








УДК 636.4+636.52/.58:577.161
ОБ УЧАСТИИ α-ТОКОФЕРИЛХИНОНА И УБИХИНОНА В РЕГУЛЯЦИИ ПРОЦЕССОВ РОСТА У СВИНЕЙ И ПТИЦЫ
В.И. ДУДИН, С.В. ГРИЩУК, Т.В. САКОВЦЕВА
В условиях вивария в опытах на свиньях — помесях пород ландрас и крупная белая, а также цыплятах кросса Гибро 6, полученного скрещиванием пород корниш и плимутрок, изучали влияние α-токоферилхинона, являющегося продуктом разрушения витамина Е в свободнорадикальных процессах in vivo, при добавлении в корм и внутримышечных инъекциях. В испытаниях обнаружили ростдепрессирующее действие изучаемого вещества и его функциональную связь с убихиноном.
Ключевые слова: свиньи, цыплята, свободнорадикальные процессы, ростдепрессирующее действие, убихинон, α-токоферилхинон, регуляция электронно-транспортной системы, радиоиндикаторный метод.
α-Токоферилхинон широко распространен в фотосинтезирующих водорослях и высших растениях (1). У растений он локализован в оболочке хлоропластов (2) и, по-видимому, участвует в транспорте электронов при фотосинтезе (3). α-Токоферилхинон найден также в микроорганизмах (4), хотя долгое время присутствие токоферола и его производных у них отрицалось (5).
Как видно, синтез α-токоферилхинона широко распространен в природе и его физиологическая роль, возможно, отличается от таковой a-токоферола (6). In vivo α-токоферилхинон может быть восстановлен в дыхательной цепи (7), при этом его восстановление не представляет собой часть классической митохондриальной электронно-транспортной системы, поскольку не ингибируется ротеноном или антимицином А. Из данных A. Bindoli с соавт. (8) вытекает, что a-токоферилхинон не способен заменять эндогенный убихинон, так как он частично ингибирует сукцинат-цитохром-c-редуктазную активность и окисление сукцината. Вместе с тем функциональная взаимосвязь между α-токоферилхиноном и убихиноном продолжает вызывать исследовательский интерес.
Целью нашей работы было выявление механизма регуляции ростдепрессирующего действия экзогенного α-токоферилхинона в исследованиях на свиньях и птице с применением высокоэффективной жидкостной хроматографии и радиоактивных индикаторов.
Методика. Эксперименты выполняли в условиях физиологического двора и радиохимического вивария Всероссийского НИИ физиологии, биохимии и питания сельскохозяйственных животных на свиньях — помесях пород ландрас и крупная белая и цыплятах кросса Гибро 6, полученного скрещиванием пород корниш и плимутрок.
Первый опыт проводили на двух группах поросят, сформированных по принципу аналогов (по 5 гол. в каждой) в возрасте 87 сут. Животные I группы (контроль) согласно установленным нормам (9) получали полнорационный комбикорм СК-5 с фоном природного a-токоферола 17,27, α-токоферилхинона — 6,00 мг/кг корма, II (опыт) — дополнительно α-токоферилхинон (50 мг/кг корма) (по данным анализа готовой кормосмеси). В опыте учитывали прирост живой массы, содержание α-токоферола и α-токоферилхинона в корме.
Второй опыт выполняли на двух группах поросят (по 3 гол. в каждой) (I — контроль, II — опыт) в период с 28- до 50-суточного возраста (первый вариант), а также на двух группах поросят (по 4 гол. в каждой) (I — контроль, II — опыт) в период с 40- до 48-суточного возраста (второй вариант). В течение последней недели перед убоем животным опытных групп внутримышечно инъецировали a-токоферилхинон (по 600 мг·гол-1·сут-1). Условия кормления и содержания животных в обоих вариантах были одинаковыми (полнорационный комбикорм СК-4, клеточно-групповое содержание). В первом и втором вариантах учитывали прирост живой массы, содержание α-токоферола и α-токоферилхинона в корме.
