doi: 10.15389/agrobiology.2020.6.1204rus
УДК 636.2.034:636.087.8:579.6:577.2
Работа выполнена при поддержке РФФИ (грант № 18-016-00207).
ТАКСОНОМИЧЕСКАЯ И ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА МИКРОБИОТЫ РУБЦА ЛАКТИРУЮЩИХ КОРОВ ПОД ВЛИЯНИЕМ ПРОБИОТИКА ЦЕЛЛОБАКТЕРИНА+
Е.А. ЙЫЛДЫРЫМ1 ✉, Г.Ю. ЛАПТЕВ1, Л.А. ИЛЬИНА1, Т.П. ДУНЯШЕВ1, Д.Г. ТЮРИНА1, В.А. ФИЛИППОВА1,
Е.А. БРАЖНИК1, Н.В. ТАРЛАВИН1, А.В. ДУБРОВИН1, Н.И. НОВИКОВА1, В.В. СОЛДАТОВА1, С.Ю. ЗАЙЦЕВ2
В настоящее время рационы дойных коров построены так, чтобы обеспечить максимальную скорость роста и продуктивность за короткий промежуток времени. Однако интенсивное ведение животноводства сказывается в первую очередь на здоровье животных, поскольку нарушаются свойственные жвачным пути обмена веществ. Использование подходов 16S-метагеномики позволяет оценить генетическое и метаболическое разнообразие микробиома коров и установить факторы, которые способствуют увеличению продуктивности и улучшению состояния здоровья. В настоящей работе с применением программного комплекса PICRUSt2 и MetaCyс впервые установлен факт усиления предсказанного функционального потенциала некоторых метаболических путей в микробиоме рубца коров, в рацион которых вводили штамм бактерии Enterococcus faecium 1-35 в составе пробиотика Целлобактерина+. Целью работы была оценка влияния Целлобактерина+ на зоотехнические показатели, состав микробиома рубца и функциональный потенциал микробиоты при введении препарата в рацион дойных коров. Эксперимент проводили в зимне-весенний период 2018 года на базе АО ПЗ «Пламя» (Ленинградская обл., Гатчинский р-н). По принципу аналогов были сформированы две группы (по 10 гол.) дойных коров (Bos taurus taurus) голштинизированной черно-пестрой породы 2-й и 3-й лактации со средним годовым удоем 7000-7500 кг. Пробиотик Целлобактерин+ (ООО «БИОТРОФ», г. Санкт-Петербург) вводили в рацион коров опытной группы из расчета 40 г/гол. Продолжительность опыта — 60 сут. Отбор проб рубцового содержимого (10-50 г) от трех коров из каждой группы проводили в конце эксперимента с использованием стерильного зонда. Параллельно отбирали натощак кровь для биохимического анализа из подхвостовой вены. Также определяли массовую долю жира, белка и число соматических клеток в молоке. Тотальную ДНК из исследуемых образцов выделяли с использованием набора Genomic DNA Purification Kit («Fermentas, Inc.», Литва). Амплификацию для последующего NGS-секвенирования (Veriti Thermal Cycler, «Life Technologies, Inc.», США) с эубактериальными праймерами (IDT) 343F (5´-CTCCTACGGRRSGCAGCAG-3´) и 806R (5´-GGACTACNVGGGTWTCTAAT-3´), фланкирующими участок V1V3 гена 16S рРНК.Метагеномное секвенирование (система MiSeq, «Illumina, Inc.», США) осуществляли с набором MiSeq Reagent Kit v3 («Illumina, Inc.», США). Таксономическую принадлежность микроорганизмов до рода определяли в программе RDP Classifier. Рассчитывали индекс α-биоразнообразия микробиома рубца Сhao1. Анализ микробного β-разнообразия выборок методом главных компонент проводили по методике Weighted UniFrac PCoA Emperor с использованием программного пакета QIIME. Реконструкцию и прогнозирование функционального содержания метагенома, семейств генов, ферментов осуществляли при помощи программного комплекса PICRUSt2 (v.2.3.0). Для анализа метаболических путей и ферментов пользовались базой данных MetaCyc (https://metacyc.org/). Скармливание пробиотика оказало достоверное влияние (р = 0,049) на повышение надоев, а также на снижение (р = 0,003) числа соматических клеток в молоке коров (на 38 тыс. · мл-1 · гол.-1). С помощью NGS-секвенирования была дана полная таксономическая и функциональная характеристика рубцовой микробиоты, включая некультивируемых представителей. Выявлены существенные различия между группами по 13 бактериальным родам. В частности, в рубце коров, получавших пробиотик Целлобактерин+ по сравнению контрольной группой снижалась доля представителей порядка Clostridia — бактерий родов Anaerofilum sp. (в 2,3 раза ниже, р ≤ 0,05) и Anaerostipes sp. (в 1,8 раза ниже, р ≤ 0,05), образующих в рубце лактат в качестве конечного продукта метаболизма глюкозы. В рубце животных, в рацион которых вводили Целлобактерин+, уменьшалась доля бактерий родов Campylobacter, Gemella, Mycoplasma, Shewanella (р ≤ 0,05) и Fusobacterium (в том числе F. necrophorum) (p ≤ 0,001), среди которых нередко встречаются патогены. Изменения в таксономической структуре микробиоты рубца под воздействием биопрепарата были связаны с метаболическими сдвигами. Предсказанный функциональный потенциал семи метаболических путей оказался усилен у коров из опытной группы по сравнению с контрольной. Так, при интродукции пробиотического штамма бактерий в 3,5 раза (р ≤ 0,05) увеличивались прогнозируемые метаболические возможности микробиома, связанные с синтезом глиоксилата из аллантоина, и в 2,3 раза (p ≤ 0,05) — связанные с биосинтезом высокоценного для жвачных пропионата из L-глутамата. Полученные данные позволяют предположить важную роль биопрепарата для поддержания гомеостаза метаболических процессов.
Ключевые слова: биопрепараты, Целлобактерин+, лактирующие коровы, рубец, 16S-метагеномика, NGS-секвенирование, метаболизм.
Жвачные занимают особое место среди других сельскохозяйственных животных в связи с уникальными особенностями функционирования пищеварительного аппарата. В рубце обитает многочисленное микробное сообщество, состоящее из бактерий, архей, микромицетов, которое позволяет животному использовать лигноцеллюлозный материал и конвертировать небелковый азот в микробный белок для получения энергии и аминокислот (1, 2). При ферментации полисахаридов образуются короткоцепочечные (так называемые летучие) жирные кислоты (ЛЖК) — ацетат, бутират, пропионат и другие, которые абсорбируются через эпителий рубца и используются животным для поддержания метаболизма.
Рубец жвачных населен разнообразными бактериями, не поддающимися традиционному культивированию, поэтому часто бывает затруднительно сделать правильные выводы об их физиологии и функциях (3). В то же время эти микроорганизмы могут нести гены, определяющие значительную часть метаболического разнообразия микробиома, играть решающую роль в расщеплении некрахмалистых полисахаридов, белков, синтезе биологически активных веществ (БАВ), вступать в активные межмикробные взаимодействия и оказывать существенное воздействие на макроорганизм. Появление методов 16S-метагеномики позволило установить последовательности ДНК для всей совокупности микроорганизмов из природных источников и получить полную таксономическую и функциональную характеристику рубцовой микробиоты независимо от возможности культивирования микроорганизма (4). Высокопроизводительное секвенирование позволяет оценить генетическое и метаболическое разнообразие микробиома коров и выявить факторы, которые способствуют как экологическому балансу, так и здоровью хозяина (5).
Современные рационы дойных коров построены таким образом, чтобы обеспечить максимальную скорость роста и продуктивность за короткий промежуток времени (6, 7). Однако интенсивные технологии сказывается в первую очередь на здоровье животных, поскольку нарушает свойственные жвачным метаболические пути. Вследствие того, что рубцовые микроорганизмы служат практически единственным источником ферментов для расщепления растительных кормов, а также непосредственными участниками обмена веществ, нарушение структуры микробиоценоза рубца может приводить ко многим негативным последствиям. И наоборот, направленное изменение состава микробиоты рубца сопровождается позитивными сдвигами в продуктивности, качественных характеристиках молока, воспроизводстве, сроке хозяйственного использования, что может стать одним из ключевых факторов в увеличении эффективности молочного животноводства (8).
Из-за роли жвачных как продуцентов метана, выделяемого в атмосферу, основное внимание исследователей было сосредоточено на группе микроорганизмов-метаногенов (9, 10). Однако в настоящее время в связи с проблемами физиологии и питания высокопродуктивных животных существует потребность в продолжении работ по изучению структуры микробиома и метаболических путей, реализуемых рубцовой микробиотой.
