной группы животных, где мы наблюдали умеренный размах вариабельности по первой (PC1 = 7,82 %) и по второй (PC2 = 4,65 %) компонентам изменчивости. Для миристиновой и пальмитиновой ЖК выявлены общие кластеры (QTL) на хромосомах BTA5, BTA10, BTA14, BTA18 и BTA27; для стеариновой и олеиновой ЖК (как входящих в группу длинноцепочечные ЖК) показана схожая локализация QTL на хромосомах BTA9, BTA10, BTA11, BTA14, BTA17, BTA18, BTA19, BTA20 и BTA29. Для коротко- и среднецепочечных ЖК обнаружены ассоциации на хромосомах BTA1, BTA5, BTA10, BTA11, BTA14, BTA18, BTA19 и BTA24, для длинноцепочечных ЖК детектированы QTL на BTA6, BTA7, BTA9, BTA10, BTA11, BTA17, BTA18 и BTA29. Для коротко- и среднецепочечных ЖК, насыщенных ЖК, C14:0, C16:0, C18:0 и C18:1 установлены гены, образующие QTL на хромосомах BTA10, BTA11 и BTA14, — CACNA1C, GCH1,ATG14, KCNH5, PRKCE, CTNNA2, CYHR1, VPS28, DGAT1, ZC3H3, RHPN1, TSNARE1. Коротко- и среднецепочечные ЖК, миристиновая и пальмитиновая ЖК, насыщенные ЖК показали связь с полиморфизмами в генах MED12L, EPHB1, GRIN2B, PRMT8, ERC1, PELI2, ARHGAP39, MROH1, MAF1, GSDMD, LY6D. Для длинноцепочечных и мононенасыщенных ЖК, стеариновой и олеиновой ЖК в результате аннотации получены селекционно значимые гены RPS6KA2, CPQ, CPE, FTO, FAT3, LUZP2. Продолжение изучения генетических механизмов наследования содержания жирных кислот и компонентов молока необходимо для формирования стратегии селекции молочного скота с лучшим жирнокислотным профилем и составом компонентов.
Ключевые слова: корова, жирные кислоты, компоненты молока, коэффициент наследуемости, GWAS, SNP, QTL, гены.
Молочный жир обладает наибольшей энергетической ценностью и широким спектром биологической активности. Он необходим для усвоения различных витаминов, токоферолов, фосфатов и других важных нутриенов. В молоке молочный жир находится в виде суспензии, состоящей из мелких жировых шариков размером от 0,1 до 20 мкм. По химическому составу он представляет собой производное спирта глицерина и жирных кислот (ЖК), на долю которых приходится 93-95 % массы жира. Содержание жирных кислот в молоке может значительно изменяться в зависимости от условий кормления животных, сезона года, стадии лактации и других факторов. Жирные кислоты подразделяются на две категории — насыщенные и ненасыщенные. Последние, в свою очередь, делятся на мононенасыщенные и полиненасыщенные. Содержание насыщенных кислот в коровьем молоке колеблется от 50,3 до 73,8 %, ненасыщенных — от 49,3 до 26,2 %. Из ненасыщенных ЖК в молоке больше всего содержится мононенасыщенных и меньше — полиненасыщенных (1).
Жирные кислоты представляют собой органические соединения, которые отличаются количеством атомов углерода и положением, а также количеством в них двойных связей. Коровье молоко в среднем содержит от 3,6 до 4,8% массовой доли жира. В молоко жирные кислоты поступают как свободными, так и в связанной форме глицеридов или других липидов. На триацилглицериды, которые состоят из глицерола и трех жирных кислот, приходится от 96 от 99 % молочного жира, на свободные жирные кислоты —лишь от 0,1 до 0,4 % (2).
Набор жирных кислот различается в зависимости от породы, времени года и применяемых зоотехнологий, немаловажную роль играют условия кормление коров. Состав жира молока изменяется в течение лактации. В ранний период при раздое организм животного преимущественно использует жирные кислоты С16 (пальмитиновая) и С18 (стеариновая) из жирового депо тканей. В течение лактации повышается доля вновь синтезированных (de novo) жирных кислот (С4:0-С14:0), в то время как доля жирных кислот с 17 и более атомами углерода снижается (3).
В составе молочного жира обнаружено около 140 жирных кислот, однако лишь 13 главных ЖК с четным числом атомов углерода (С4:0-C18:3) встречаются в количестве, которое для каждой составляет более 1 %. Остальные кислоты (например, С10:1, C12:1), присутствующие в количестве менее 1 % и в виде следов, относятся к так называемым минорным ЖК (2-4). У минорных ЖК доля в триглицеридах липидов молока составляет для масляной кислоты — 2,0-4,2 %, для капроновой — 1,5-3,0 %, каприловой — 1,0-2,0 %, каприновой — 2,0-3,5 %, деценовой — 0,2-0,4 %, лауриновой — 2,0-4,0 %, миристинолеиновой — 0,6-1,5 %, пальмитолеиновой — 1,5-2,0 %, линолевой — 3,0-5,5 %, линоленовой — до 1,5 %, арахиновой — до 0,3 %, бегеновой — до 0,1 %. У главных ЖК этот показатель для миристиновой кислоты составляет 8,0-13,0 %, для пальмитиновой — 22,0-33,0 %, стеариновой — 9,0-13,0 %, олеиновой — 22,0-32,0 % (4-7).
Жирные кислоты действуют на организм человека неодинаково: миристиновая оказывает негативное влияние на сердечно-сосудистую систему, вызывая заболевания, а стеариновая не имеет такого эффекта. Отмечено, что присутствие жирных кислот в организме животного в большей степени определяется их генетически детерминированным синтезом, нежели поступлением с кормом либо мобилизацией из жировых тканей организма. Для образования жирных кислот С6:0-С16:0, согласно данным литературы, характерна высокая наследуемость (h2 = 0,41-0,43), и это повышает эффективность селекции. Образование важных для здоровья человека ЖК (С18:2 cis-9,12) характеризуется относительно низкой наследуемостью (h2 = 0,17-0,33), но так как их продукция отрицательно коррелирует с синтезом коротких и средних по длине цепи ЖК, то селекция по этому признаку также может быть успешной (2).
Известно, что соединения с молекулярной массой по углероду С18:0-С18:1 влияют на фертильность на ранней стадии лактации, а количество С18:1 cis-9 опосредованно указывает на энергетический статус коровы и может служить для раннего прогнозирования кетозов. С16:0 и С17:1 cis-9 представляют собой удобный инструмент для оценки продукции метана и конверсии корма у коров (чем ниже выделение метана, тем лучше переваривается корм) (8, 9).
Для повышения эффективности отбора животных и поиска информативных ДНК маркеров продуктивности большой интерес представляет анализ функции каждого из компонентов молока в связи с тем или иным биологическим признаком и изучение генетической и геномной изменчивости признаков. Так, ведутся исследования по идентификации каузальных нуклеотидов (точечных мутаций) для количественных признаков (QTL), которые наравне со множеством известных некодирующих полиморфных замен (SNP) могут повысить точность детекции соответствующих мутаций и прогноза племенной ценности животного. Ведутся работы по оптимизации числа SNP с высокой долей изменчивости каузальных вариантов, достаточного для построения геномной матрицы родства при учете информации по большому числу генотипов и повышения точности оценок (10). Так, каузальные SNP, достоверно ассоциированные с хозяйственно полезными признаками у молочного скота, были детектированы на 5-й, 6-й, 9-й, 14-й, 15-й и 20-й хромосомах. Наибольшую генетическую дисперсию по показателям молочной продуктивности имели полиморфизмы, расположенные близко либо внутри генов DGAT1 (BTA14), GHR (BTA20), ABCG2 (BTA6) (11-15).
Увеличение плотности SNP (уменьшение расстояния между ними) повысит вероятность выявления QTL и (до некоторой степени) точность картирования. Полногеномные ассоциации были использованы для анализа состава жира в молоке коров голштинской и джерсейской пород датского происхождения (16). Помимо стандартной процедуры генотипирования на чипах высокой плотности (777К), в этом исследовании использовалась база метаболических путей KEGG PATHWAY Database (биоинформатический ресурс для анализа геномов, https://www.genome.jp/kegg/pathway.html). Отмечено, что ген-кандидат DGAT1, который очень часто фигурирует в исследованиях молочной продуктивности крупного рогатого скота, не определялся как играющий заметную роль в жировом составе молока, что еще раз указывает на сложность наследования этого признака. Однако для гена SCD, участвующего в катализируемом превращении кислот С10:0 в С18:0, и ACSS3, вовлеченного в активацию и внутриклеточный перенос жирных кислот, были найдены значимые ассоциации с жирнокислотным составом молока.