В третьем опыте изучали взаимосвязь между распределением ретинола, ретинилацетата, убихинона, α-токоферилхинона в печени А-гиповита-минозных свиней и временем их бодрствования после ночного сна. Животные (3 свинки и 3 боровка) со времени отъема в 30-суточном возрасте потребляли кормосмесь СК-3 без добавки ретинола. За 2 нед до убоя в рацион вводили d,l- α-токоферилацетат (250 мг/кг корма). Убой проводили в 7.00 (сразу после ночного сна), 9.00 и 11.00 (по 1 свинке или 1 боровку) в течение двух смежных суток (учет времени по свинкам и боровкам раздельный), готовили по одному продольному срезу правой медиальной части печени толщиной 1,5 см. Образцы для анализов отбирали в 16 точках среза. Определяли линейные размеры срезов, содержание убихинона, α-токоферилхинона, ретинола, ретинилацетата в тканях печени, содержание α-токоферола и α-токоферилхинона в корме.
Четвертый опыт проводили на 12 цыплятах-бройлерах в период с суточного до 10-суточного возраста. Условия кормления и содержания птицы соответствовали возрастным нормам (9). Птице (по 4 гол. для каждой возрастной группы) перорально однократно вводили D-α-[5-метил-3H]токоферол («Amersham», Великобритания): по 4,45 мкКu/гол. — 1- и 5-суточным цыплятам и по 8,90 мкКu/гол. — 10-суточным. Через 24 ч подопытных цыплят декапитировали. Из ткани печени и легких экстрагировали общие липиды, которые подвергали хроматографии на колонках размером 20 х 200 мм с силикагелем L (ЧССР) (размер частиц 40-100 мкм); элюировали смесью гексана, серного эфира и метанола с возрастающей полярностью (10). Радиоактивность фракций определяли с помощью сцинтилляционного счетчика Ultrobeta (Швеция). Тушение сцинтилляций учитывали авторским способом (11).
Измерение количества ретинола, его эфиров, убихинона и α-токоферилхинона в ткани печени, α-токоферола и α-токоферилхинона в кормах проводили с помощью высокоэффективной жидкостной хроматографии без омыления после ацетоновой экстракции образцов, обезвоженных с помощью сульфата натрия (11). Препарат α-токоферилхинона готовили окислением α-токоферола азотной кислотой (12) с последующей хроматографической очисткой на колонке с силикагелем, как описано выше (10), под контролем высокоэффективной жидкостной хроматографии (ВЭЖХ) (11). При приготовлении водорастворимой формы для инъекций его солюбилизировали Твином-80 (13).
Данные обрабатывали статистически по Е.К. Меркурьевой (14).
Результаты. Включение в корм поросят экзогенного α-токоферил-хинона в период с 87- по 105-суточный возраст (первый опыт) приводило к снижению среднесуточного прироста живой массы по сравнению с контролем (486,0±15,0 против 694,0±10,3 г/сут). Такой же эффект вызывало внутримышечное введение a-токоферилхинона (второй опыт, первый вариант) перед убоем в 50-суточном возрасте (на фоне содержания a-то-коферола в корме 270 мг/кг у поросят с 28-суточного возраста до убоя): 279,0±17,0 в опыте против 354,0±8,0 г/сут в контроле, то есть ростдепрессирующее действие экзогенного a-токоферилхинона было очень заметным (30 и 21 % по отношению к соответствующим контрольным значениям) и существенным (P < 0,05). В ранее проведенном подобном эксперименте, но на фоне обычного содержания витамина Е в корме поросят (33 мг/кг корма) такого эффекта экзогенный α-токоферилхинон не оказал (6). Во втором варианте (внутримышечное введение α-токоферилхинона поросятам в период с 40-х по 48-е сут жизни) приводило к резкому (более чем 2-кратному) снижению среднесуточного прироста живой массы (144,0 против 66,0 г/гол.).