Стратегии регуляции состава микробиома включают введение в рацион скота кормовых добавок (пробиотиков, пребиотиков, фитобиотиков и т.д.) (11-13). Позитивные эффекты от влияния пробиотиков на микробиом рубца связывают в основном с их положительным воздействием на пищеварительные процессы, особенно на расщепление целлюлозы и синтез микробных белков. Представитель дрожжей Saccharomyces cerevisiae — наиболее популярный вид, применяемый для интродукции в рубец (12). Что касается пробиотиков на основе бактерий, то, по мнению S. Fernández с соавт. (13), использование бактерий из рода Lactobacillus может стать альтернативой при лечении и профилактике ряда заболеваний, поражающих жвачных животных. Так, введение в рацион телят штаммов L. johnsonii TP1.1, L. reuteri TP1.3B, L. johnsonii TP1.6 и L. amylovorus TP8.7 снижало выраженность симптомов диареи (13).
Несмотря на то, что положительное влияние биодобавок на микробиом рубца коров достаточно хорошо известно, необходимость в изучении свойств существующих и новых пробиотических препаратов для животных остается высокой.
В настоящем исследовании с применением программного комплекса PICRUSt2 и MetaCyс впервые установлен факт усиления предсказанного функционального потенциала некоторых метаболических путей у микроорганизмов рубцовой микробиоты коров, в рацион которых вводили штамм бактерии Enterococcus faecium 1-35 в составе пробиотика Целлобактерина+.
Нашей целью была оценка влияния пробиотика Целлобактерина+ на зоотехнические показатели, состав рубцового микробиома и его функциональный потенциал у дойных коров.
Методика. Эксперимент проводили в зимне-весенний период 2018 года на базе АО ПЗ «Пламя» (Ленинградская обл., Гатчинский р-н). По принципу аналогов сформировали две группы (по 10 гол.) дойных коров (Bos taurus taurus) голштинизированной черно-пестрой породы 2-й и 3-й лактации со средним годовым удоем 7000-7500 кг молока. Животные находились в одинаковых привязных условиях содержания.
Основной рацион включал 10 кг комбикорма, 2 кг кукурузы желтой, 0,5 кг жмыха подсолнечного, 0,5 кг жмыха рапсового, 1 кг сена, 25 кг силоса из трав, 1 кг мелассы из свеклы, 0,2 кг МИНВИТ®-3 («АгроБалт трейд», Россия). Пробиотик Целлобактерин+ (ООО «БИОТРОФ», г. Санкт-Петербург), в состав которого входил штамм Enterococcus faecium 1-35, вводили в рацион коров опытной группы утром из расчета 40 г/гол. Дозировка была отработана нами ранее на дойных коровах (14). Продолжительность опыта составляла 60 сут, опыту предшествовал подготовительный период 15 сут.
Пробы рубцового содержимого (10-50 г) отбирали у трех коров из каждой группы в конце эксперимента вручную стерильным зондом, соблюдая условия асептики. Одновременно натощак отбирали кровь для биохимического анализа из подхвостовой вены, используя вакутейнеры. В сыворотке крови определяли общий белок, общий билирубин, глюкозу, кальций, фосфор, мочевину, резервную щелочность, кетоновые тела в соответствии с общепринятыми методиками (15). Массовую долю жира в молоке анализировали по ГОСТ 5867-90, белка — по ГОСТ 23327-98, количество соматических клеток — по ГОСТ Р 54761-2011.
Тотальную ДНК из образцов выделяли с использованием набора Genomic DNA Purification Kit («Fermentas, Inc.», Литва) согласно прилагаемой инструкции. Метод основан на селективном детергентно-опосредованном осаждении ДНК из субстрата с применением растворов для лизиса клеточных стенок и осаждения ДНК, 1,2 М хлорида натрия и хлороформа. Амплификацию (Veriti Thermal Cycler, «Life Technologies, Inc.», США) для последующего NGS-секвенирования проводили с эубактериальными праймерами (IDT) 343F (5´-CTCCTACGGRRSGCAGCAG-3´) и 806R (5´-GGACTACNVGGGTWTCTAAT-3´), фланкирующими участок V1V3 гена 16S рРНК. Режим амплификации: 3 мин при 95 °С (1 цикл); 30 с при 95 °С, 30 с при 55 °С, 30 с при 72 °С (25 циклов); 5 мин при 72 °С (1 цикл).
Метагеномное секвенирование (система MiSeq®, «Illumina, Inc.», США) выполняли с набором MiSeq Reagent Kit v3 («Illumina, Inc.», США). Максимальная длина полученных последовательностей составила 2×300 п.н. Химерные последовательности исключали из анализа с помощью программы USEARCH 7.0 (http://drive5.com/usearch/). Обработка полученных ридов с применением биоинформатической платформы CLC Bio GW 7.0 («Qiagen», Нидерланды) включала тестирование перекрываний, фильтрацию по качеству (QV > 15), триммирование праймеров. Таксономическую принадлежность микроорганизмов до рода определяли в программе RDP Classifier (http://rdp.cme.msu.edu/).
Рассчитывали индекс α-биоразнообразия микробиома рубца Сhao1 (16). Анализ микробного β-разнообразия выборок методом главных компонент проводили по методике Weighted UniFrac PCoA Emperor с использованием программного пакета QIIME (17). Реконструкцию и прогнозирование функционального содержания метагенома, семейств генов, ферментов осуществляли при помощи программного комплекса PICRUSt2 (v.2.3.0) (18). Для анализа метаболических путей и ферментов пользовались базой данных MetaCyc (https://metacyc.org/) (19). Профили метаболических путей MetaCyc оценивали после нормализации обилия ASV (Amplicon Sequence Variants), используя двоичный логарифм (log2) (18).
Математическую и статистическую обработку результатов проводили с применением программных пакетов Microsoft Office Excel 2003, R-Studio (Version 1.1.453) (https://rstudio.com). Определяли средние значения (М) и стандартные ошибки средних (±SEM). Результаты статистического анализа считались значимыми при p < 0,05.
Результаты. Введение в рацион коров пробиотика Целлобактерина+ не оказало достоверного влияния на содержание жира и белка в молоке, однако прослеживалась определенная тенденция позитивного воздействия препарата на эти показатели (табл. 1). В то же время скармливание Целлобактерина+ достоверно влияло (р = 0,049) на повышение надоев, а также на снижение (р = 0,003) числа соматических клеток в молоке коров (на 38 тыс.·мл-1·гол.-1). Ранее схожие результаты были получены J.S. Spaniol с соавт. (20) на дойных коровах: введение в рацион пробиотика не влияло на биохимический состав молока, но приводило к снижению числа соматических клеток на 15-е сут эксперимента. В работе австралийских авторов (21) среднесуточный надой молока коров, которые потребляли на пастбищах траву, обработанную пробиотическими штаммами бактерий, был на 1,21 л больше, чем у контрольных животных.
По результатам нескольких исследований, основанных на характеристике гена 16S рРНК, было выдвинуто предположение, что мастит крупного рогатого скота, сопровождающийся увеличением числа соматических клеток в молоке, становится результатом дисбаланса между нормобиотой молочной железы и патогенами (22). Возможно, уменьшение числа соматических клеток в молоке в нашем эксперименте было связано с модуляцией иммунной системы животного под воздействием пробиотического штамма бактерии. Ранее было продемонстрировано, что благодаря взаимодействию с моноцитами, макрофагами и дендритными клетками пробиотики могут модулировать баланс хелперных Т-клеток и таким образом оказывать влияние на адаптивный иммунный ответ (23, 24). Другими исследователями (20) показано, что введение пробиотика связано с ростом содержания циркулирующих цитокинов (фактора некроза опухоли, интерлейкина-4 и интерферона) в крови коров.
Практически все изученные нами биохимические показатели крови коров находились в пределах нормы или существенно не превышали их границ (табл. 2). Биохимические профили крови у коров контрольной и опытной групп не имели значимых различий.
На основании данных NGS-секвенирования были рассчитаны значения параметров α-биоразнообразия микробиома рубца (рис. 1), которое характеризовалось богатством представленности операционных таксономических единиц (ОТЕ) внутри сообществ (25, 26). Между опытным и контрольным вариантами не было отмечено значимых различий в числе ОТЕ и значениях индекса Chao1.
Результаты оценки β-разнообразия, то есть разнообразия между сообществами (25, 26), представлены в виде трехмерного графика PCoA Emperor (рис. 2). Главная компонента PC1 позволила описать 67,97 % данных, РС2 — 15,96 %, РС3 — 7,87 %, то есть в целом применение метода дало возможность охарактеризовать изменения, происходящие в составах микробиома, при сохранении 91,8 % информации. Сопоставление рубцовой микробиоты коров из разных групп методом главных компонент показало, что микробиомы трех коров из контрольной группы объединялись в один кластер, а микробиомы коров из опытной группы частично составляли свой кластер, что может свидетельствовать о влиянии пробиотика на структуру микробиома. Тем не менее кластерообразование в контрольной группе оказалось более выражено по сравнению с группой с введением пробиотика: смещение по оси первой главной компоненты РС1 было меньше.