В работе F. Kawaguchi (17) у японского черного крупного рогатого скота на основе аллельных различий между группами с высоким и низким содержанием олеиновой кислоты С18:1 с помощью полногеномного исследования ассоциации (GWAS) обнаружены 1993 полиморфизма в 23 генах. Среди этих 23 генов на основе анализа их функции в метаболизме жирных кислот определены три гена-кандидата, влияющие на изменчивость со-держания олеиновой ЖК — CYB5R4, MED23 и VNN1.
На итальянской популяции симменталов и голштинов по результатам GWAS для жирных кислот молока выявили значимые сигналы на хромосомах BTA19 и BTA26. Дальнейший анализ идентифицировал не только некоторые хорошо известные гены (FASN, SCD и DGAT1) локусов количественных признаков для компонентов молочных ЖК, но и другие значимые гены-кандидаты, которые были связаны с функциональными ролями в путях липидного метаболизма. Выявленные мутации, которые связаны с профилем жирных кислот, находятся в генах ECI2, PCYT2, DCXR, G6PC3, PYCR1, ALG12, CYP17A1, ACO2, PI4K2A, GOT1, GPT, NT5C2, PDE6G, POLR3H и COX15 (18).
Обнаружение локусов количественных признаков и входящих в них генов, связанных с составом молочного жира, может дать ценную информацию о сложных метаболических сетях, лежащих в основе изменений в синтезе жирных кислот, и указать на возможные «точки воздействия» для улучшения состава молочного жира посредством селекции. C. Li с соавт. (19) провели GWAS анализ по 22 жирным кислотам молока у 784 коров голштинской породы китайской популяции. Всего обнаружили 83 значимых SNP и 314 предполагаемых суггестивных SNP для 18 признаков, связанных с метаболизмом жирных кислот молока. Участки хромосом, влияющие на свойства ЖК молока, в основном локализовались на BTA1, BTA2, BTA5, BTA6, BTA7, BTA9, BTA13, BTA14, BTA18, BTA19, BTA20, BTA21, BTA23, BTA26 и BTA27. Из них 146 SNP оказались связаны более чем с одним признаком в метаболизме жирных кислот молока; большинство были статистически значимо связаны с несколькими SNP, особенно содержание ЖК C18:0 (105 SNP), C18 (93 SNP) и С14 (84 SNP). Некоторые SNP находятся рядом или внутри генов DGAT1, SCD1 и FASN, которые, как известно, влияют на состав молока у молочного скота. Кроме того, выявлены 20 новых высокодостоверных полиморфизмов-кандидатов для С10:0, C12:0, C14:0, C14:1, индекса С14, C18:0, C18:1n9c, индекса C18, включая мутации в генах HTR1B, CPM, PRKG1, MINPP1, LIPJ, LIPK, EHHADH, MOGAT1, ECHS1, STAT1, SORBS1, NFKB2, AGPAT3, CHUK, OSBPL8, PRLR, IGF1R, ACSL3, GHR и ОХСТ1 (19).
Особое место в анализе химического состава молока занимает концентрация жирных кислот, так как характеристики качества этого продукта (выход сыра, масла и сливок) в значительной степени зависят от липидного обмена в организме животного. В исследованиях P. Gottardo (20) на 2977 голштинских, бурых швицких и симментальских коровах было установлено, что лучшим соотношением доли насыщенных и ненасыщенных жирных кислот от общего содержания жира в молоке характеризовались животные голштинской породы. Симментальские коровы по этому показателю занимали промежуточное положение, далее следовали животные бурой швицкой породы (20).
Исследования жирнокислотного состава молока коров с использованием метода инфракрасной спектрометрии широко применяются во многих странах на местных популяциях крупного рогатого скота разных пород. На содержание жирных кислот молока оказывает влияние множество факторов, поэтому для более точного ответа об эффективности селекции по таким признакам требуется изучение показателей изменчивости и наследуемости. Для нашей страны ЖК являются сравнительно новыми признаками для отбора животных и повышения качества молока. Однако в ряде западноевропейских стран жирнокислотный состав молока применяется в селек-ции не только как критерий оценки фракций насыщенных и ненасыщенных ЖК, но также для контроля за физиологическим состоянием животных и полноценностью их кормления. В этой связи накопление информации, полученной на уровне популяции в стадах, а также в экспериментальных исследованиях, позволит уточнить перспективы отбора животных по ЖК. Валидация генетических полиморфизмов, связанных с изменчивостью жирнокислотного состава молока коров наряду с другими компонентами, даст новые знания по геномной архитектуре показателей молочной продуктивности.
В настоящей работе на российской популяции коров голштинизированной черно-пестрой и голштинской пород впервые определены параметры генетической вариабельности фракционного состава жирных кислот в молоке для использования этих данных в селекционном процессе и генетическом улучшении животных. С помощью инфракрасной спектрометрии молока и высокопроизводительного геномного сканирования в условиях экспериментальных исследований создана база данных прямых фенотипов и генотипов коров, которая позволила определить локусы количественных признаков и функциональные мутации, регулирующие синтез липидов молока в организме животных.
Цель исследований состояла в поиске полногеномных ассоциаций и полиморфизмов в генах, детерминирующих жирнокислотный состав молока коров. Для его определения применяли инфракрасную спектрометрию как один из наиболее быстрых и точных экспресс-методов физико-химического анализа состава молока.
Методика. Для формирования баз данных использовали результаты межпопуляционных наблюдений и исследований в экспериментальной популяции. На первом этапе были проведены популяционного-генетические исследования содержания жирных кислот (на основе ИК-спектров) в периоды контрольных доений (общее число образцов молока 36982) в течение 9 мес 2017-2018 годов. В исследуемую базу вошли 14 племенных стад голштинизированного черно-пестрого и голштинского скота из Московской области (размер выборки животных n = 11529). Созданная база данных компонентного состава молока стала основой для изучения популяционно-генетических параметров и определения изменчивости содержания в молоке следующих жирных кислот и компонентов: миристиновой (C14:0), пальмитиновой (C16:0), стеариновой (C18:0), олеиновой (C18:1) кислот, насыщенных ЖК (НЖК), мононенасыщенных ЖК (МНЖК), полиненасыщенных ЖК (ПНЖК), короткоцепочечных ЖК (КЦЖК), среднецепочечных ЖК (СЦЖК), длинноцепочечных ЖК (ДЦЖК) и транс-изомеров ЖК (ТЖК), массовой доли жира (МДЖ), белка (МДБ), казеина (МДК), лактозы (МДЛ), сухого вещества (СВ), СОМО (сухого обезжиренного молочного остатка), следов ацетона и бета-гидроксибутирата (БГБ), концентрации мочевины, точки замерзания и кислотности.
Для расчета наследуемости показателей жирнокислотного состава молока и генетических корреляций использовали следующее уравнение смешанной модели:
y = μ + HFMTDj + Agek + Lactl + sirem + ejklm,
где y — исследуемый показатель молока коров-дочерей; μ — средняя популяционная константа по выборке из 14 стад; HFMTDj — эффект хозяйства, месяца и даты контрольной дойки; Agek — возраст 1-го отела; Lactl — эффект номера последней завершенной лактации; sirem— эффект быка-отца; ejklm — случайная ошибка (нераспределенная варианса). Расчет по 14 стадам проводили на основе метода REML (residual maximum likelihood), суть которого состоит в максимизации правдоподобия получаемых значения варианс через множественную процедуру итераций с использованием семейства программ BLUPF90 (21, 22).
Первичные данные по составу жирных кислот молока в экспериментальном стаде голштинизированных черно-пестрых коров получили в ПЗ «Ладожский» (филиал ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, Краснодарский край, 2020-2021 годы). Сформировали группу из 144 коров, у которых было предварительно получено полное фенотипическое описание спектра ЖК и компонентов молока, а также выполнено генотипирование с помощью биочипа Bovine GGP 150K («Neogen», США). Выделение ДНК из ушных выщипов животных, SNP генотипирование и анализ проб молока проводили на базе объекта научной инфраструктуры «Биотехнология животных» (ОНИС БиоТехЖ) при ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста по стандартным протоколам.
Компонентный состав и жирные кислоты молока коров для популяционного анализа в стадах и в экспериментальной группе животных определяли на автоматическом анализаторе MilkoScan 7 DC («FOSS», Дания) по показателям, аналогичным для популяционных исследований (см. выше). В основе действия прибора лежит инфракрасная спектроскопия (применялся протокол, рекомендованный производителем). Продуктивность животного учитывали индивидуально при числе контрольных доений от 5 до 12 (в среднем 8,9 на одно животное). Пробы молока отбирали в стаканчики объемом 50 мл (использовался консервант «Microtabs», США) в момент доения животных утром, в обед и вечером.
Контроль качества генотипирования (110884 SNP), анализ полногеномных ассоциаций (GWAS, genome-wide association study) и многомерного шкалирования (MDS, multidimensional scaling) выполняли с помощью программы Plink 1.9 (23). В результате сформирована выборка генотипов из 143 геномных SNP-профиля коров экспериментальной группы с уровнем генотипирования от 99,3 до 99,7 %.