|
Рис. 1.Взаимосвязь между длиной продольных срезов правых медиальных долей печени и средним содержанием α-токоферилхинона в тканях срезов у поросят — помесей пород ландрас и крупная белая (r = -0,98; P = 0,0001), в течение 3 нед получавших d,l-α-токоферилацетат в количестве 250 мг на 1 кг корма, (А) и корреляционная зависимость между концентрацией a-токоферилхинона и убихинона в тканях срезов у свинок (1) и боровков (2) в зависимости от времени бодрствования после ночного сна (Б) (ви- |
варий Всероссийского НИИ физиологии, биохимии и питания сельскохозяйственных животных, n = 16 на каждую точку). |
|
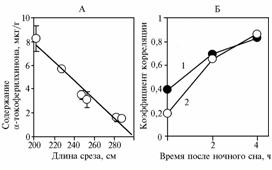
Для выяснения, связана ли интенсивность окисления α-токоферола в α-токоферилхинон в печени со степенью развития этого органа, в третьем опыте соотнесли среднее содержание α-токоферилхинона в 16 точках среза и длину этих срезов (в качестве характеристики размера органа) (рис. 1, А) и выявили обратную зависимость, что можно рассматривать как указание на способность α-токоферилхинона ингибировать рост животных на тканевом уровне. В этом опыте у животных в печени были обнаружены ретинол и ретиниловые эфиры (соответственно около 2 и 10 мкг/г), что свидетельствовало о выраженном А-гиповитаминозе. Известно, что дефицит витамина А у животных вызывает увеличение количества убихинона в печени, возможно, вследствие усиления его синтеза (15). По нашим данным, изменения усредненного содержания окисленного убихинона в основном соответствовали таковым α-токоферилхинона, причем корреляционная взаимосвязь между этими показателями (см. рис. 1, Б) зависела от времени убоя животных после ночного сна. Наименьший коэффициент корреляции отмечали у животных при убое сразу после ночного сна.
Как известно, состояние сна характеризуется пониженной метаболической активностью тканей, а следовательно, уменьшенным потреблением кислорода, в связи с чем можно предположить, что соотношение между содержанием a-токоферилхинона и убихинона в печени зависит от поступления кислорода в организм. При окислительном фосфорилировании 1 % O2 восстанавливается до супероксиданиона — О2-·. В митохондриях супероксид в основном образуется за счет неферментативного окисления семиубигидрохинона Uq10-·: Uq10-· + O2 -> О2-· + Uq10 (16).
Согласно современным представлениям, в свободнорадикальных патологиях повреждающее действие обусловлено не столько супероксидом О2-·, сколько продуктами его дальнейшего преобразования — преимущественно перекисью водорода (H2O2). В отличие от супероксиданиона она может свободно проникать через клеточные мембраны и, кроме того, способна участвовать не только в окислительных, но и в восстановительных реакциях (17):
|
Рис. 2. Профиль элюции радиоактивной метки при колоночной хроматографии общих липидов легких (А) и печени (Б) 10-суточных цыплят кросса Гибро 6 через 24 ч после однократного перорального введения [3H]α-токоферола в растворе растительного масла: 1 — |
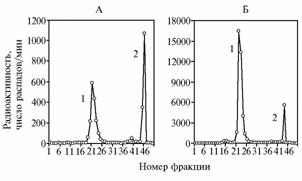
Перекись водорода образуется при двухэлектронном восстановлении кислорода, что происходит в реакциях с участием оксидазы L-аминокис-лот, альдегиддегидрогеназы, 5-монооксигеназы, супероксиддисмутазы. Активность каталазы наблюдается почти во всех животных клетках, где она предотвращает аккумуляцию токсичной перекиси водорода, образующейся при аэробном окислении восстановленных флавопротеидов и разрушении супероксиданиона. Поскольку в физиологических условиях отмечается высокое содержание восстановленных кофакторов и низкое — H2O2, очевидно, что в животных тканях каталаза действует в основном как пероксидаза, в качестве кофактора здесь может выступать восстановленный, в том числе в дыхательной цепи, α-токоферилхинон. В наших опытах интенсивность окисления α-токоферола в α-токоферилхинон была заметно более высокой в легких (рис. 2, А), причем далее большие количества α-токоферилхинона восстанавливались в тканях печени и легких в α-токоферилхинол. Абсолютные показатели интенсивности восстановления α-токоферилхинона в α-токоферилхинол через 24 ч после однократного перорального введения меченого [3H]α-токоферола в печени (см. рис. 2, Б) значительно превышали таковые в легких. По данным А. Уайта с соавт. (18), у крыс потребление кислорода в печени почти в 2 раза больше, чем в легких (15 против 8 мкл/мг сухой массы в час). В наших опытах (см. рис. 2) легкие также характеризовались значительным увеличением интенсивности восстановления a-токоферилхинона по сравнению с печенью.