По результатам оценки таксономической приуроченности микробиоты в рубце исследованных коров было обнаружено 27 филумов, из ко-торых доминирующими оказались Bacteroidetes (от 42,2±2,9 до 44,5±3,1 %), Proteobacteria (от 23,2±1,5 до 26,3±1,9 %), Firmicutes (от 16,3±0,9 до 17,2±1,2 %) (рис. 3). В филуме Bacteroidetes преобладали бактерии рода Prevotella (от 26,4±1,8 до 27,0±2,3 %). Ранее неоднократно было показано доминирование микроорганизмов этого рода в рубце жвачных (27, 28). Бактерии рода Prevotella играют важную роль в углеводном и азотном обмене, один из конечных продуктов их метаболизма — сукцинат (29). Установлено, что внеклеточный сукцинат в рубце служит основным предшественником пропионата (30) — важнейшего субстрата для процесса глюконеогенеза у жвачных у животных (31). Как было выявлено ранее на примере микробиома рубца овец, большая часть бактерий рода Prevotella представлена некультивируемыми формами (32).
Содержание в рубце бактерий, которые не принадлежали ни к одному известному таксону из существующих (по базам данных последовательностей гена 16S РНК), составляло от 6,9±0,5 до 7,5±0,8 % (см. рис. 3).
Значимых различий между вариантами на уровне филумов обнаружить не удалось (см. рис. 3). Однако детальный анализ состава микробиома рубца на уровне родов позволил выявить существенные различия между группами по 13 родам бактерий (рис. 4).
В частности, в рубце у коров, получавших Целлобактерин+, по сравнению с контрольной группой была детектирована более низкая доля представителей порядка Clostridia — бактерий родов Anaerofilum sp. (в 2,3 раза ниже, р ≤ 0,05) и Anaerostipes sp. (в 1,8 раза ниже, р ≤ 0,05). Представители этих родов образуют лактат в качестве конечного продукта метаболизма глюкозы (33, 34). Наши наблюдения могут указывать на позитивную роль пробиотика в контроле состояния здоровья коров, поскольку при высококонцентратном кормлении у животных нередко возникают дисбиотические нарушения рубцовой микрофлоры, сопровождающиеся смещением метаболизма в сторону образования лактата (35). Избыток лактата коррелирует с уменьшением pH рубца и лактатным ацидозом (35). На фоне ацидоза в результате подавления чувствительных к снижению рН продуцентов летучих жирных кислот, таких как Selenomonas ruminantium и Megasphaera elsdenii (36), скорость образования полезных метаболитов в рубце снижается. Аналогичным образом уменьшается и содержание бактерий, синтезирующих целлюлазы, что приводит к нарушению процессов переваривания некрахмалистых полисахаридов кормов (35). Наши результаты совпадают с данными H. Goto с соавт. (37), показавшими, что введение в рацион коров с индуцированным подострым ацидозом рубца мультиштаммового бактериального пробиотика вызывало уменьшение количества молочной кислоты в рубцовой жидкости.
В рубце животных, получавших пробиотик, мы обнаружили снижение представленности родов Campylobacter, Gemella, Mycoplasma, Shewanella (р ≤ 0,05) и Fusobacterium (p ≤ 0,001), среди которых нередко встречаются патогены. Данные об уменьшении обилия представителей родов Campylobacter и Fusobacterium у животных в опытной группе согласуются с приведенными выше результатами об уменьшении числа соматических клеток в молоке, поскольку доказано (38, 39), что эти микроорганизмы связаны с возникновением мастита у крупного рогатого скота. Наблюдаемое у коров контрольной группы увеличение в 60,5 раза (р ≤ 0,001) численности бактерий рода Fusobacterium, представленных преимущественно F. necrophorum, могло иметь связь с возрастанием обилия лактат-продуцирующих микроорганизмов в рубце. Дело в том, что низкие значения кислотности оптимальны для развития патогена F. necrophorum, для которого молочная кислота — основной питательный субстрат. F. necrophorum — это оппортунистический патоген, который вызывает некротические поражения рубца (некробактериозы), ламиниты, абсцессы печени (40). Присутствие бактерий рода Campylobacterв молоке может представлять опасность для людей, поскольку C. jejuni и С. coli способны инициировать желудочно-кишечный кампилобактериоз. Бактерии C. fetusассоциированы с бесплодием и абортированием у крупного рогатого скота (41).
Известно, что бактерии рода Gemella, содержание которых в результате применения Целлобактерина+ снизилось в 3,3 раза (р ≤ 0,05), связаны с инфекциями дыхательных путей и бактериемией (42). Аналогичным образом представители рода Mycoplasma, в частности M. bovis, становятся причиной хронических бронхопневмоний с казеозным и коагулятивным некрозом, а также артритов у крупного рогатого скота и телят (43). Бактерий рода Shewanella (S. haliotis и S. upenei) выделяли из легочной ткани людей с респираторной инфекцией и бактериемией (44). Увеличение пула легочных патогенов в рубце животных контрольной группы может свидетельствовать о связи между микробиомами, имеющими разную локализацию в организме хозяина, и существовании оси рубец—респираторный тракт, а также о возможном вмешательстве микробиома рубца в течение респираторных заболеваний. Ранее на примере крыс было показано, что фекальная трансплантация микробиома индуцировала изменения в легочной микробиоте (45).
Полученные данные указывают на роль пробитических бактериальных штаммов в поддержании не только гомеостаза микробиоты, но и здоровья макроорганизма.
Снижение обилия нежелательных форм микроорганизмов в результате воздействия пробиотика могло быть связано с прямым антагонизмом через продукцию антимикробных метаболитов (бактериоцинов, органических кислот) (46), а также с модуляцией состава и активности аборигенной микробиоты под воздействием штамма в составе биопрепарата. Так, в результате применения Целлобактерина+ в рубце увеличивалась численность бактерий родов Bifidobacterium и Bdellovibrio. Представители Bifidobacteriumшироко известны благодаря ярко выраженным антимикробным свойствам в отношении широкого круга патогенов (47). Представители рода Bdellovibrio, например B. bacteriovorus, — это хищные микроорганизмы, осуществляющие контроль таких патогенов, как Salmonella sp. и Escherichia coli (48).
Наши результаты, демонстрирующие модулирующий эффект, оказываемый пробиотиком на микрофлору, который выражался в снижении числа патогенных форм и увеличении обилия микроорганизмов с антимикробной активностью, перекликаются с данными, полученными на телятах (49). Применение болюсов на основе Pediococcus acidilactici, Enterococcus faecium, Lactobacillus acidophilus, Lactobacillus casei, Bifidobacterium bifidum способствовало снижению проявления диареи у животных.
Увеличение (р ≤ 0,05) в рубце коров из опытной группы численности бактерий семейств Lachnospiraceae (Pseudobutyrivibrio sp.и Lachnospira sp.) и Selenomonadaceae (Anaerovibrio sp.) могло также внести позитивный вклад в активацию метаболических процессов. Бактерии рода Pseudobutyrivibrio были представлены видом P. xylanivorans, который обладает мощной ксиланолитической ферментной системой, состоящей по меньшей мере из семи различных ксилангидролаз с молекулярной массой 27-145 кДа (50). В связи с этим он способен сбраживать полисахарид ксилан, содержащийся в кормах. Конечный продукт его метаболизма — летучие жирные кислоты, важные для обмена веществ, состояния здоровья и продуктивности животных, а также бактериоциноподобные ингибиторные вещества, активные в отношении патогенов. Микроорганизмы рода Lachnospira были представлены видом L. pectinoschiza, проявляющим ярко выраженную способность сбраживать пектин с помощью внеклеточной пектинметилэстеразы и Са2+-зависимой экзополигалактуронатлиазы (51). Конечный продукт метаболизма бактерий рода Lachnospira — уксусная кислота, которая служит основным субстратом для синтеза de novo липидов, в частности в молочных железах лактирующих коров.
Как показали результаты измерения скорости липолиза чистыми культурами Anaerovibrio sp., в том числе A. lipolytica (52), эти бактерии играют важную роль в липолитической активности содержимого рубца. Продуктами ферментации при этом становятся такие важные соединения, как пропионат, который образуется по пути превращения дикарбоновых кислот до сукцината. С усилением биосинтеза пропионата может быть ассоциировано увеличением молочной продуктивности коров, получавших пробиотик (30). Кроме того, короткоцепочечные жирные кислоты, продуцируемые бактериями, имеют ряд других важных свойств. Например, они вовлечены в эпигеномную регуляцию взаимодействий между микробиотой и макроорганизмом-хозяином (53). Давно известно, что эпигенетические модификации способны регулировать экспрессию генов, оказывая влияние на ее интенсивность и продолжительность, без изменений в основной последовательности ДНК.
Полученные в настоящем исследовании результаты закономерны, поскольку ранее мы описали механизмы позитивного влияния Целлобактерина+ на состав микробиоты рубца и кишечника (14). Эти механизмы выражаются в способности штаммов бактерий, входящих в состав биопрепарата, продуцировать низкомолекулярные органические кислоты и ряд других биологически активных веществ, включая антимикробные факторы. Мы доказали, что в результате синтеза ферментов биодеструкции ксенобиотиков в рубце происходит детоксикация микотоксинов корма, обладающих антимикробной активностью в отношении нормобиоты (14). Следовательно, появление новых метаболитов в рубце в результате интродукции пробиотического штамма приводит к изменениям в составе микроорганизмов.