Поиск генов по выявленным значимым полиморфизмам по результатам GWAS проводили в браузере Ensembl (https://www.ensembl.org/index.html) по сборке генома крупного рогатого скота версии UMD 3.1. Аннотацию генов для определения локусов количественных признаков на хромосомах животных осуществляли по международной базе данных Animal QTLdb (24).
С использованием пакета «Анализ данных» в среде MS Excel 2013 рассчитывали средние значения (M) и их стандартные ошибки (±SEM), а также среднеквадратическое отклонение (SD) для изученных показателей. Степень изменчивости признаков оценивали по коэффициенту фенотипической вариации (Cvp). Для расчета наследуемости (h2) использовали отношение генетической вариансы к сумме генетической и остаточной варианс.
Результаты. У коров из всех компонентов молока жирные кислоты по количественному составу наиболее подвержены высокой изменчивости под влиянием как условий среды (например, кормление, технология содержания), так и генетических факторов (порода, предки, генотип). Соотношение жирных кислот в зависимости от числа атомов углерода, длины цепи и степени ее насыщения различается как между особями, так и на популяционном уровне. Понимание механизмов синтеза молочных ЖК важно для определения детерминирующих их локусов количественных признаков (QTL). Мы изучили генетическую изменчивость содержания жирных кислот в молоке коров из популяций голштинизированной черно-пестрой и голштинской пород на примере нескольких стад Подмосковья, чтобы уточнить перспективность этих признаков для селекции (табл. 1).
Наследуемость количественного состава жирных кислот молока варьировала от низких значений для полиненасыщенных ЖК (h2 = 0,018) до умеренных для средне- (h2 = 0,125), длинноцепочечных (h2 = 0,155) и миристиновой кислоты (h2 = 0,155), мононенасыщенных ЖК (h2 = 0,176) и олеиновой кислоты (h2 = 0,196).
Генетические корреляции между показателями удоя, массовой долей жира (МДЖ) и жирнокислотным составом молока представлены в таблице 2 (если rg> 0,050, полученные коэффициенты достоверны при p < 0,001). Установлено, что генетическая связь суточного удоя и МДЖ практически отсутствовала (rg = -0,032), при этом наблюдалась более тесная отрицательная корреляция с содержанием транс-изомеров ЖК (rg = -0,129), миристиновой кислоты (rg = -0,110) и СЦЖК (rg = -0,106). Взаимосвязь МДЖ и содержания различных ЖК колебалась в пределах значений rg от 0,393 для олеиновой до 0,955 для НЖК, для ТЖК (rg = -0,286) и более желательным ПНЖК (rg = -0,465) значения были отрицательными. Содержание насыщенных ЖК миристиновой и пальмитиновой отрицательно коррелировало с количеством ненасыщенной олеиновой (rg от -0,160 до -0,427), МНЖК (rg от -0,072 до -0,337), ПНЖК (rg от -0,554 до -0,584) и с содержанием кислот из более сложной группы с высокой молекулярной массой — ДЦЖК (rg от -0,030 до -0,325). Связь между количеством ЖК C14:0, C16:0, с одной стороны, и ТЖК — с другой показывала, что при увеличении содержания насыщенных ЖК в молоке снижались следовые количества изомеров, блокирующих синтез молочного жира (rgсоответственно от -0,469 до -0,637).
Количество ПНЖК отрицательно коррелировало с количеством КЦЖК (rg = -0,455) и СЦЖК (rg= -0,596), но взаимосвязь между продукцией ПНЖК и ТЖК была наиболее высокой и положительной (rg = 0,469), что в сочетании с коэффициентом наследуемости свидетельствует о сложном селекционном процессе повышения этих показателей в молоке коров. Однако необходимо отметить, что проблема улучшения соотношения насыщенных и ненасыщенных ЖК в молоке в пользу последних остается нерешенной. Увеличение доли олеиновой кислоты, МНЖК, ПНЖК, ДЦЖК в молоке при отборе животных одновременно приводит к изменению жирнокислотного состава по транс-изомерам, ЖК с короткой и средней длиной углеродной цепи.
Суммируя полученные данные (см. табл. 1, табл. 2), можно заключить, что выявленные корреляции и показатели генетической изменчивости жирных кислот в молоке коров перспективны для дальнейшего GWAS анализа с целью корректировки программ селекции крупного рогатого скота.
Результаты анализа полученных в контрольных дойках образцов молока коров из экспериментальной группы приведены в таблице 3. Было установлено, что 50,5 % суточного удоя приходилось на утро, остальное количество распределялось между дневной и вечерней дойками примерно поровну — соответственно 23,5 и 26,0 %. Прослеживалась явная обратная линейная зависимость между величиной суточного удоя и компонентным составом молока. При меньшем объеме молока за обеденное доение в 6,1 кг повышалось процентное содержание жировой фракции (до 4,15 %, включая жирные кислоты) и сухого вещества (13,33 %).
Для белка и казеина молока, лактозы, СОМО, БГБ и мочевины не установлено значительных количественных различий при отборе образцов в разное время. Значения молярной массы ацетона, выявляемого в следовых количествах, были выше в утренних и вечерних пробах молока (соответственно 0,047 и 0,040 ммоль/л), тогда как точка замерзания — ниже (соответственно -536,5×10-3 и -537,2×10-3 °С). Коэффициент фенотипической вариации (Cvp) вне зависимости от времени (утро, обед, вечер) отбора проб молока (20,0-24,2 %) был выше для массовой доли жира, чем для других селекционно значимых признаков качества молока. Исходя из фенотипической изменчивости, можно полагать, что потенциальная эффективность отбора по жирным кислотам будет выше для пальмитиновой (Cvp = 22,0-25,0 %), стеариновой (Cvp= 24,6-32,1 %) кислот, длинноцепочечных ЖК (Cvp = 20,2-27,8 %), короткоцепочечных ЖК (Cvp = 23,0-27,5 %), а также суммы насыщенных ЖК (Cvp = 21,5-25,8 %).
Повторяемость (repeatability, r) между смежными контрольными доениями (утро и вечер) по изученным показателям состава молока была достаточно высокой, исключение касалось следов метаболитов — ацетона, БГБ и мочевины (r = 0,565-0,630) и точки замерзания молока (r = 0,505). Сравнительно умеренные значения были получены для олеиновой кислоты (r = 0,625) и полиненасыщенных ЖК (r = 0,590). В целом можно заключить, что контроль компонентного состава молока коров вполне целесообразно осуществлять либо по средней пробе, либо по отдельности утром и вечером, тогда как учет объема надоенного молока от коровы должен быть равен кратности доений в сутки. Полагаем, что при анализе полногеномных ассоциаций не будет значительного смещения в выявленных QTL при использовании данных, полученных при 2-кратном контроле продуктивности в сутки, то есть такого контроля достаточно.
Мы также определили генетическую структуру экспериментальной группы животных с помощью метода многомерного шкалирования (MDS) (рис. 1). Так как стадо представлено голштинизированной черно-пестрой породой молочного крупного рогатого скота, наблюдался умеренный размах вариабельности по компонентам изменчивости (PC1 = 7,82 %, PC2 = 4,65 %). В настоящее время в экспериментальном стаде проводится консолидация поголовья посредством индивидуального подбора родительских пар (коров и голштинских быков-производителей), чтобы получить как можно более однородных в генетическом отношении особей для изучения наследования количественных признаков, включая компонентный состав молока. По нашему мнению, это позволит с более высокой точностью вести оценку генотипа коров на основе фенотипирования хозяйственно полезных качеств каждого животного. Результаты MDS анализа выборки использованы нами как коварианты по осям PC1/PC2 для корректировки влияния генетической изменчивости на популяционную структуру экспериментального стада и снижения вероятности получения ложноположительных значений GWAS ассоциаций с прямыми фенотипическими данными по ряду компонентов молока и содержанию в нем жирных кислот.
Ранее в одном из исследований мы детектировали 32 (p < 0.001-0.00001) каузальные SNP мутации, ассоциированные с оценкой племенной ценности быков по содержанию ЖК в молоке коров-дочерей (наиболее значимые находились на хромосомах BTA1, BTA5, BTA6, BTA10, BTA11, BTA14, BTA19, BTA22 и BTA26) (25). Гены CHST11, ACO2, PPARGC1A, NRXN1, LPIN1, ASIC2, PCDH15, PRKG1 оказались напрямую связаны с синтезом ЖК С14, С16, С18, конъюгированной линолевой кислоты, с индексом насыщенных и ненасыщенных ЖК. Также были обнаружены гены, входящие в QTL, которые ассоциированы с показателями фертильности животных, линейными промерами вымени и конечностей (NCAM2, FGD4, KCNIP4, SFXN1, NBAS, PGR, MON1B, GPLD1, PRKG1). Анализ международной базы данных NCBI (https://www.ncbi.nlm.nih.gov/) по выявленным полиморфизмам показал, что выявленные гены часто проявляют плейотропное действие. Это еще раз подтверждает сложный характер наследуемости количественного состава ЖК и одновременно указывает на возможность использовать содержание ЖК в молоке для управления показателями здоровья и фертильности молочных коров (25).