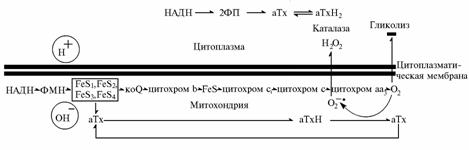 |
Рис. 3. Предлагаемая схема участия a-токоферилхинона в процессах окислительного фосфорилирования, происходящих в митохондриях аэробных эукариотических клеток: НАДН — никотинамидадениндинуклеотид восстановленный, ФП — флавопротеид, ФМН — флавинмононуклеотид, Ух — убихинон, аТх — α-токоферилхинон, аТхН2 — α-токоферилхинон восстановленный, FeS — железосерные центры. |
Одно из возможных метаболических различий между тучными и худыми особями — степень сопряжения окислительного фосфорилирования с величиной соотношения между интенсивностью аэробного и анаэробного обмена (19). В наших исследованиях результаты, полученные при скармливании (первый опыт) или парентеральном введении (второй опыт) a-токоферилхинона, наводят на мысль, что причина отмеченного уменьшения скорости роста животных может заключаться в отведении части потока НАДН из цепи переноса электронов в митохондриях на некие иные нужды. Одно из наиболее вероятных следствий участия α-токоферилхинона в процессах, происходящих в митохондриях, — снижение степени вовлечения НАДН в окислительное фосфорилирование из-за конкуренции за НАДН-дегидрогеназу между α-токоферилхиноном и убихиноном (рис. 3). Ввиду хорошей проницаемости митохондриальных мембран для перекиси водорода вследствие свободной диффузии можно предположить участие a-токоферилхинола в качестве донора протонов в пероксидазных реакциях, в том числе за счет каталазы, находящейся вне митохондрий.
Известно, что рубцовая бактерия Butyrivibrio fibrisolvens биогидрогенизирует линолевую кислоту в два этапа с участием a-токоферилхинола на втором (13). Выделенный из бактерии и частично очищенный препарат фермента представляет собой флавопротеид и катализирует восстановление α-токоферилхинона в соответствующий хинол в присутствии НАДН со стехиометрией 1:1 (11). То есть восстановление α-токоферилхинона в аэробных и анаэробных условиях протекает при участии ферментов флавопротеидной природы.
Наши данные подтверждают, что регуляция массы тела животных в значительной мере зависит от работы дыхательной цепи митохондрий (19). При этом в природе имеет место независимый биосинтез α-токоферил-хинона в печени животных (7) и более тесная, чем у α-токоферола, связь a-токоферилхинона с процессами роста животных и растений (3, 11).
Таким образом, полученные результаты позволяют заключить, что механизм регуляции процессов роста a-токоферилхиноном состоит в шунтировании дыхательной цепи между НАДН-дегидрогеназой и молекулярным кислородом в процессе его конкуренции с убихиноном. При этом a-токофе-рилхинон восстанавливается в α-токоферилхинол, в результате чего восстановительные эквиваленты НАДН расходуются без синтеза АТФ и происходит спонтанное окисление a-токоферилхинола в щелочной среде митохондрий с интенсивностью, зависящей от концентрации гидроксильных ионов.
Л И Т Е Р А Т У Р А
1. B u c k e C., L e e c h R.M., H a l l o w a y M. e.a. The taxonomic distribution of plastoquinone and tocopherolquinone and their intracellular distribution in leaves of Vicia faba. Biochim. Biophys. Acta, 1966, 112: 19-34.