С применением программного комплекса PICRUSt2 и MetaCyс мы провели реконструкцию и прогнозирование функционального содержания метагеномного сообщества рубца коров. Изменения в таксономической структуре микроорганизмов рубца под воздействием биопрепарата были связаны с метаболическими сдвигами. Предсказанный функциональный потенциал семи метаболических путей был усилен у коров, в рацион которых вводили Целлобактерин+ (рис. 5).
В частности, у коров из опытной группы по сравнению с контролем повышались (в 3,5 раз, р ≤ 0,05) прогнозируемые метаболические возможности микробиома, связанные с синтезом глиоксилата из аллантоина при участии ферментов аллантоиназы (ЕС 3.5.2.5), аллантоин-амидогидролазы (ЕС 3.5.3.9), уреидоглицин-аминогидролазы (ЕС 3.5.3.26), уреидогликолевой лиазы (ЕС 4.3.2.3). Исходный субстрат цикла — аллантоин, продукт катаболизма пуринов. Аллантоин богат азотом, и многие микроорганизмы способны его перерабатывать. Образующийся в результате превращений аллантоина глиоксилат служит субстратом для глиоксилатного цикла (цикла двууглеродных кислот). Принципиальная возможность функционирования глиоксилатного цикла в рубце связана с проявлением каталитической активности ключевых ферментов — изоцитратлиазы (ЕС 4.1.3.1) и малатсинтазы (ЕС 4.1.3.2) (54). Наличие этих ферментов позволяет синтезировать глюкозу, дефицитную для считающихся физиологически гипогликемичными жвачных, из уксусной кислоты, образующейся в высоких концентрациях в рубце. По сравнению с циклом трикарбоновых кислот путь окисления двууглеродных кислот энергетически более экономичен и эффективен, поскольку представляет собой более короткий цикл, способный функционировать как цикл трикарбоновых кислот, исключая лимитирующие реакции с участием изоцитратдегидрогеназы и α-кетоглутаратдегидрогеназы (54). Способность микроорганизмов желудочно-кишечного тракта крупного рогатого скота осуществлять глиоксилатный цикл можно рассматривать как фактор интенсификации обмена веществ и увеличения продуктивности. Еще один ответ на введение в рацион пробиотика Целлобактерина+, детектированный в результате биоинформатической обработки данных, — это активация (в 4,8 раза, р ≤ 0,01) превращения аллантоина через уреидогликолат в мочевину. Хорошо известно (55), что у жвачных животных эндогенная мочевина частично рециркулируется в организме и используется для синтеза высокоценного микробиального протеина, который всасывается в тонкой кишке хозяина.
У животных из опытной группы происходила активация (в 2,8 раза, р ≤ 0,05) потенциала микрофлоры рубца, связанного с синтезом γ-амино-Н-бутирата из L-орнитина, что ранее было подробно изучено S. Kurihara с соавт. (56). γ-Амино-бутират — главный ингибиторный нейротрансмиттер в центральной нервной системе млекопитающих, оказывает этиотропный эффект на состояние здоровья и скорость роста телят (57), обладает защитным действием против вызванной нейротоксикантами гибели клеток (58). Хорошо известно, что активно образуют γ-амино-Н-бутират из L-орнитина бактерии рода Bifidobacterium (59), обилие которых возрастало в нашем эксперименте в рубце животных, получавших пробиотик Целлобактерин+.
При введении в рацион Целлобактерина+ у коров повышались (в 2,3 раза, p ≤ 0,05) метаболические возможности микробиома, связанные с биосинтезом пропионата из L-глутамата. Этот путь был впервые описан для двух представителей семейства Veillonellaceae — Anaeromusa acidaminophila и Barkera propionica (60, 61). Бактерии семейства Veillonellaceae обнаружены нами в рубце коров контрольной и опытной групп, однако значимых различий по их содержанию выявить не удалось. Пропионовая кислота, включаясь в процесс глюконеогенеза, становится основным источником глюкозы в крови жвачных (62). Введение в рацион Целлобактерина+ способствовало также активации синтеза важного соединения — сукцината через L-аргинин, путресцин и γ-амино-Н-бутират. Cукцинат включается в цикл трикарбоновых кислот и служит главным предшественником пропионата, образующегося в рубце (30).
По сравнению с контрольной группой при использовании Целлобактерина+ увеличивались (в 2,8 раза, p ≤ 0,05) метаболические возможности микробиома, связанные с биосинтезом из фенилацетата сукцинил-СоА, представляющего собой тиоэфир дикарбоновой янтарной кислоты и кофермента. О существовании подобного пути у бактерий сообщалось еще в 1955 году (63). Деароматизация кольца достигается за счет превращения фенил-ацетил-CoA в 2-(1,2-эпокси-1,2-дигидрофенил)ацетил-СоА при участии фенилацетил-СоА 1,2-эпоксидазы (EC 1.14.13.149). Далее реактивный неароматический эпоксид изомеризуется до семичленного о-гетероциклического енольного эфира (2-оксепин-2(3Н)-илиденацетила-СоА), в результате чего кольцо расщепляется. Остальная часть пути состоит из β-окислительных стадий, приводящих к образованию сукцинил-СоА (64). Хорошо известно, что сукцинил-СоА принимает участие во многих биохимических путях, в частности служит интермедиатом цикла Кребса (65) и предшественником синтеза d-аминолевулиновой кислоты — специфического интермедиата в синтезе порфиринов.
Повышение прогнозируемых метаболических возможностей микробиома, связанных с синтезом глиоксилата, γ-амино-Н-бутирата, пропионата, мочевины, пептидогликана, сукцинил-CoA, сукцината, которое мы выявили у коров, получавших Целлобактерина+, подтверждает важную роль биопрепарата для поддержания гомеостаза метаболических процессов, здоровья и продуктивности животных. Это ценный научно-практический вывод, поскольку современные интенсивные методы ведения животноводства требуют включения в рацион значительного количества крахмала, что подвергает животное риску развития метаболических нарушений, возникновению заболеваний и снижению продуктивного долголетия. Объяснение полученных данных о повышении потенциала физиолого-биохимических процессов в рубце кров также требует углубленного анализа комплексных взаимовлияний микробиоты и макроорганизма. Целлобактерин+ также может быть полезен для иммуннобиологических воздействий при болезнях молочной железы, в том числе мастите, однако для подтверждения этого предположения необходимы дальнейшие испытания.
Таким образом, биопрепараты на основе микроорганизмов, эффективно модулирующие состав микробного сообщества, расширяют список средств для модификации структуры микробиома. В России после вступления в 2020 году в силу закона № 280-ФЗ от 3 августа 2018 года «Об органической продукции и о внесении изменений в отдельные законодательные акты Российской Федерации» интерес к таким натуральным добавкам резко возрос в связи с ограничением на применение антибиотиков (за исключением препаратов, разрешенных действующими в РФ национальными, межгосударственными и международными стандартами в сфере производства органической продукции).
Итак, введение в рацион дойных коров пробиотика Целлобактерина+ приводило к достоверному (р = 0,049) повышению надоев, а также к снижению числа соматических клеток в молоке (на 38 тыс.·мл-1·гол.-1, р = 0,003). По результатам NGS-секвенирования микробиоты рубца было установлено, что биопрепарат благотворно влиял на восстановление микробного сообщества. При этом детальный анализ состава микробиома рубца выявил существенные различия по 13 бактериальным родам. В частности, в рубце у коров, получавших Целлобактерин+, снизилась представленность родов бактерий Anaerofilum sp. (в 2,3 раза, р ≤ 0,05) и Anaerostipes sp. (в 1,8 раза, р ≤ 0,05), образующих лактат в качестве конечного продукта метаболизма глюкозы, и таксонов, среди которых нередко встречаются патогены, — Campylobacter, Gemella, Mycoplasma, Shewanella (р ≤ 0,05) и Fusobacterium (в том числе F. necrophorum) (p ≤ 0,001). Уменьшение числа соматических клеток в молоке было ассоциировано со снижением численности возбудителей мастита в рубце. На основании биоинформатической обработки данных мы подробно описали метаболические сдвиги у рубцовой микробиоты, происходящие на генном уровне в результате интродукции пробиотического штамма и изменения структуры микробиома. Предсказанный функциональный потенциал семи метаболических путей усиливался у коров, в рацион которых вводили Целлобактерин+. Интересным представляется дальнейшее изучение механизмов благоприятного воздействия интродуцированных бактерий на хозяина, в частности оценка жизнеспособности, адгезионного потенциала и выживаемости штамма бактерии в составе биопрепарата в условиях пищеварительного тракта.