Для дальнейшего поиска полногеномных ассоциаций с количественным составом ЖК молока мы соотнесли данные GWAS анализа с результатами прямого фенотипирования коров экспериментального стада по этому признаку и выявленными SNP, которые ассоциированы с собственной продуктивности дочерей быков-отцов, оцененных на популяции из Подмосковья (14 стад). Для показателей племенной ценности этих быков-отцов по ЖК ранее были получены первые результаты поиска ассоциаций.
Установлено, что для всех изученных признаков молочной продуктивности коров был характерен полигенный характер наследования и множественное действие генов, вовлеченных в контроль показателей количественного состава жирных кислот молока с разной селекционной значимостью. Так, для суточного удоя молока мы обнаружили локусы количественных признаков (QTL) на хромосомах BTA1, BTA4, BTA7, BTA9, BTA14, BTA15, BTA18 и BTA25 (рис. 2). Изменчивость массовой доли жира (МДЖ) при GWAS анализе послужила индикатором достоверности фенотипирования остальных изученных признаков: спектры основных жирных кислот и МДЖ определяли в одних и тех же образцах, поэтому выявленные ассоциации можно считать значимыми. На хромосоме BTA14 мы обнаружили кластеры, связанные с процентом молочного жира, в которых находится ген-кандидат жирномолочности DGAT1 (SNP мутация ARS-BFGL-NGS-4939) и ряд других сцепленных с ним генов. Также были детектированы QTL по массовой доле жира на хромосомах BTA5, BTA10, BTA11 и BTA19.
Для миристиновой и пальмитиновой ЖК мы выявили общие кла-стеры на BTA5, BTA10, BTA14, BTA18 и BTA27, что в значительной степени согласовывалось с профилем ассоциаций для МДЖ молока. Стеариновая и олеиновая ЖК как длинноцепочечные ЖК показали схожую локализацию на хромосомах BTA9, BTA10, BTA11, BTA14, BTA17, BTA18, BTA19, BTA20 и BTA29 (см. рис. 2). Для коротко- и среднецепочечных ЖК детектировались ассоциации на хромосомах BTA1, BTA5, BTA10, BTA11, BTA14, BTA18, BTA19 и BTA24. В этом отношении КЦЖК и СЦЖК оказались в большей степени схожими с миристиновой и пальмитиновой кислотами, по содержанию которых в молоке отмечались тесные генетические корреляции: для КЦЖК — соответственно rg = 0,551 и rg = 0,608, для СЦЖК — rg = 0,920 и rg = 0,980. Для длинноцепочечных ЖК были детектированы QTL на BTA6, BTA7, BTA9, BTA10, BTA11, BTA17, BTA18 и BTA29 (см. рис. 2), что в целом согласовывалось с данными для стеариновой и олеиновой кислот, имеющими схожий профиль ассоциаций, и с выявленной генетической взаимосвязью между этими признаками (rg = 0,831 для стеариновой кислоты, rg = 0,979 — для олеиновой).
Группа насыщенных и ненасыщенных ЖК по выявленным ассоциациям показала разный характер локализации QTL, что, по нашему мнению, преимущественно связано с особенностями их синтеза и метаболическими путями (рис. 3). Моно- и полиненасыщенные ЖК (как наиболее значимые с точки зрения питательной ценности молока) показывали общие QTL на BTA1, BTA2, BTA9, BTA11, BTA18 и BTA19 (см. рис. 3). Наличие разных локусов, контролирующих образование и секрецию ЖК молока, и их расположение на разных хромосомах могут быть также связаны с давлением искусственного отбора.
Для транс-изомеров ЖК не были детектированы значимые QTL, за исключением выявленных на BTA1, BTA6, BTA18, BTA22 и BTA27, что, вероятно, связано с малой дисперсией по этому показателю (данные на рисунках не приведены).
Мы аннотировали выявленные полиморфизмы в генах, связанных с величиной суточного удоя, массовой долей жира и жирнокислотным составом молока у коров из экспериментальной группы (рис. 4). Сравнение проводили по международной базе Animal QTLdb (https://animalgenome.org/cgi-bin/QTLdb/BT/index).
При картировании локусов количественных признаков для суточного удоя мы выявили наличие трех высокодостоверных ассоциаций с полиморфизмами в генах CDH13 и AKTIP (BTA18) и ABCC1 (BTA25) (см. рис. 4). Они также связаны с содержанием холестерина в молоке, фертильностью животных, долголетним использованием и числом соматических клеток в молоке (24). Для МДЖ, коротко-, среднецепочечных, насыщенных ЖК, C14:0, C16:0, C18:0 и C18:1 были установлены гены CACNA1C, GCH1, ATG14, KCNH5, PRKCE, CTNNA2, CYHR1, VPS28, DGAT1, ZC3H3, RHPN1, TSNARE1, образующие QTL на хромосомах BTA10, BTA11 и BTA14. Следует отметить, что все перечисленные гены обладали плейотропным действием в отношении ряда жирных кислот. При аннотировании были обнаружены гены, сопряженные с энергетическим метаболизмом, определяющим устойчивость к кетозу (26), содержанием конъюгированной линолевой кислоты в молоке, процентом и выходом молочного жира и белка, содержанием холестерина в молоке, содержанием пальмитиновой и пальмитолеиновой жирных кислот, удоем за лактацию, воспроизводительными качествам животных. С помощью GWAS анализа мы выявили ген диацилглицерол-О-ацилтрансферазы 1, который может служить маркером полиморфизма при изучении показателей жирномолочности и позволяет опосредованно оценить точность полу-ченных результатов (14, 27). Мы выявили 70 различных QTL, преимущественно ассоциированных с профилем жирных кислот в молоке коров, содержанием казеинов, энергетическим статусом животных, содержанием кальция, калия и фосфора в молоке (24, 28).
Коротко- и среднецепочечные ЖК, миристиновая и пальмитиновая кислоты, насыщенные ЖК показали связь с полиморфизмами в генах MED12L, EPHB1, GRIN2B, PRMT8, ERC1, PELI2, ARHGAP39, MROH1, MAF1, GSDMD, LY6D (см. рис. 4), которые были ассоциированы с количеством плодотворных осеменений, легкостью отела, коэффициентом стельности дочерей быков, процентным содержанием жира и белка в молоке, пальмитиновой ЖК, прикреплением и глубиной вымени коров, маститом и числом соматических клеток в молоке (24).
Для длинноцепочечных, мононенасыщенных ЖК, стеариновой и олеиновой кислот в результате аннотации выявлены следующие селекционно значимые гены: RPS6KA2, CPQ, CPE, FTO, FAT3, LUZP2 (см. рис. 4). Их полиморфизмы также сопряжены с изменчивостью предрасположенности коров к маститу, линейными промерами экстерьера животных (конечности и вымя), фертильностью, выходом молочного жира и числом соматических клеток в молоке (29). Транс-изомеры ЖК, несмотря на низкую вариабельность по сравнению с другими фракциями жирных кислот молока, в нашем исследовании показали связь с полиморфизмами в генах ARFGEF3 и TNKS, о которых известно, что они ассоциировались с удоем молока коров (30).
Таким образом, на основе изучения генетической и фенотипической вариабельности состава молока на популяционном уровне в 14 стадах молочного скота (Московская область) и затем в экспериментальном стаде коров голштинизированной черно-пестрой породы (Краснодарский край) установлены селекционные константы и ряд достоверных ассоциаций между выявленными полиморфизмами генов и образованием жирных кислот молока. С использованием популяционно-генетического анализа по соотношению межгрупповой и общегрупповой варианс показана наибольшая наследуемость содержания олеиновой (h2 = 0,196) кислоты, мононенасыщенных жирных кислот (ЖК) (h2 = 0,176), длинно- и среднецепочечных ЖК (h2 = 0,125-0,55), стеариновой (h2 = 0,125) кислоты. Эти признаки могут быть рекомендованы при оценке быков-производителей по качеству потомства. Исследования, проведенные на группе коров, генотипированных и фенотипированных по расширенному компонентному составу молока, позволили получить новые данные о локализации QTL жирнокислотного состава в геномах животных российского происхождения. В результате аннотации в генах CACNA1C, ARFGEF3, RPS6KA2, GCH1, ATG14, PELI2, KCNH5, PRKCE, CTNNA2, ARHGAP39, CYHR1, VPS28, DGAT1, MROH1, MAF1, GSDMD, ZC3H3, RHPN1, LY6D, TSNARE1, CPQ, CPE, FTO, TNKS, FAT3 и LUZP2 установлены полиморфизмы, достоверно ассоциированные с изменчивостью содержания ЖК в молоке. Насыщенные жирные кислоты в сравнении с ненасыщенными показали большую вариабельность результатов GWAS, что, вероятно, связано с более сильным селекционным давлением. Дальнейшее изучение генетических механизмов наследования жирнокислотного состава молока позволит разработать основу стратегии селекции по этому признаку.
