doi: 10.15389/agrobiology.2022.5.882rus
УДК 633.72:581.19:575
Исследования проводятся при поддержке РНФ (проект № 22-16-00058).
ГЕНЕТИЧЕСКИЕ МЕХАНИЗМЫ БИОСИНТЕЗА КАТЕХИНОВ, КОФЕИНА И L-ТЕАНИНА У ЧАЙНОГО РАСТЕНИЯ Camellia sinensis (L.) Kuntze (обзор)
Л.С. МАЛЮКОВА1 ✉, Л.С. САМАРИНА1, 2, Н.В. ЗАГОСКИНА3
Катехины, кофеин и L-теанин — основные вторичные метаболиты чайного растения Camellia sinensis (L.) Kuntze. Им отводится ключевая роль в формировании вкусовых качеств, пищевой и лекарственной ценности чая (W.J.M. Lorenzo с соавт., 2016; Z. Yan с соавт., 2020). Кроме того, они вовлечены в регуляцию жизнедеятельности растений, в частности в процессы адаптации к неблагоприятным условиям (Y.S. Wang с соавт., 2012; L.G. Xiong с соавт., 2013; G.J. Hong с соавт., 2014). Перечисленным определяется интерес к физиолого-биохимическим и молекулярным механизмам продукции катехинов, кофеина и L-теанина, селекции на повышение их содержания в растении (R. Fang с соавт., 2017; Kong W с соавт., 2022), а также к изучению их участия в ответе растений на стресс (P.O. Owuor с соавт., 2010). За последние 5 лет получено много новых знаний о генах биосинтеза катехинов, L-теанина и кофеина, однако в мировой литературе отсутствуют обзоры, которые обобщают эти данные и связывают их с новыми данными по регуляции стрессовых ответов у чая. Цель настоящего обзора — анализ и обобщение современных сведений о генетических механизмах биосинтеза катехинов, L-теанина, кофеина в тканях чайного растения, а также их связь с генами-регуляторами абиотических стрессовых ответов. Биосинтез катехинов осуществляется по фенилпропаноидному и флавоноидному путям (A. Laura с соавт., 2019; S. Alseekh с соавт., 2020) при участии генов халконсинтазы (CHS), антоцианидинсинтетазы (ANS), антоцианидинредуктазы (ANR) и лейкоантоцианидинредуктазы (LAR) (J. Bogs с соавт., 2005). В накоплении катехинов в чайном растении участвуют факторы регуляции транскрипции семейства MYB, которые регулируют экспрессию генов PAL, F3′H и FLS (C.-F. Li с соавт., 2015). Образование кофеина происходит в основном в листьях чая при модификации пурина (H. Ashihara, 2015) с участием генов IMPDH (inosine monophosphate dehydrogenase), SAMS (S-adenosylmethioninesynthetase), MXMT (7-methylxanthine methyltransferase) и TCS (tea caffeine synthase). Уже известны 132 транскрипционных фактора, относящихся к 30 семействам (в их числе кодируемые генами семейств bZIP, bHLH и MYB), которыесвязаны с экспрессией генов биосинтеза кофеина (C.-F. Li с соавт., 2015). У C. sinensis образование L-теанина из глутамата с участием пирувата контролируется каскадом генов, основные из которых GS (glutamine synthetase), GOGAT (glutamate synthase), GDH (glutamate dehydrogenase), ALT (alanine transaminase), ADC (arginine decarboxylase) и TS (theanine synthetase) (C.Y. Shi с соавт., 2011; Y. Li с соавт., 2019). В регуляции этих процессов задействованы гены более 90 транскрипционных факторов — членов семейств AP2-EREBP, bHLH, C2H2 и WRKY, bZIP, C3H, MADS и REM (C.-F. Li с соавт., 2015). Обсуждается влияние стрессовых условий (засуха, холод, засоление, дефицита биогенных элементов) на образование и накопление биологически активных веществ. Недостаточно изученными остаются взаимосвязи между экспрессией генов метаболизма изучаемых соединений и транскрипционных факторов, а также изменения регуляторных сетей биосинтеза ценных метаболитов растений чая при различных экологических стрессах.
Ключевые слова: Camellia sinensis (L.) Kuntze, вторичные метаболиты, алкалоиды, аминокислоты, катехины, L-теанин, кофеин, гены метаболизма, экспрессия генов, транскрипционные факторы, засуха, пониженные температуры, засоление, питательные элементы.