1. Молочная продуктивность дойных коров (Bos taurus taurus) голштинизированной черно-пестрой породы при введении в рацион пробиотика Целлобактерина+ (M±SEM, АО ПЗ «Пламя», Ленинградская обл., Гатчинский р-н, 2018 год) |
|||
Показатель |
Контроль |
Опыт |
Значение р при сравнении групп |
Среднесуточный удой натурального молока, кг |
31,7±1,50 |
33,3±1,60 |
0,049 |
Массовая доля жира в молоке, % |
3,68±0,150 |
3,97±0,200 |
0,260 |
Массовая доля белка в молоке, % |
2,88±0,170 |
3,14±0,140 |
0,250 |
Среднесуточный удой молока с 4 % жирностью, кг |
29,2±1,20 |
33,0±1,40 |
0,048 |
Содержание соматических клеток, тыс.·мл-1·гол.-1 |
163±8,5 |
125±6,9 |
0,003 |
Примечание. Описание групп см. в разделе «Методика». |
|||
2. Биохимические показатели крови дойных коров (Bos taurus taurus) голштинизированной черно-пестрой породы при введении в рацион пробиотика Целлобактерина+ (M±SEM, АО ПЗ «Пламя», Ленинградская обл., Гатчинский р-н, 2018 год) |
||||
Показатель |
Контроль |
Опыт |
Значение р при сравнении групп |
Норма |
Белок общий, г/л |
78,3±4,10 |
81,8±4,90 |
0,62 |
70-89 |
Альбумин, % от общего белка |
50,7±2,90 |
40,9±2,00 |
0,07 |
38-50 |
Билирубин общий, ммоль/л |
2,2±0,20 |
2,33±0,110 |
0,6 |
0,17-5,13 |
Глюкоза, ммоль/л |
2,19±0,100 |
2,28±0,130 |
0,62 |
2,22-3,33 |
Кальций, ммоль/л |
2,28±0,140 |
2,37±0,190 |
0,73 |
2,6-3,5 |
Фосфор, ммоль/л |
2,52±0,200 |
2,09±0,100 |
0,07 |
1,29-2,25 |
Резервная щелочность, об.% СО2 |
57,8±2,30 |
55,0±2,30 |
0,45 |
46-56 |
Мочевина, ммоль/л |
4,4±0,30 |
3,77±0,150 |
0,16 |
3,3-6,7 |
Кетоновые тела |
– |
– |
|
– |
Примечание. Описание групп см. в разделе «Методика». Прочерки означают, что кетоновые тела не обнаружили. |
||||
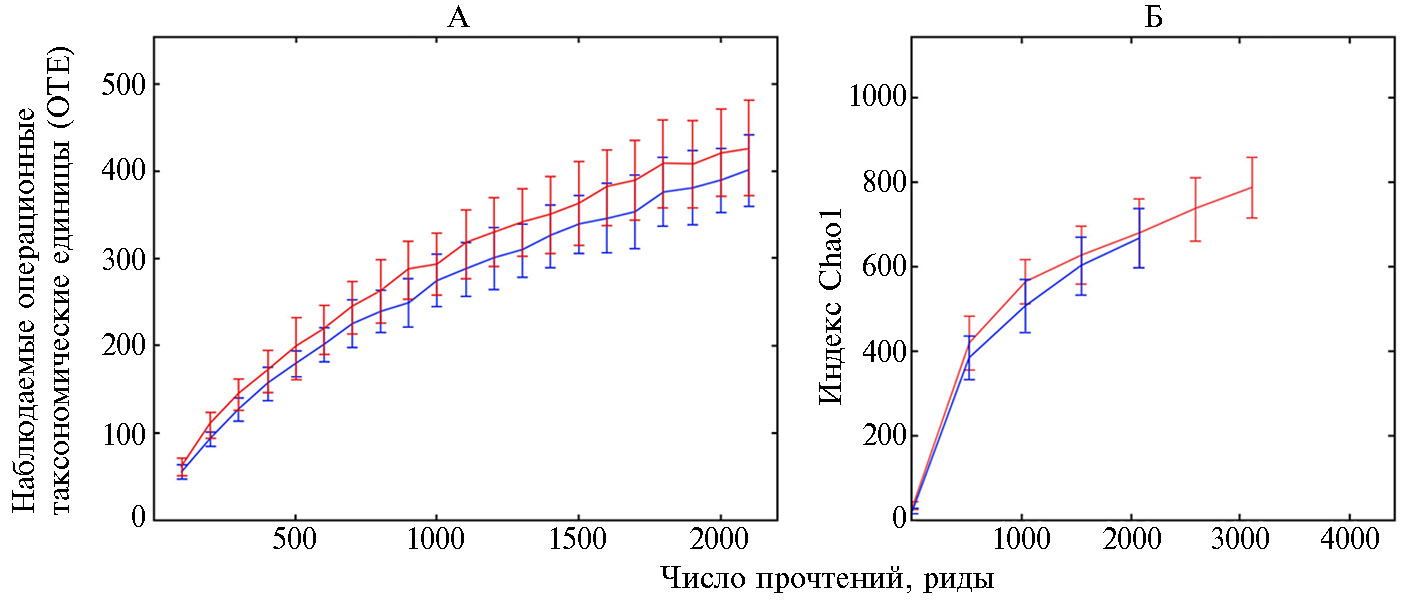
Рис. 1. Значения параметров α-биоразнообразия микробиома рубца у дойных коров (Bos taurus taurus) голштинизированной черно-пестрой породы в контроле (синий график) и при введении в рацион пробиотика Целлобактерина+ (красный график): А — разнообразие ОТЕ, Б — индекс Chao1 (M±SEM, АО ПЗ «Пламя», Ленинградская обл., Гатчинский р-н, 2018 год).
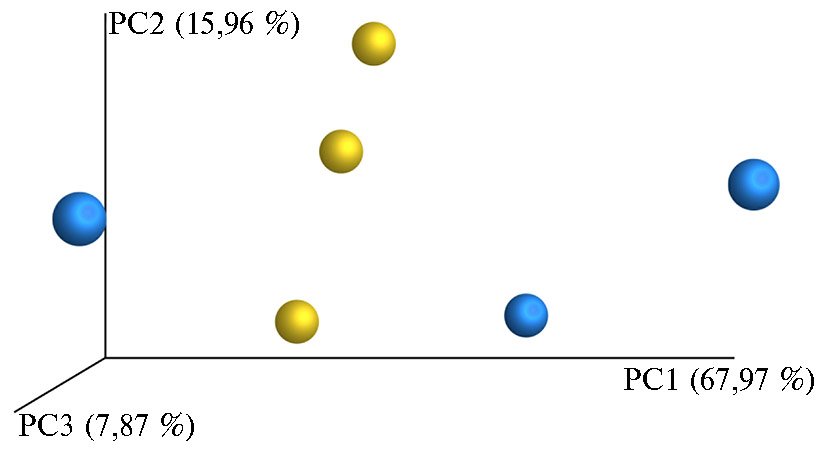
Рис. 2. Анализ β-разнообразия между микробиомами рубца у дойных коров (Bos taurus taurus) голштинизированной черно-пестрой породы методом главных компонент (одна точка соответствует одному животному) в контроле (желтые шары) и при введении в рацион пробиотика Целлобактерина+ (голубые шары) (АО ПЗ «Пламя», Ленинградская обл., Гатчинский р-н, 2018 год).