1. Фенотипические и генетические параметры состава жирных кислот в молоке коров (Bos taurus) из 14 племенных стад голштинизированного черно-пестрого и голштинского скота по данным контрольных доений (Московская обл., 2017-2018 годы) |
||||
Жирная кислота, группа жирных кислот |
г/100 г молока |
Cvp, % |
h2 |
|
M±SEM |
SD |
|||
Миристиновая |
0,680±0,001 |
0,148 |
21,8 |
0,155 |
Пальмитиновая |
1,845±0,003 |
0,633 |
34,3 |
0,071 |
Стеариновая |
0,585±0,001 |
0,195 |
33,4 |
0,125 |
Олеиновая |
0,875±0,001 |
0,281 |
32,2 |
0,196 |
Насыщенные |
3,600±0,004 |
0,861 |
23,9 |
0,083 |
Мононенасыщенные |
1,002±0,001 |
0,213 |
23,0 |
0,176 |
Полиненасыщенные |
0,005±0,000 |
0,011 |
238,8 |
0,018 |
Короткоцепочечные |
0,538±0,001 |
0,113 |
21,1 |
0,114 |
Среднецепочечные |
2,206±0,003 |
0,652 |
29,6 |
0,125 |
Длинноцепочечные |
1,486±0,002 |
0,463 |
31,2 |
0,155 |
Примечание. Общее число образцов n = 36982. Cvp— коэффициент фенотипической вариации, h2 – коэффициент наследуемости. |
||||
2. Генетические корреляции (rg) между суточным удоем, массовой долей жира и количественным составом жирных кислот |
||||||||||||
Признак |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
МДЖ |
-0,032 |
|||||||||||
C14:0 |
-0,110 |
0,634 |
||||||||||
C16:0 |
-0,101 |
0,801 |
0,874 |
|||||||||
C18:0 |
-0,023 |
0,615 |
-0,122 |
0,300 |
||||||||
C18:1 |
0,053 |
0,393 |
-0,427 |
-0,160 |
0,764 |
|||||||
НЖК |
-0,031 |
0,955 |
0,781 |
0,905 |
0,485 |
0,150 |
||||||
МНЖК |
-0,008 |
0,463 |
-0,337 |
-0,072 |
0,767 |
0,983 |
0,211 |
|||||
ПНЖК |
0,075 |
-0,465 |
-0,554 |
-0,584 |
-0,037 |
0,113 |
-0,569 |
0,097 |
||||
КЦЖК |
0,045 |
0,815 |
0,551 |
0,608 |
0,482 |
0,264 |
0,852 |
0,255 |
-0,455 |
|||
СЦЖК |
-0,106 |
0,684 |
0,980 |
0,920 |
-0,033 |
-0,384 |
0,818 |
-0,293 |
-0,596 |
0,554 |
||
ДЦЖК |
0,053 |
0,499 |
-0,325 |
-0,030 |
0,831 |
0,979 |
0,276 |
0,970 |
0,038 |
0,350 |
-0,276 |
|
ТЖК |
-0,129 |
-0,286 |
-0,637 |
-0,469 |
0,282 |
0,384 |
-0,383 |
0,355 |
0,469 |
-0,300 |
-0,643 |
0,396 |
Примечание. Общее число образцов молока 36982; 1 — суточный удой, 2 — массовая доля жира (МДЖ), 3 — C14:0, 4 — C16:0, 5 — C18:0, 6 — C18:1, 7 — насыщенные жирные кислоты (НЖК), 8 — мононенасыщенные жирные кислоты (МНЖК), 9 — полиненасыщенные жирные кислоты (ПНЖК), 10 — короткоцепочечные жирные кислоты (КЦЖК), 11 — среднецепочечные жирные кислоты (СЦЖК), 12 — длинноцепочечные жирные кислоты (ДЦЖК). Eсли rg > 0,050, корреляции достоверны при p < 0,001. |
||||||||||||
3. Результаты расширенного анализа компонентного состава молока коров в зависимости от времени отбора проб (n = 144, экспериментальное стадо голштинизированных черно-пестрых коров, ПЗ «Ладожский», Краснодарский край, 2020-2021 годы) |
|||||||||||||||
Показатель |
Контрольное доение |
Повторяемость |
|||||||||||||
утро |
обед |
вечер |
|||||||||||||
М |
±SEM |
SD |
Cvp, % |
М |
±SEM |
SD |
Cvp, % |
М |
±SEM |
SD |
Cvp, % |
у/о |
о/в |
у/в |
|
Удой, кг |
13,0 |
0,1 |
3,6 |
27,3 |
6,1 |
0,1 |
1,8 |
30,1 |
6,70 |
0,10 |
1,90 |
27,8 |
0,750 |
0,724 |
0,762 |
МДЖ, % |
3,22 |
0,03 |
0,78 |
24,2 |
4,15 |
0,03 |
0,80 |
19,3 |
3,81 |
0,03 |
0,76 |
20,0 |
0,596 |
0,740 |
0,652 |
МДБ (истинный), % |
3,24 |
0,02 |
0,43 |
13,3 |
3,25 |
0,02 |
0,42 |
12,8 |
3,26 |
0,02 |
0,44 |
13,6 |
0,869 |
0,915 |
0,867 |
МДБ (сырой), % |
3,44 |
0,02 |
0,44 |
12,7 |
3,46 |
0,02 |
0,42 |
12,2 |
3,46 |
0,02 |
0,45 |
12,9 |
0,868 |
0,914 |
0,866 |
МДК, % |
2,71 |
0,01 |
0,36 |
13,4 |
2,76 |
0,01 |
0,35 |
12,8 |
2,75 |
0,01 |
0,38 |
13,7 |
0,868 |
0,914 |
0,862 |
МДЛ, % |
4,78 |
0,01 |
0,20 |
4,2 |
4,80 |
0,01 |
0,20 |
4,1 |
4,79 |
0,01 |
0,20 |
4,1 |
0,765 |
0,855 |
0,751 |
СВ, % |
12,33 |
0,04 |
1,11 |
9,0 |
13,33 |
0,04 |
1,09 |
8,1 |
12,97 |
0,04 |
1,12 |
8,7 |
0,734 |
0,835 |
0,764 |
СОМО, % |
9,06 |
0,02 |
0,49 |
5,4 |
9,11 |
0,02 |
0,49 |
5,4 |
9,10 |
0,02 |
0,52 |
5,8 |
0,835 |
0,886 |
0,826 |
Ацетон, ммоль/л |
0,047 |
0,002 |
0,049 |
105,1 |
0,036 |
0,002 |
0,040 |
111,5 |
0,040 |
0,002 |
0,056 |
139,0 |
0,608 |
0,681 |
0,569 |
Ацетон, логарифм |
-1,758 |
0,028 |
0,794 |
45,2 |
-1,930 |
0,030 |
0,817 |
42,3 |
-1,827 |
0,027 |
0,775 |
42,4 |
0,581 |
0,623 |
0,630 |
БГБ, ммоль/л |
0,017 |
0,001 |
0,024 |
137,7 |
0,017 |
0,001 |
0,024 |
136,7 |
0,017 |
0,001 |
0,027 |
156,5 |
0,631 |
0,717 |
0,570 |
БГБ, логарифм |
-2,279 |
0,026 |
0,739 |
32,4 |
-2,282 |
0,027 |
0,738 |
32,3 |
-2,291 |
0,026 |
0,733 |
32,0 |
0,644 |
0,685 |
0,600 |
Мочевина, мгŁ100 мл-1 |
41,3 |
0,2 |
4,9 |
12,0 |
42,1 |
0,2 |
4,8 |
11,5 |
39,9 |
0,2 |
4,8 |
12,1 |
0,617 |
0,746 |
0,565 |
Точка замерзания, |
-536,5 |
0,3 |
8,1 |
1,5 |
-538,5 |
0,4 |
9,5 |
1,8 |
-537,2 |
0,3 |
9,6 |
1,8 |
0,561 |
0,669 |
0,505 |
Кислотность, pH |
6,57 |
0,00 |
0,06 |
0,9 |
6,56 |
0,00 |
0,06 |
0,9 |
6,56 |
0,00 |
0,06 |
0,9 |