Из молодых побегов растений чая (Camellia sinensis L.) получают напиток, который распространен во всем мире и высоко ценится благодаря широкому спектру полезных свойств, обусловленных сложным комплексом веществ (фенольные соединения, алкалоиды, эфирные масла, незаменимые аминокислоты, углеводы, минеральные соли, витамины, пектиновые вещества, пигменты, ферменты), присутствующих в листьях чайного растения (1). Многие из этих компонентов (около 700) относятся к биологически активным (1). Содержавшиеся в чае вещества влияют на сердечную деятельность и функцию нервной системы человека (2), повышают работоспособность мышечных тканей (3), вызывают состояние бодрости и стимулируют умственную деятельность (4, 5), укрепляют стенки кровеносных сосудов и капилляров (6), проявляют противолучевое, бактериостатическое и бактерицидное действие (7-10), активируют иммунную систему и способствуют профилактике некоторых видов рака (9).
Катехинам, которые представляют собой фенольные соединения, и алкалоиду кофеину (эти вещества относятся ко вторичным метаболитам), а также уникальной аминокислоте L-теанину (обнаружена только в растениях чая и не синтезируется в организме человека) отводится ключевая роль в формировании вкусовых качеств, пищевой и лекарственной ценности чая (2, 8, 10). Эти растительные метаболиты используются при производстве фармацевтических препаратов, пищевых добавок, ароматизаторов и других продуктов (11, 12). Содержание перечисленных веществ в тканях растений чая (и, соответственно, в получаемых продуктах) определяется генотипом (13, 14), районом произрастания (15-18), сезоном сбора урожая (19-22), элементным составом листьев (23-26), возрастом чайного листа (27), сроками и способами его переработки и хранения (28-30). Кроме того, на накопление биологически активных веществ существенно влияет количество и состав питательных элементов, поступающих в почву с удобрениями (31-35), что создает возможности для управления этим процессом.
Вторичные метаболиты проявляют свойства адаптогенов, смягчая последствия стресса, который испытывают растения при воздействии высоких и низких температур (36-39), ультрафиолетового облучения (40, 41), осмотического шока in vitro (42-44), инфекции патогенов (45, 46), минеральной недостаточности (47, 32), уровня освещения и генетических факторов, других факторов (48-51).
Основные пути биосинтеза катехинов, L-теанина и кофеина чайного растения детализированы в последние десятилетия, но механизмы регуляции происходящих биохимических процессов еще недостаточно изучены (41, 52-55). Так, ответственные за это генные сети у C. sinensis выявлены сравнительно недавно (56, 57). Благодаря транскриптомным исследованиям выявлены метаболические пути и ключевые гены, участвующие в биосинтезе, транспорте и метаболизме катехинов, кофеина и L-теанина (58-61), о которых подробнее обсуждаются ниже.
За последние 5 лет получено много новых знаний о генах биосинтеза катехинов, L-теанина и кофеина, однако в мировой литературе отсутствуют новые обзоры, которые обобщают эти сведения и связывают их с новыми данные по регуляции стрессовых ответов у чая.
Цель настоящего обзора — анализ и обобщение современных данных о генетических механизмах биосинтеза катехинов, L-теанина и кофеина в тканях чайного растения, а также связи их с генами-регуляторами абиотических стрессовых ответов.
Биосинтез основных вторичных метаболитов в растениях чая. Катехины. Эти фенольные соединений относятся к одному из наиболее распространенных классов растительных вторичных метаболитов. Катехины составляют 12-24 % сухой массы чайного листа (58) и на 70-75 % определяют крепость и терпкость получаемого напитка (62, 63). Катехины чайного растения представлены четырьмя простыми формами — (+)-катехином (C), (-)-эпикатехином (EC), (+)-галлокатехином (GC), (-)-эпигаллокатехином (EGC), а также их галлоильными производными — (-)-катехин-3-галлатом (CG), (-)-эпикатехин-3-галлатом (ECG), (-)-галлокатехин-3-галлатом (GCG), (-)-эпигалокатехин-3-галлатом (EGCG). Свойства катехинов в основном обусловлены числом и положением гидроксильной группы, обеспечивающей связывание и нейтрализацию свободных радикалов (64, 65). In vitro установлено, что катехины чая служат донорами электронов и эффективными гасителями активных форм кислорода, включая супероксидный анион, пероксильные радикалы и синглетный кислород (66).