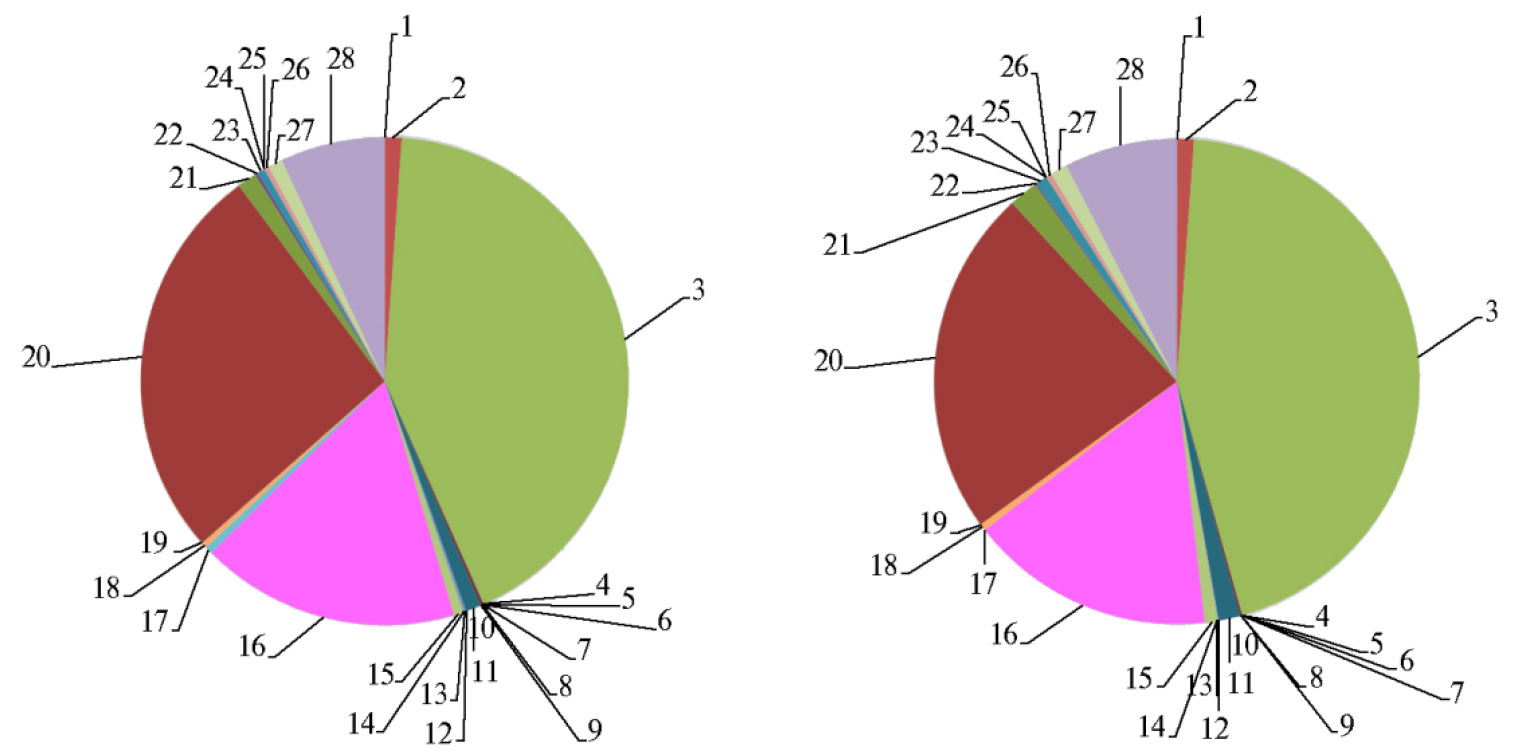
Рис. 3. Состав микроорганизмов на уровне филумов в рубце дойных коров (Bos taurus taurus) голштинизированной черно-пестрой породы в контроле (А) и при введении в рацион пробиотика Целлобактерина+ (Б) (по результатам NGS-секвенирования): 1 — Acidobacteria, 2 — Actinobacteria, 3 — Bacteroidetes, 4 — Caldiserica, 5 — Caldithrix, 6 — Chlamydiae, 7 — Chlorobi, 8 — Chloroflexi, 9 — Chrysiogenetes, 10 — Crenarchaeota, 11 — Cyanobacteria, 12 — Deferribacteres, 13 — Elusimicrobia, 14 — Euryarchaeota, 15 — Fibrobacteres, 16 — Firmicutes, 17 — Fusobacteria, 18 — Nitrospirae, 19 — Planctomycetes, 20 — Proteobacteria, 21 — Spirochaetes, 22 — Synergistetes, 23 — Tenericutes, 24 — Thermi, 25 — Thermodesulfobacteria, 26 — Thermotogae, 27 — Verrucomicrobia, 28 — неидентифицируемые (АО ПЗ «Пламя», Ленинградская обл., Гатчинский р-н, 2018 год).
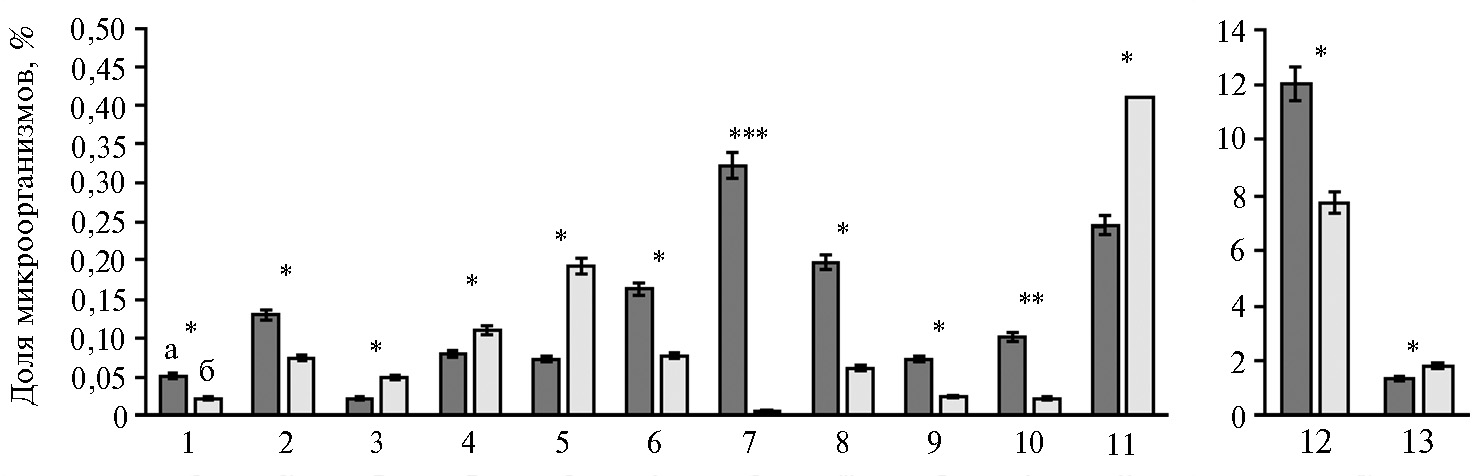
Рис. 4. Состав микроорганизмов на уровне родов в рубце дойных коров (Bos taurus taurus) голштинизированной черно-пестрой породы в контроле (а) и при введении в рацион пробиотика Целлобактерина+ (б) (по результатам NGS-секвенирования): 1 — Anaerofilum, 2 — Anaerostipes, 3 — Anaerovibrio, 4 — Bdellovibrio, 5 — Bifidobacterium, 6 — Campylobacter, 7 — Fusobacterium, 8 — Gemella, 9 — Mycoplasma, 10 — Odoribacter, 11 — Pseudobutyrivibrio, 12 — Shewanella, 13 — Lachnospira (M±SEM, АО ПЗ «Пламя», Ленинградская обл., Гатчинский р-н, 2018 год).
*, **, *** Различия между группами статистически значимы соответственно при p ≤ 0,05; p ≤ 0,01 и p ≤ 0,001.
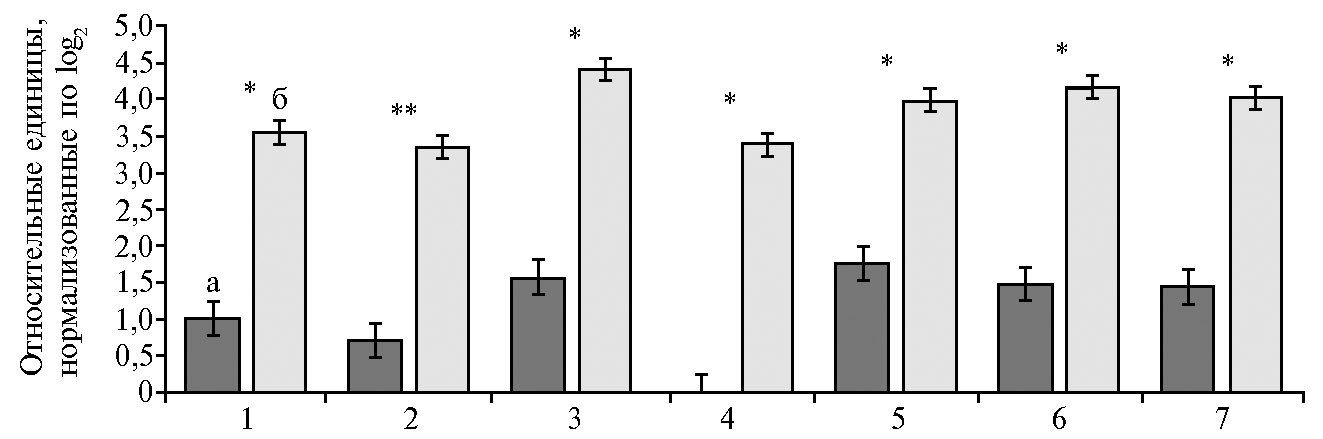
Рис. 5. Данные функциональной аннотации метаболических путей у метагеномного сообщества рубца дойных коров (Bos taurus taurus) голштинизированной черно-пестрой породы в контроле (а) и при введении в рацион пробиотика Целлобактерина+ (б) (получены с применением программного комплекса PICRUSt2 и MetaCyс): 1 — синтез глиоксилата, 2 — синтез мочевины, 3 — синтез γ-амино-Н-бутирата, 4 — синтез пептидогликана, 5 — синтез пропионата, 6 — синтез сукцинил-СоА, 7 — синтез сукцината (M±SEM, АО ПЗ «Пламя», Ленинградская обл., Гатчинский р-н, 2018 год).
*, ** Различия между группами статистически значимы соответственно при p ≤ 0,05 и p ≤ 0,01.
ЛИТЕРАТУРА
- Freetly H.C., Dickey A., Lindholm-Perry A.K., Thallman R.M., Keele J.W., Foote A.P., Wells J.E. Digestive tract microbiota of beef cattle that differed in feed efficiency. Journal of Animal Science, 2019, 98(2): skaa008 (doi: 10.1093/jas/skaa008).