0,647 |
0,758 |
0,658 |
C14:0, г/100 г |
0,307 |
0,003 |
0,074 |
24,1 |
0,374 |
0,003 |
0,082 |
21,8 |
0,357 |
0,003 |
0,084 |
23,5 |
0,685 |
0,797 |
0,711 |
C16:0, г/100 г |
0,794 |
0,007 |
0,198 |
25,0 |
0,969 |
0,008 |
0,225 |
23,2 |
0,923 |
0,008 |
0,220 |
23,8 |
0,684 |
0,806 |
0,718 |
C18:0, г/100 г |
0,295 |
0,003 |
0,095 |
32,1 |
0,383 |
0,003 |
0,094 |
24,6 |
0,352 |
0,003 |
0,091 |
25,8 |
0,658 |
0,775 |
0,709 |
C18:1, г/100 г |
1,016 |
0,009 |
0,247 |
24,3 |
1,327 |
0,009 |
0,247 |
18,6 |
1,194 |
0,008 |
0,231 |
19,3 |
0,577 |
0,692 |
0,625 |
ДЦЖК, г/100 г |
1,243 |
0,012 |
0,345 |
27,8 |
1,664 |
0,012 |
0,336 |
20,2 |
1,494 |
0,011 |
0,315 |
21,1 |
0,590 |
0,705 |
0,633 |
СЦЖК, г/100 г |
1,241 |
0,011 |
0,316 |
25,4 |
1,489 |
0,013 |
0,339 |
22,8 |
1,432 |
0,012 |
0,345 |
24,1 |
0,712 |
0,825 |
0,735 |
КЦЖК, г/100 г |
0,437 |
0,004 |
0,120 |
27,5 |
0,576 |
0,005 |
0,133 |
23,0 |
0,524 |
0,004 |
0,125 |
23,8 |
0,609 |
0,758 |
0,637 |
МНЖК, г/100 г |
0,931 |
0,008 |
0,229 |
24,6 |
1,223 |
0,009 |
0,232 |
19,0 |
1,103 |
0,008 |
0,215 |
19,5 |
0,577 |
0,687 |
0,633 |
ПНЖК, г/100 г |
0,124 |
0,001 |
0,025 |
20,3 |
0,150 |
0,001 |
0,026 |
17,3 |
0,135 |
0,001 |
0,024 |
18,0 |
0,545 |
0,656 |
0,590 |
НЖК, г/100 г |
2,139 |
0,019 |
0,551 |
25,8 |
2,730 |
0,022 |
0,587 |
21,5 |
2,542 |
0,020 |
0,571 |
22,5 |
0,636 |
0,775 |
0,676 |
Примечание. Общее число образцов молока2340. МДЖ — массовая доля жира, МДБ — массовая доля белка, МДК — массовая доля казеина, МДЛ — массовая доля лактозы, СВ — сухое вещество, СОМО — сухой обезжиренный молочный остаток, БГБ — бета-гидроксибутират, ДЦЖК — длинноцепочечные жирные кислоты, СЦЖК — среднецепочечные жирные кислоты, КЦЖК — короткоцепочечные жирные кислоты, МНЖК — мононенасыщенные жирные кислоты, ПНЖК — полиненасыщенные жирные кислоты, НЖК — насыщенные жирные кислоты; у/о — повторяемость между показателями контрольных доек утром и днем, о/в — днем и вечером, у/в — утром и вечером. |
|||||||||||||||
4. Результаты аннотации выявленных значимых полиморфизмов (p < 0,0001) на хромосомах крупного рогатого скота (BTA) (n = 144, экспериментальное стадо голштинизированных черно-пестрых коров, ПЗ «Ладожский», Краснодарский край, 2020-2021 годы) |
||||
Ген |
Набор признаков |
BTA |
Позиция, bp |
|
начало |
конец |
|||
MED12L |
СЦЖК, C14:0 |
1 |
117548538 |
117917463 |
EPHB1 |
СЦЖК |
1 |
135191077 |
135518801 |
GRIN2B |
СЦЖК, НЖК, C16:0 |
5 |
96408804 |
96761516 |
PRMT8 |
СЦЖК, НЖК, C16:0 |
5 |
106812249 |
106812249 |
ERC1 |
СЦЖК, C16:0 |
5 |
108308618 |
108549124 |
CACNA1C |
НЖК, C16:0 |
5 |
109152548 |
109417890 |
ARFGEF3 |
ТЖК |
9 |
77035587 |
77158234 |
RPS6KA2 |
ДЦЖК, МНЖК |
9 |
102918982 |
103074109 |
GCH1 |
МДЖ, ДЦЖК, СЦЖК, КЦЖК, НЖК, C14:0, C18:1 |
10 |
67576390 |
67631089 |
ATG14 |
МДЖ, СЦЖК, КЦЖК, НЖК |
10 |
68073407 |
68110299 |
PELI2 |
КЦЖК |
10 |
68778347 |
68974093 |
KCNH5 |
МДЖ, ДЦЖК, СЦЖК, КЦЖК, НЖК, C14:0, C18:1 |
10 |
75235434 |
75637242 |
PRKCE |
МДЖ, C18:0 |
11 |
27935104 |
28472632 |
CTNNA2 |
МДЖ |
11 |
54723190 |
55906462 |
ARHGAP39 |
КЦЖК |
14 |
1563866 |
1600378 |
CYHR1 |
МДЖ, СЦЖК, КЦЖК, НЖК, C14:0, C16:0, C18:0, C18:1 |
14 |
1663923 |
1677519 |
VPS28 |
МДЖ, СЦЖК, КЦЖК, НЖК, C14:0, C16:0, C18:0, C18:1 |
14 |
1693641 |
1698490 |
DGAT1 |
МДЖ, СЦЖК, КЦЖК, НЖК, C14:0, C16:0 |
14 |
1795351 |
1804562 |
MROH1 |
СЦЖК, КЦЖК, НЖК, C14:0, C16:0 |
14 |
1844664 |
1894424 |
MAF1 |
СЦЖК, КЦЖК, НЖК |
14 |
1921784 |
1924818 |
GSDMD |
СЦЖК, C16:0 |
14 |
2341290 |
2346302 |
ZC3H3 |
МДЖ, СЦЖК, КЦЖК, НЖК, C14:0, C16:0 |
14 |
2354390 |
2418557 |
RHPN1 |
C14:0 |
14 |
2462544 |
2471434 |
LY6D |
СЦЖК, C16:0 |
14 |
2801383 |
2803020 |
TSNARE1 |
МДЖ, СЦЖК, КЦЖК, НЖК, C14:0, C16:0 |
14 |
3054763 |
3171546 |
CPQ |
C18:0, C18:1 |
14 |
69287302 |
69893052 |
CPE |
ДЦЖК, МНЖК |
17 |
546398 |
697915 |
CDH13 |
Суточный удой |
18 |
9512739 |
10162782 |
AKTIP |
Суточный удой |
18 |
21926577 |
21937955 |
FTO |
МНЖК, C18:0, C18:1 |
18 |
22118201 |
22541532 |
ABCC1 |
Суточный удой |
25 |
14469282 |
14570639 |
TNKS |
ТЖК, C16:0 |
27 |
24632930 |
24789416 |
FAT3 |
ДЦЖК, МНЖК, C18:0, C18:1 |
29 |
1965869 |
2605125 |
LUZP2 |
ДЦЖК, МНЖК, C18:0, C18:1 |
29 |
20259769 |
20557376 |
Примечание. МДЖ — массовая доля жира, ДЦЖК — длинноцепочечные жирные кислоты, СЦЖК — среднецепочечные жирные кислоты, КЦЖК — короткоцепочечные жирные кислоты, МНЖК — мононенасыщенные жирные кислоты, ПНЖК — полиненасыщенные жирные кислоты, НЖК — насыщенные жирные кислоты, ТЖК — транс-изомеры жирных кислот. |
||||
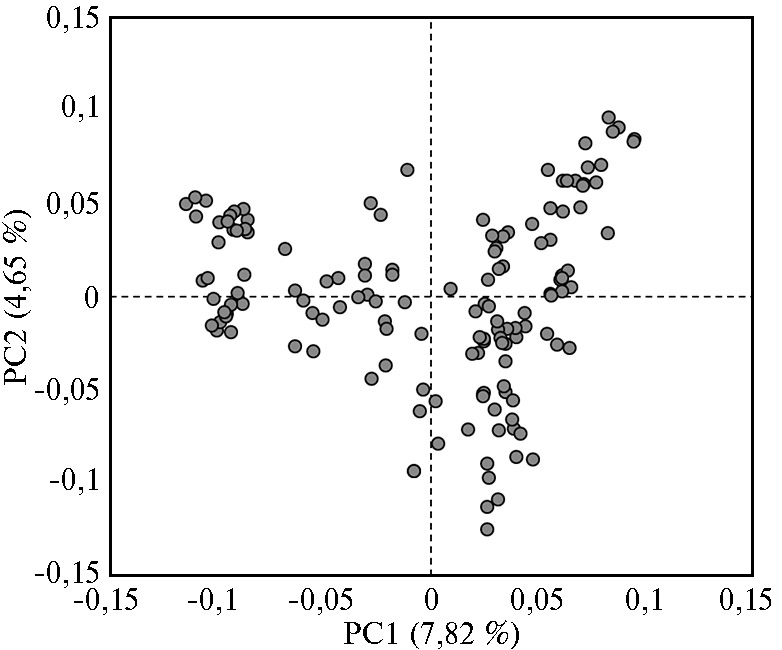
Рис. 1. Результаты анализа генетической структуры группы коров с применением метода многомерного шкалирования (n = 144, экспериментальное стадо голштинизированных черно-пестрых коров, ПЗ «Ладожский», Краснодарский край, 2020-2021 годы).