Биосинтез катехинов, который в настоящее время хорошо изучен, осуществляется по фенилпропаноидному и флавоноидному путям (41, 67, 68). Флавоноидный путь биосинтеза начинается с образования халкона с участием халконсинтазы, кодируемой геном CHS, экспрессия которого коррелировала ссодержанием флавоноидов в растениях (69-71). У нескольких видов растений в геноме выявлено более одной копии CHS (72, 73). У C. sinensis найдены три копии гена CHS (CHS1, CHS2 и CHS3), экспрессия которых также коррелировала с накоплением катехинов в листьях и побегах (74), а профили экспрессии, по предположению G.E. Mamati с соавт. (75), зависели от возраста листа. Однако до сих пор неизвестно, какой из трех генов CHS играет ключевую роль в биосинтезе катехинов у C. sinensis.
Кроме CHS, в биосинтезе катехинов участвуют гены флавоноид 3'-монооксигеназы (F3'H), флаванолсинтазы (FLS), гены антоцианидинсинте-тазы (ANS), антоцианидинредуктазы (ANR) и лейкоантоцианидинредуктазы (LAR), фенилаланин-аммоний-лиазы (PAL). Продукт гена PAL катализирует первый шаг метаболизма в фенилпропаноидном пути. F3'H и FLS кодируют ферменты синтеза флавонолов в флавоноидном пути. ANS катализирует превращение лейкоцианидинов в антоцианидины (76). Ген ANR вовлечен в биосинтез мономеров флаван-3-олов, а именно в превращение антоцианидина в эпикатехин (77). Продукт гена LAR катализирует превращение лейкоцианидина, лейкодельфинина или лейкопеларгонидина в соответствующие 2,3-транс-флаван-3-олы (78). Все эти гены играют важную роль в определении состава катехинов в листьях чая (41, 79-81). L. Zhang с соавт. (82) обнаружили положительную корреляцию между интенсивностью экспрессии генов CHS1, CHS3, ANR1, ANR2, LAR и общим содержанием катехинов. При этом уровень экспрессии гена ANS имел положительную связь с накоплением простых катехинов, а генов ANR1,ANR2 и LAR — с содержанием (-)-эпигаллокатехин-галлата и (-)-эпикатехин-галлата (82). Высказывалось предположение, что у растений чая наиболее важны для биосинтеза катехинов гены F3'H и ANS, экспрессия которых существенно повышалась в осенний период одновременно с накоплением катехинов (83). Однако до сих пор крайне мало данных о взаимосвязи экспрессии этих генов с составом и количественным соотношением катехинов в чайном растении. Предполагается, что экспрессию генов, вовлеченных в биосинтез соединений фенольной природы, регулируют факторы транскрипции MYB, bHLH, WRKY и другие, транскрипционные факторы, связанные с АБК-опосредованным ответом растений на стрессы (84-87). Так, гены семейства MYB вовлечены в регуляцию экспрессии генов биосинтеза флавоноидов (PAL, F3'H и FLS) у чайного растения, что подтверждает значение факторов транскрипции MYB в контроле накопления флавоноидов. В частности, у растений чая гены CsMYB8, CsMYB99, MYB23 (семейство MYB), bHLH96 (семейство bHLH) и NAC008 (семейство NAC) участвуют в регуляции биосинтеза флавоноидов, включая катехины, антоцианы и флавонолы (88, 89). Повышенная экспрессия этих транскрипционных факторов положительно коррелировала с накоплением катехинов (88, 89). Сообщалось о 206 транскрипционных факторах из 33 семейств, ассоциированных с изменением функциональной активности 36 генов биосинтеза флавоноидов (57). При этом признается, что в настоящее время механизмы регуляции биосинтеза и транспорта флавоноидов и антоцианов все еще остаются недостаточно изученными (89).