- Wang L., Zhang K., Zhang C., Feng Y., Zhang X., Wang X., Wu G. Dynamics and stabilization of the rumen microbiome in yearling Tibetan sheep. Scientific Reports, 2019, 9: 19620 (doi: 10.1038/s41598-019-56206-3).
- Li F., Li C., Chen Y., Liu J., Zhang C., Irving B., Fitzsimmons C., Plastow G., Guan L.L. Host genetics influence the rumen microbiota and heritable rumen microbial features associate with feed efficiency in cattle. Microbiome, 2019, 7: 92 (doi: 10.1186/s40168-019-0699-1).
- Chaucheyras-Durand F., Ossa F. REVIEW: The rumen microbiome: Composition, abundance, diversity, and new investigative tools. The Professional Animal Scientist, 2014, 30(1): 1-12 (doi: 10.15232/S1080-7446(15)30076-0).
- Xue M.Y., Sun H.Z., Wu X.H., Liu J.-X., Gua L.L. Multi-omics reveals that the rumen microbiome and its metabolome together with the host metabolome contribute to individualized dairy cow performance. Microbiome, 2020, 8: 64 (doi: 10.1186/s40168-020-00819-8).
- Ильина Л.А. Содержание микроорганизмов в рубце телят разного возраста. Вестник мясного скотоводства, 2017, 3(99): 128-133.
- Schingoethe D.J. A 100-Year Review: Total mixed ration feeding of dairy cows. Journal of Dairy Science, 2017, 100(12): 10143-10150 (doi: 10.3168/jds.2017-12967).
- Uyeno Y., Shigemori S., Shimosato T. Effect of probiotics/prebiotics on cattle health and productivity. Microbes and Environments, 2015, 30(2): 126-132 (doi: 10.1264/jsme2.ME14176).
- Difford G.F., Plichta D.R., Løvendahl P., Lassen J., Noel S.J., Højberg O., Wright A.G., Zhu Z., Kristensen L., Nielsen H.B., Guldbrandtsen B., Sahana G. Host genetics and the rumen microbiome jointly associate with methane emissions in dairy cows. PLOS Genetics, 2018, 14(10): e1007580 (doi: 10.1371/journal.pgen.1007580).
- Matthews C., Crispie F., Lewis E., Reid M., O'Toole P.W., Cotter P.D. The rumen microbiome: a crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes, 2019, 10(2): 115-132 (doi: 10.1080/19490976.2018.1505176).
- Ghorbani G.R., Morgavi D.P., Beauchemin K.A., Leedle J.A.Z. Effects of bacterial direct-fed microbials on ruminal fermentation, blood variables, and the microbial populations of feedlot cattle. Journal of Animal Science, 2002, 80(7): 1977-1985 (doi: 10.2527/2002.8071977x).
- Uyeno Y., Shigemori S., Shimosato T. Effect of probiotics/prebiotics on cattle health and productivity. Microbes and Environments, 2015, 30(2): 126-132 (doi: 10.1264/jsme2.ME14176).
- Fernández S., Fraga M., Silveyra E., Trombert A.N., Rabaza A., Pla M., Zunino P. Probiotic properties of native Lactobacillus spp. strains for dairy calves. Beneficial Microbes, 2018, 9(4): 613-624 (doi: 10.3920/BM2017.0131).
- Лаптев Г.Ю., Новикова Н.И., Йылдырым Е.А., Ильина Л.А., Тарлавин Н.В. Микробиом сельскохозяйственных животных: связь со здоровьем и продуктивностью. СПб, 2020.
- Архипов А.В., Левченко В.И., Таланов Г.А., Фролова Л.А., Новиков В.Э. Методы ветеринарной клинической лабораторной диагностики. Справочник. М., 2004.
- Kim B.-R., Shin J., Guevarra R., Lee J.-H., Kim D.-W., Seol K.-H., Lee J.-H., Kim H.B., Isaacson R.E. Deciphering diversity indices for a better understanding of microbial communities. Journal of Microbiology and Biotechnology, 2017, 27(12): 2089‐2093 (doi: 10.4014/jmb.1709.09027).
- Vázquez-Baeza Y., Pirrung M., Gonzalez A., Knight R. EMPeror: a tool for visualizing high-throughput microbial community data. Gigascience, 2013, 2(1): 16 (doi: 10.1186/2047-217X-2-16).
- Douglas G.M., Maffei V.J., Zaneveld J.R., Yurgel S.N., Brown J.R., Taylor C.M., Huttenhower C., Langille M.G.I. PICRUSt2 for prediction of metagenome functions. Nature Biotechnology, 2020, 38: 685-688 (doi: 10.1038/s41587-020-0548-6).
- Caspi R., Billington R., Fulcher C.A., Keseler I.M., Kothari A., Krummenacker M., Latendresse M., Midford P.E., Ong Q., Ong W.K., Paley S., Subhraveti P., Karp P.D. The MetaCyc database of metabolic pathways and enzymes. Nucleic Acids Research, 2018, 46(D1): 633-639 (doi: 10.1093/nar/gkx935).
- Spaniol J.S., Oltramari C.E., Locatelli M., Volpato A., Campigotto G., Stefani L., Da Silva A.S. Influence of probiotic on somatic cell count in milk and immune system of dairy cows. Comparative Clinical Pathology, 2014, 24(3): 48-52 (doi: 10.1007/s00580-014-1966-y).
- Olchowy T.W.J., Soust M., Alawneh J. The effect of a commercial probiotic product on the milk quality of dairy cows. Journal of Dairy Science, 2019, 102(3): 2188-2195 (doi: 10.3168/jds.2018-15411).
- Oikonomou G., Machado V.S., Santisteban C., Schukken Y.H., Bicalho R.C. Microbial diversity of bovine mastitic milk as described by pyrosequencing of metagenomic 16s rDNA. PLoS One, 2012, 7(10): e47671 (doi: 10.1371/journal.pone.0047671).
- Lebeer S., Vanderleyden J., De Keersmaecker S.C. Host interactions of probiotic bacterial surface molecules: comparison with commensals and pathogens. Nature Reviews Microbiology, 2010, 8(3): 171-184 (doi: 10.1038/nrmicro2297).
- Klaenhammer T.R., Kleerebezem M., Kopp M.V., Rescigno M. The impact of probiotics and prebiotics on the immune system. Nature Reviews Immunology, 2012, 12(10): 728-734 (doi: 10.1038/nri3312).
- Розенберг Г.С. Информационный индекс и разнообразие: Больцман, Котельников, Шеннон, Уивер. Самарская Лука: проблемы региональной иглобальной экологии, 2010, 19(2): 4-25.
- Скопина М.Ю., Васильева А.А., Першина Е.В., Пиневич А.В. Разнообразие малочисленности: феномен “Разреженной бактериальной биосферы”. Микробиология, 2016, 85(3): 248-260.
- Gasparic A., Marinsek-Logar R., Martin J., Wallace R.J., Nekrep F.V., Flint H.J. Isolation of genes encoding β-D-xylanase, β-D-xylosidase and a-L-arabinofuranosidase activities from the rumen bacterium Prevotella ruminicola B14. FEMS Microbiology Letters, 1995, 125(2-3): 135-141 (doi: 10.1111/j.1574-6968.1995.tb07349.x).
- Stevenson D.M., Weimer P.J. Dominance of Prevotella and low abundance of classical ruminal bacterial species in the bovine rumen revealed by relative quantification real-time PCR. Applied Microbiology and Biotechnology, 2007, 75(1): 165-174 (doi: 10.1007/s00253-006-0802-y).
- Bryant M.P., Small N., Bouma C., Chu H. Bacteroides ruminicola n. sp. and Succinimonas amylolytica; the new genus and species; species of succinic acid-producing anaerobic bacteria of the bovine rumen. Journal of Bacteriology, 1958, 76(1): 15-23 (doi: 10.1128/JB.76.1.15-23.1958).
- Blackburn T.H., Hungate R.H. Succinic acid turnover and propionate production in the bovine rumen. Applied Microbiology and Biotechnology, 1963, 11(2): 132-135.
- Reynolds C.K., Huntington G.B., Tyrrell H.F., Reynolds P.J. Net metabolism of volatile fatty acids, D-β-hydroxybutyrate, nonesterified fatty acids, and blood gases by portal-drained viscera and liver of lactating Holstein cows. Journal of Dairy Science, 1988 71(9): 2395-2405 (doi: 10.3168/jds.S0022-0302(88)79824-0).
- Bekele A.Z., Koike S., Kobayashi Y. Genetic diversity and diet specificity of ruminal Prevotella revealed by 16S rRNA gene‐based analysis. FEMS Microbiology Letters, 2010, 305(1): 49-57 (doi: 10.1111/j.1574-6968.2010.01911.x).
- Zellner G., Stackebrandt E., Nagel D., Messner P., Weiss N., Winter J. Anaerofilum pentosovorans gen. nov., sp. nov., and Anaerofilum agile sp. nov., two new, strictly anaerobic, mesophilic, acidogenic bacteria from anaerobic bioreactors. International Journal of Systematic and Evolutionary Microbiology, 1996, 46(4): 871-875 (doi: 10.1099/00207713-46-4-871).