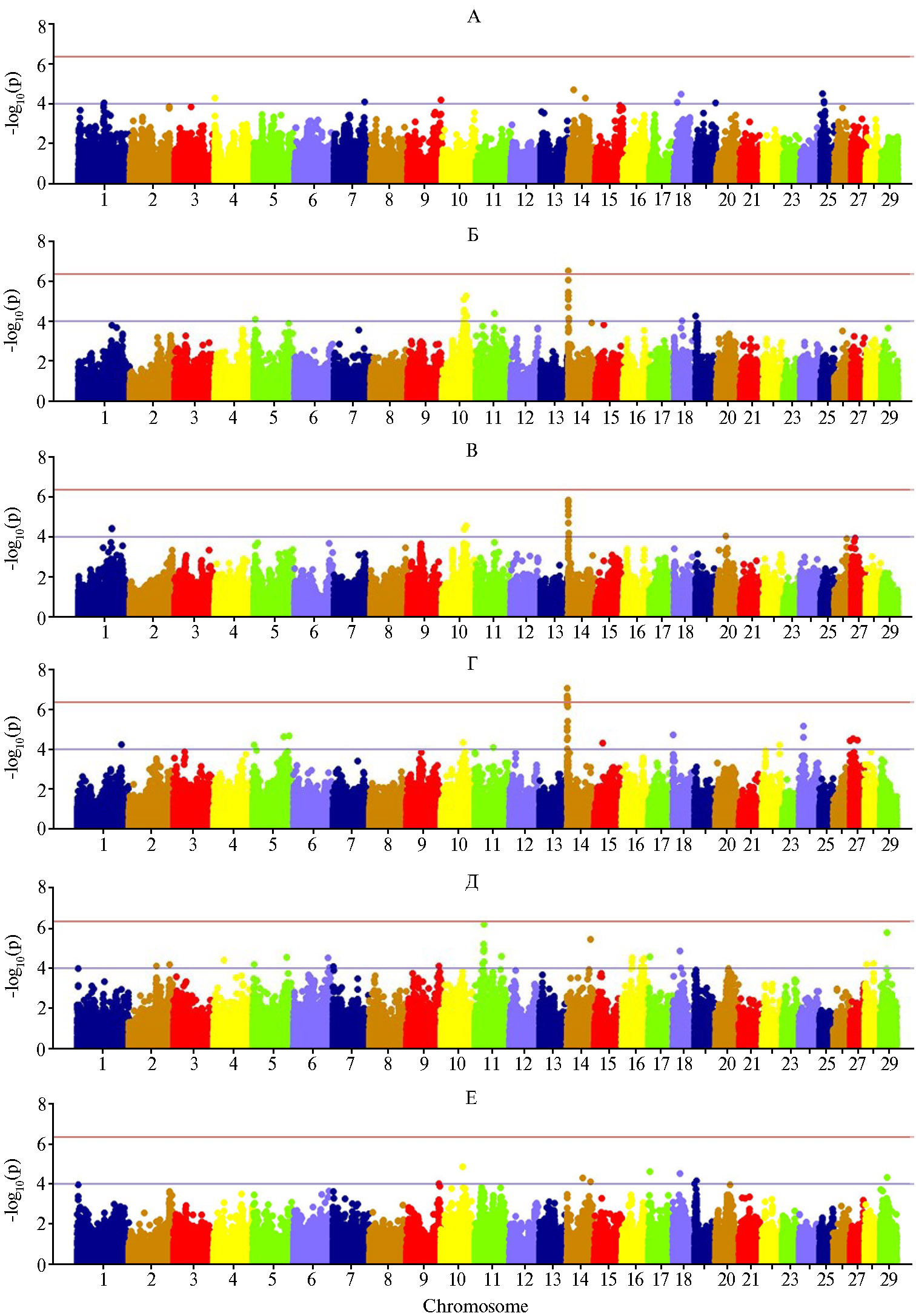
Рис. 2. Результаты GWAS анализа для суточного удоя (А), массовой доли жира (Б) и основных жирных кислот в молоке — миристиновой кислоты (В), пальмитиновой кислоты (Г), стеариновой кислоты (Д) и олеиновой кислоты (Е) на основе прямых фенотипических показателей (n = 144, экспериментальное стадо голштинизированных черно-пестрых коров, ПЗ «Ладожский», Краснодарский край, 2020-2021 годы).
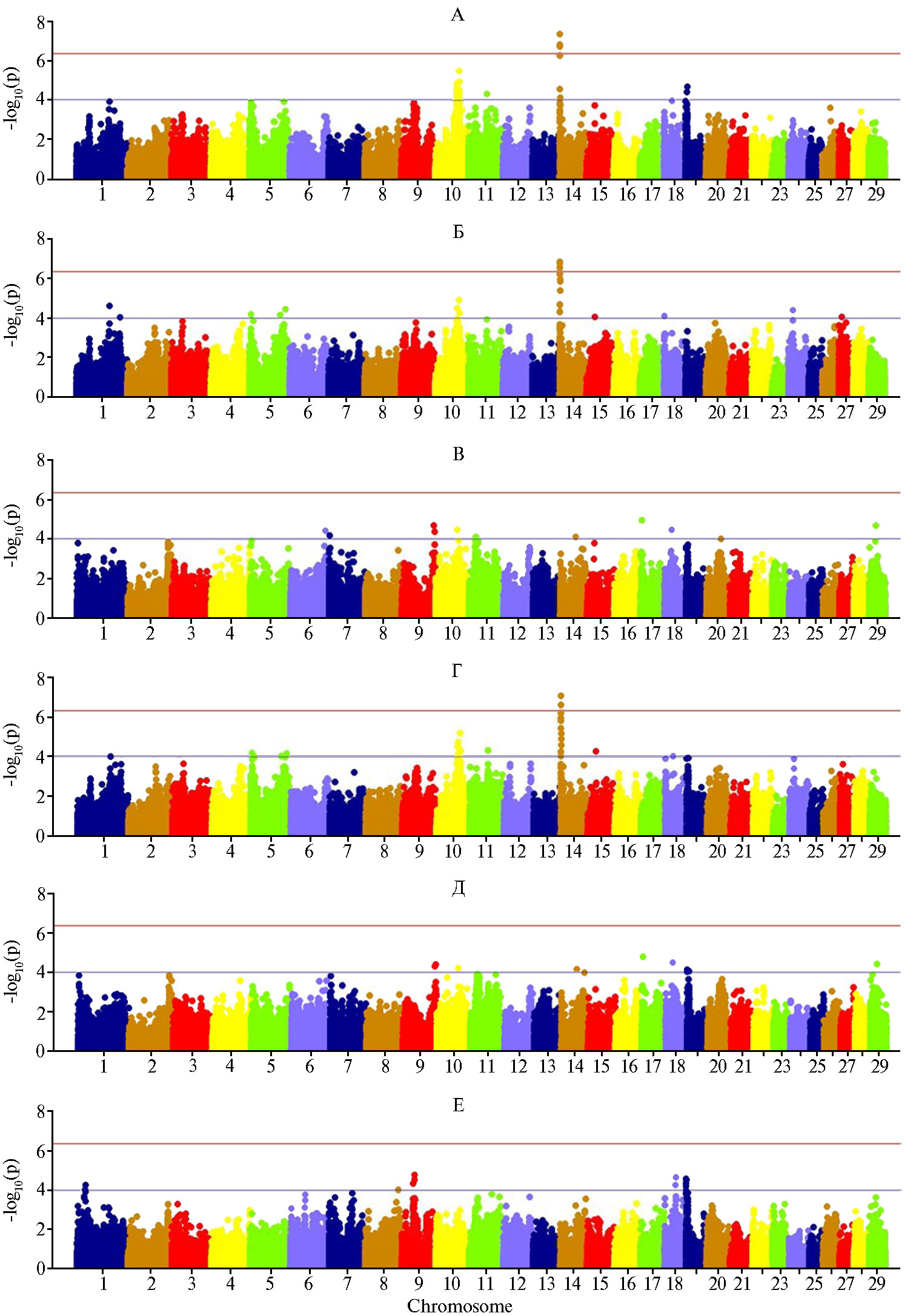
Рис. 3. Результаты GWAS анализа для короткоцепочечных жирных кислот (А), среднецепочечных
жирных кислот (Б), длинноцепочечных жирные жирных кислот (В), насыщенных жирных кислот (Г), мононенасыщенных жирных кислот (Д) и полиненасыщенных жирных кислот (Е) на основе прямых фенотипических показателей (n = 144, экспериментальное стадо голштинизированных черно-пестрых коров, ПЗ «Ладожский», Краснодарский край, 2020-2021 годы).