2. L i c h t e n t h a l e r H.K., P r e n z e i U., D o u c e R. e.a. Localization of prenylquinones in the envelope of spinach chloroplasts. Biochim. Biophys. Acta, 1981, 641: 99-105.
3. K r u k J., S t r z a l k a K. Occurence and function of a-tocopherol quinone in plants. J. Plant. Physiol., 1995, 145(4): 405-409.
4. R o u s s e a u B., D o s t a l L., R o s a z z a J.P.N. Biotransformations of tocopherols by Streptomyces catenulae. Lipids, 1997, 32(1): 79-84.
5. W h i s t a n c e G.R., D i l l o n J.F., T h r e l f a l l D.R. The nature, intergeneric description and biosynthesis of isoprenoid quinones and phenols in gramnegative bacteria. Biochem. J., 1969, 111: 461-72.
6. Г р и щ у к С.В. Метаболизм альфа-токоферола у поросят раннего возраста. Канд. дис. Боровск, 1995.
7. H u g h e s P.E., T o v e S.B. Synthesis of a-tocopherolquinone by the rat and its reduction by mitochondria. J. Biol. Chem., 1980a, 255(15): 7095-7097.
8. B i n d o l i A., V a l e n t e M., C a v a l i n i L. Inhibition of lipids peroxidation by a-tocopherolquinon and a-tocopherolhydroquinone. Biochem. Int., 1985, 10(5): 735-761.
9. К а л а ш н и к о в А.П., Щ е г л о в В.В., П е р в о в Н.Г. Нормы и рационы кормления сельскохозяйственных животных. Справ. пос. М., 2003.
10. Д у д и н В.И., Д в и н с к а я Л.М. Участие альфа-токоферола в синтезе кислых липидов в организме цыплят. С.-х. биол., 1986, 101-105.
11. Д у д и н В.И. Метаболизм витамина Е и связанных с ним биологически активных веществ. М., 2004.
12. R a o G.H.R., K e i c h T.P., W h i t e J.G. Preparation, separation and characterisation of vitamin E quinone. J. Chromat., 1980, 196: 506-511.
13. S k l a n D., D o n o g h u e S. Vitamin E response to high dietary vitamin A in the chick. J. Nutr., 1982, 112(4): 759-763.
14. М е р к у р ь е в а Е.К. Биометрия в животноводстве. М., 1964.
15. E d w i n E.E., B u n j a n J., G r e e n J. e.a. The effect of vitamin A on ubiquinone and ubichromenol in the rat and its relation the effective vitamin E. Brit. J. Nutrit., 1962, 16(1): 135-149.
16. Т у р п а е в К.Т. Активные формы кислорода и регуляция экспрессии генов. Биохимия, 2002, 67(3): 339-352.
17. С а м у и л о в В.Д., Б е з р я д н о в Д.В., Г у с е в М.В. и др. Н2О2 ингибирует рост цианобактерий. Биохимия, 1999, 64(1): 60-67.
18. У а й т А., Х е н д л е р Ф., С м и т Э. и др. Основы биохимии. М., 1981, т. 1.
19 Т е п п е р м е н Дж., Т е п п е р м е н Х.М. Физиология обмена веществ и эндокринной системы. Вводный курс. М., 1989.
ABOUT PARTICIPATION OF α-TOCOPHERYLQUINONE AND UBIQUINONE IN REGULATION OF GROWING PROCESSES IN PIGS AND POULTRY
V.I. Dudin, S.V. Grishchuk, T.V. Sakovtseva
In the conditions of vivarium in the experiments on the Landras x Big White pigs and chicken of the Gibro 6 cross the authors studied the effect of α-tocopherylquinone, as a product of vitamin E splitting in free radical reactions in vivo, after its addition to the feed and intramuscular introduction. During the tests the growth depressing effect of studied preparation and its functional connection with ubiquinone were detected.
Key words: pigs, chicks, free radical processes, growth-depressing action, ubiquinone, α-tocopherylquinone, electron transport system regulation, radioisotopic method.
ГНУ Всероссийский НИИ физиологии, |
Поступила в редакцию |
Оформление электронного оттиска
![]()
![]()