- Schwiertz A., Hold G.L., Duncan S.H. Anaerostipes caccae gen. nov., sp. nov., a new saccharolytic, acetate-utilising, butyrate-producing bacterium from human faeces. Systematic and Applied Microbiology, 2002, 25(1): 46-51 (doi: 10.1078/0723-2020-00096).
- Russell J.B., Sharp W.M., Baldwin R.L. The effect of pH on maximum bacterial growth rate and its possible role as a determinant of bacterial competition in the rumen. Journal of Animal Science, 1979, 48(2): 251-255 (doi: 10.2527/jas1979.482251x).
- Wang X., Li X., Zhao C., Hu P., Chen H., Liu Z., Liu G., Wang Z. Correlation between composition of the bacterial community and concentration of volatile fatty acids in the rumen during the transition period and ketosis in dairy cows. Applied and Environmental Microbiology, 2012, 78(7): 2386‐2392 (doi: 10.1128/AEM.07545-11).
- Goto H., Qadis A.Q., Kim Y.H., Ikuta K., Ichijo T., Sato S. Effects of a bacterial probiotic on ruminal pH and volatile fatty acids during subacute ruminal acidosis (SARA) in cattle. Journal of Veterinary Medical Science, 2016, 78(10): 1595-1600 (doi: 10.1292/jvms.16-0211).
- Neil S.D., Mackie D.P., Logan E.F. Campylobacter mastitis in dairy cows. Veterinary Record, 1982, 110(21): 505-506 (doi: 10.1136/vr.110.21.505).
- McGillivery D.J., Nicholls T.J., Hatch P.H. Isolation of Fusobacterium necrophorum from a case of bovine mastitis. Australian Veterinary Journal, 1984, 61(10): 325 (doi: 10.1111/j.1751-0813.1984.tb07141.x).
- Pillai D.K., Amachawadi R.G., Baca G., Narayanan S., Nagaraja T.G. Leukotoxic activity of Fusobacterium necrophorum of cattle origin. Anaerobe, 2019, 56: 51-56 (doi: 10.1016/j.anaerobe.2019.02.009).
- Hoffer M.A. Bovine campylobacteriosis: a review. Canadian Veterinary Journal, 1981, 22(11): 327-330.
- Jayananda S., Gollol-Raju N.S., Fadul N. Gemella species bacteremia and stroke in an elderly patient with respiratory tract infection. Case Reports in Medicine, 2017, 2017: 1098527 (doi: 10.1155/2017/1098527).
- Caswell J.L., Archambault M. Mycoplasma bovis pneumonia in cattle. Animal Health Research Reviews, 2007, 8(2): 161-186 (doi: 10.1017/S1466252307001351).
- Zhang F., Fang Y., Pang F. Rare Shewanella spp. associated with pulmonary and bloodstream infections of cancer patients, China: a case report. BMC Infectious Diseases, 2018, 18, 454 (doi: 10.1186/s12879-018-3354-8).
- Liu T., Yang Z., Zhang X., Han N., Yuan J., Cheng Y. 16S rDNA analysis of the effect of fecal microbiota transplantation on pulmonary and intestinal flora. 3Biotech, 2017, 7: 370 (doi: 10.1007/s13205-017-0997-x).
- Lebeer S., Bron P.A., Marco M.L., Van Pijkeren J.-P., O'Connell Motherway M., Hill C., Pot B., Roos S., Klaenhammer T. Identification of probiotic effector molecules: present state and future perspectives. Current Opinion in Biotechnology, 2018, 49: 217-223 (doi: 10.1016/j.copbio.2017.10.007).
- Lee D.K., Park S.Y., An H.M., Kim J.R., Kim M.Ji., Lee S.W., Cha M.K., Kim S.A., Chung M.J., Lee K.O., Ha N.J. Antimicrobial activity of Bifidobacterium spp. isolated from healthy adult Koreans against cariogenic microflora. Archives of Oral Biology, 2011, 56(10): 1047-1054 (doi: 10.1016/j.archoralbio.2011.03.002).
- Page J.A., Lubbers B., Maher J., Ritsch L., Gragg S.E. Investigation into the efficacy of Bdellovibrio bacteriovorus as a novel preharvest intervention to control Escherichia coli O157:H7 and Salmonella in cattle using an in vitro model. Journal of Food Protection, 2015, 78(9): 1745-1749 (doi: 10.4315/0362-028X.JFP-15-016).
- Renaud D.L., Kelton D.F., Weese J.S., Noble C., Duffield T.F. Evaluation of a multispecies probiotic as a supportive treatment for diarrhea in dairy calves: A randomized clinical trial. Journal of Dairy Science, 2019, 102(5): 4498-4505 (doi: 10.3168/jds.2018-15793).
- Čepeljnik T., Zorec M., Kostanjšek R., Nekrep F.V., Marinšek-Logar R. Is Pseudobutyrivibrio xylanivorans strain Mz5T suitable as a probiotic? An in vitro study. Folia Microbiologica, 2003, 48(3): 339-345 (doi: 10.1007/BF02931363).
- Cornick N.A., Jensen N.S., Stahl D.A., Hartman P.A., Allison M.J. Lachnospira pectinoschiza sp. nov., an anaerobic pectinophile from the pig intestine. International Journal of Systematic and Evolutionary Microbiology, 1994, 44(1): 87-93 (doi: 10.1099/00207713-44-1-87).
- Prins R.A., Lankhorst A., van der Meer P., Van Nevel C.J. Some characteristics of Anaerovibrio lipolytica a rumen lipolytic organism. Antonie Van Leeuwenhoek, 1975, 41(1): 1-11 (doi: 10.1007/BF02565031).
- Woo V., Alenghat T. Host-microbiota interactions: epigenomic regulation. Current Opinion in Immunology, 2017, 44: 52-60 (doi: 10.1016/j.coi.2016.12.001).
- Галочкина В.П., Агафонова А.В., Галочкин В.А. Гипотеза о специфической взаимосвязи пероксисомальных, митохондриальных и цитоплазматических процессов в регуляции обмена веществ у высокопродуктивных жвачных. Сельскохозяйственнаябиология, 2018, 53(2): 223-234 (doi: 10.15389/agrobiology.2018.2.223rus).
- Belasco I.J. New nitrogen feed compounds for ruminants — a laboratory evaluation. Journal of Animal Science, 1954, 13: 601-610.
- Kurihara S., Oda S., Kato K., Kim H.G., Koyanagi T., Kumagai H., Suzuki H. A novel putrescine utilization pathway involves gamma-glutamylated intermediates of Escherichia coli K-12. Journal of Biological Chemistry, 2005, 280(6): 4602-4608 (doi: 10.1074/jbc.M411114200).
- Matsumoto D., Takagi M., Fushimi Y., Okamoto K., Kido M., Ryuno M., Imura Y., Matsunaga M., Inokoshi K., Shahada F., Deguchi E. Effects of gamma-aminobutyric acid administration on health and growth rate of group-housed Japanese black calves fed using an automatic controlled milk feeder. Journal of Veterinary Medical Science, 2009, 71(5): 651-656 (doi: 10.1292/jvms.71.651).
- Cho Y.R., Chang J.Y., Chang H.C. Production of γ-aminobutyric acid (GABA) by Lactobacillus buchneri isolated from Kimchi and its neuroprotective effect on neuronal cells. World Journal of Microbiology and Biotechnology, 2007, 17(1): 104-109.
- Park K.B., Ji G.E., Park M.S., Oh S.H. Expression of rice glutamate decarboxylase in Bifidobacterium longum enhances gamma-aminobutyric acid production. Biotechnology Letters, 2005, 27(21): 1681-1684 (doi: 10.1007/s10529-005-2730-9).
- Nanninga, H.J., Drent W.J., Gottschal J.C. Fermentation of glutamate by Selenomonas acidaminophila sp. nov. Archives of Microbiology, 1987, 147(2): 152-157.
- Buckel W. Unusual enzymes involved in five pathways of glutamate fermentation. Applied Microbiology and Biotechnology, 2001, 57(3), 263-273 (doi: 10.1007/s002530100773).
- Kleiber M., Black A.L., Brown M.A., Tolbert B.M. Propionate as a precursor of milk constituents in the intact dairy cow. Journal of Biological Chemistry, 1953, 203(1): 339-346.
- Kunita N. Bacterial oxidation of phenylacetic acid. I. The pathway through homoprotocatechuic acid. Medical Journal of Osaka University, 1955, 6: 697-702.
- Teufel R., Mascaraque V., Ismail W., Voss M., Perera J., Eisenreich W., Haehnel W., Fuchs G. Bacterial phenylalanine and phenylacetate catabolic pathway revealed. Proceedings of the National Academy of Sciences USA, 2010, 107(32): 14390-14395 (doi: 10.1073/pnas.1005399107).
- Berg J.M., Tymoczko J.L., Stryer L. Biochemistry (5th ed.). NY, W.H. Freeman and Company, 2002.