ЛИТЕРАТУРА
- Сермягин А., Зиновьева Н., Ермилов А., Янчуков И. Инфракрасная спектроскопия молока новые возможности в селекции и менеджменте стада. Животноводство России, 2019, S1: 65-68 (doi: 10.25701/ZZR.2019.17.64.008).
- Апполонова И.А., Смирнова Е.А., Никанорова Н.П. Исследования жирнокислотного состава липидов молока. Пищевая промышленность, 2012, 11: 72-45.
- Лашнева И.А., Сермягин А.А. Влияние наличия транс-изомеров жирных кислот в молоке на его состав и продуктивность коров. Достижения науки и техники АПК, 2020, 3: 46-50 (doi: 10.24411/0235-2451-2020-10309).
- Soyeurt H. Variation in fatty acid contents of milk and milk fat within and across breeds. Journal of Dairy Science, 2006, 89(12): 4858-4865 (doi: 10.3168/jds.S0022-0302(06)72534-6).
- DePeters E.J. Fatty acid and triglyceride composition of milk fat from lactating Holstein cows in response to supplemental canola oil. Journal of Dairy Science, 2001, 84(4): 929-936 (doi: 10.3168/jds.S0022-0302(01)74550-X).
- Collomb M. Impact of a basal diet of hay and fodder beet supplemented with rapeseed, linseed and sunflowerseed on the fatty acid composition of milk fat. International Dairy Journal, 2004, 14(6): 549-559 (doi: 10.1016/j.idairyj.2003.11.004).
- ГОСТ Р 52253-2004 «Масло и паста масляная из коровьего молока. Общие технические условия». М., 2005.
- Vanlierde A., Vanrobays M.-L., Dehareng F., Froidmon E., Soyeurt H., McParland S., Lewis E., Deighton M.H., Grandl F., Kreuzer M., Gredler B., Dardenne P., Gengler N. Hot topic: Innovative lactation-stage-dependent prediction of methane emissions from milk mid-infrared spectra. Journal of Dairy Science, 2015, 98(8): 5740-5747 (doi: 10.3168/jds.2014-8436).
- Shetty N., Difford G., Lassen J., Løvendahl P., Buitenhuis A.J. Predicting methane emissions of lactating Danish Holstein cows using Fourier transform mid-infrared spectroscopy of milk. Journal of Dairy Science, 2017, 100(11): 9052-9060 (doi: 10.3168/jds.2017-13014).
- Fragomeni B.O., Lourenco D.A.L., Masuda Y., Legarra A., Misztal I. Incorporation of causative quantitative trait nucleotides in single‑step GBLUP. Genetis Selection Evolution, 2017, 49: 59 (doi: 10.1186/s12711-017-0335-0).
- Смарагдов М.Г. Анализ расположения локусов, влияющих на показатели молока, в хромосомах крупного рогатого скота. Генетика, 2008, 44(6): 829-834.
- Смарагдов М.Г. Связь полиморфизма гена DGAT1 у быков-производителей с молочной продуктивностью коров. Генетика, 2011, 47(1): 126-132.
- Смарагдов М.Г. Исследование связи аллелей генов рецептора гормона роста и рецептора пролактина у быков-производителей с молочной продуктивностью их дочерей. Генетика, 2012, 48(9): 1085-1090.
- Sermyagin A.A., Gladyr’ E.A., Kharitonov S.N., Ermilov A.N., Strekozov N.I., Brem G., Zinovieva N.A. Genome-wide association study for milk production and reproduction traits in Russian Holstein cattle population. Sel’skokhozyaistvennaya biologiya [Agricultural Biology], 2016, 51(2): 182-193 (doi:10.15389/agrobiology.2016.2.182eng).
- Weller J.I., Ezra E., Ron M. Invited review: A perspective on the future of genomic selection in dairy cattle. Journal of Dairy Science, 2017, 100(11): 8633-8644 (doi: 10.3168/jds.2017-12879).
- Buitenhuis B, Janss LL, Poulsen NA, Larsen LB, Larsen MK, Sørensen P. Genome-wide association and biological pathway analysis for milk-fat composition in Danish Holstein and Danish Jersey cattle. BMC Genomics, 2014, 15(1): 1112 (doi: 10.1186/1471-2164-15-1112).
- Kawaguchi F., Kigoshi H., Fukushima M., Iwamoto E., Kobayashi E., Oyama K., Mannen H., Sasazaki S. Whole-genome resequencing to identify candidate genes for the QTL for oleic acid percentage in Japanese Black cattle. Animal Science Journal, 2019, 90(4): 467-472 (doi: 10.1111/asj.13179).
- Palombo V., Milanesi M., Sgorlon S., Capomaccio S., Mele‖ M., Nicolazzi E., Ajmone-Marsan P., Pilla F., Stefanon B., D'Andrea D. Genome-wide association study of milk fatty acid composition in Italian Simmental and Italian Holstein cows using single nucleotide polymorphism arrays. Journal of Dairy Science, 2018, 101(12): 11004-11019 (doi: 10.3168/jds.2018-14413).
- LI C., Sun D., Zhang S., Wang S., Wu X., Zhang Q., Liu L., Li Y., Qiao L. Genome wide association study identifies 20 novel promising genes associated with milk fatty acid traits in Chinese Holstein. PLoS ONE, 2014, 9(5): e96186 (doi: 10.1371/journal.pone.0096186)
- Gottardo P., Tiezzi F., Penasa M., Toffanin V., Cassandro M., De Marchi M. Milk fatty acids predicted by midinfrared spectroscopy in mixed dairy herds. Agricultural Conspectus Scientificus, 2013, 78(3): 263-266.
- Misztal I., Tsuruta S., Lourenço D., Aguilar I., Legarra A., Vitezica Z. Manual for BLUPF90 family of programs. Athens, University of Georgia, 2014.
- Masuda Y. Introduction to BLUPF90 suite programs. Standard Edition. University of Georgia, 2019.
- Chang C.C., Chow C.C., Tellier L.C.A.M., Vattikuti S., Purcell S.M., Lee J.J. Second-generation PLINK: rising to the challenge of larger and richer datasets. GigaScience, 2015, 4(1): s13742-015-0047-8 (doi: 10.1186/s13742-015-0047-8).
- Hu Z.L., Park C.A., Reecy J.M. Bringing the Animal QTLdb and CorrDB into the future: meeting new challenges and providing updated services. Nucleic Acids Research, 2022, 50(D1): D956-D961 (doi: 10.1093/nar/gkab1116).
- Sermyagin A.A., Lashneva I., Ignatieva L.P., Kositsin A., Gladyr E., Ermilov A., Yanchukov I., Zinovieva N.A. PSXI-3 genome-wide association study for MIR-predicted milk fatty acids composition in Russian Holstein cattle population. Journal of Animal Science, 2021, 99(Suppl_3): 245-246 (doi: 10.1093/jas/skab235.448).
- Huang H., Cao J., Hanif Q., Wang Y., Yu Y., Zhang S., Zhang Y. Genome-wide association study identifies energy metabolism genes for resistance to ketosis in Chinese Holstein cattle. Animal Genetics, 2019, 50(4): 376-380 (doi: 10.1111/age.12802).
- Schennink A., Stoop W.M., Visker M.H.P.W., Heck J.M.L., Bovenhuis H., Van Der Poel J.J., Van Valenberg H.J.F., Van Arendonk J.A.M. DGAT1 underlies large genetic variation in milk-fat composition of dairy cows. Animal Genetics, 2007, 38(5): 467-473 (doi: 10.1111/j.1365-2052.2007.01635.x).
- Bovenhuis H., Visker M.H.P.W., Poulsen N.A., Sehested J., Van Valenberg H.J..F, Van Arendonk J.A.M., Larsen L.B., Buitenhuis A.J. Effects of the diacylglycerol o-acyltransferase 1 (DGAT1) K232A polymorphism on fatty acid, protein, and mineral composition of dairy cattle milk. Journal of Dairy Science, 2016, 99(4): 3113-3123 (doi: 10.3168/jds.2015-10462).
- Ilie D.E., Mizeranschi A.E., Mihali C.V., Neamț R.I., Goilean G.V., Georgescu O.I., Zaharie D., Carabaș M., Huțu I. Genome-wide association studies for milk somatic cell score in Romanian dairy cattle. Genes (Basel), 2021, 12(10): 1495 (doi: 10.3390/genes12101495).
- Tiplady K.M., Lopdell T.J., Sherlock R.G., Johnson T.J.J., Spelman R.J., Harris B.L., Davis S.R., Littlejohn M.D., Garrick D.J. Comparison of the genetic characteristics of directly measured and Fourier-transform mid-infrared-predicted bovine milk fatty acids and proteins. Journal of Dairy Science, 2022, 105(12): 9763-9791 (doi: 10.3168/jds.2022-22089).