Кофеин (1,3,7-триметилксантин). Этот биоактивный компонент, синтезируемый C. sinensis, представляет собой пуриновый алкалоид, широко используемый в качестве стимулятора и компонента лекарственных препаратов (90). Как по накоплению в растении, так и по фармакологическому действию это доминирующий среди всех алкалоидов чая (1). Кофеин придает терпкость чайному настою, а также существенно влияет на его крепость (29). Кроме того, содержание кофеина характеризует активность физиологических процессов в растениях, в частности окислительно-восстановительных и ферментативных реакций, белкового обмена (90). Содержание кофеина в среднем составляет 3 % сухой массы чайного листа и в зависимости от ряда факторов (условий среды, генетических и географических факторов) колеблется в пределах от 1,5 до 4,5 % (30, 90).
Кофеин синтезируется преимущественно в молодых листьях растений из пуриновых нуклеотидов в реакциях аденинового метаболизма (91). Основной путь биосинтеза кофеина включает ряд последовательных превращений ксантозин (XR) → 7-метилксантозин (7-mXR) → 7-метилксантин (7-mX) → теобромин (Tb) → кофеин (Cf) с участием фермента N-метилтрансферазы, кодируемого геном NMT, который также называется геном TCS (кофеинсинтетаза) (92-94). N-метилтрансфераза проявляет трансметилирующую активность на двух стадиях, катализируя превращение 7-mX в Tb и Tb в Cf (95). В биосинтезе кофеина также участвуют гены IMPDH (инозин-5-монофосфатдегидрогеназа), SAMS (S-аденозил-L-метионинсинтаза), MXMT(7-метилксантинметилтрансфераза). Изучение активности аллельных вариантов гена TCS1 в популяциях чайных растений подтвердило, что фермент кофеинсинтетазаопределяет содержание кофеина в растительных тканях. По мнению P. Li с соавт. (88), к семейству MYB относятся основные активаторы транскрипции гена TCS1, при этом CsMYB184, CsMYB85 и CsMYB86 играют ключевую роль в регуляции биосинтеза кофеина (96). Факторы транскрипции семейств AP2/ERF, WRKY, bHLH, MYB, bZIP, TFIIIA и AT-hook регулируют экспрессию структурных генов родственных синтетаз, участвующих в биосинтезе алкалоидов (97). Например, продукты генов семейств GATA и bHLH связываются с сайтами инициации транскрипции 12 важнейших генов биосинтеза кофеина семейства AMPD(кодируют ферменты аденозин 5-монофосфатдеаминазы), влияя на их экспрессию (98). Недавно выявлены сайты узнавания для продукта гена MYB184, который проявлял высокую промоторную активность, усиливая экспрессию гена TCS1 в 4,7 раза (96). В транскриптомных исследованиях тканей чайного растения на разных стадиях развития были выявлены регуляторные сети, включающие 132 фактора транскрипции из 30 семейств, связанные с экспрессией 24 генов биосинтеза кофеина (57). Большинство из этих транскрипционных факторов принадлежат к семействам bZIP, bHLH и MYB.
Анализ сортов чая с разным содержанием кофеина показал, что транскрипционные факторы семейства NAC связаны с биосинтезом пуриновых алкалоидов (99). Один из генов этого семейства — CsNAC7, по данным W. Ma с соавт. (100),положительно регулирует активность гена основного фермента биосинтеза кофеина N-метилтрансферазы табака yhNMT1. Анализ функциональной активности CsNAC7 показал, что его временная сверхэкспрессия может значительно усиливать экспрессию yhNMT1 в листьях табака (96). Однако связь функциональной активности генов метаболизма кофеина с транскрипционными факторами требует дальнейшего изучения. В частности, недостаточно данных об изменении регуляторных сетей биосинтеза этого алкалоида при азотном дефиците, влияющем на продуктивность растений чая.
L-Теанин (5-N-этилглутамин). Эта аминокислота составляет до 50 % от общего количества аминокислот в черном чае и 1-2 % от сухой массы зеленого чая (101-103).Она придает сладкий и пикантный вкус чайному напитку (9, 104). Образование L-теанина происходит в корнях, откуда он транспортируется по флоэме к растущим побегам и накапливается в молодых листьях (90, 105). У C. sinensis образование L-теанина из глутамата с участием пирувата контролируется каскадом генов, основные из которых GS (глутаминсинтетаза), GOGAT (глутаматсинтетаза), GDH (глутаматдегидрогеназа), ALT (аланинтрансаминаза), ADC (аргининдекарбоксилаза) и TS (треанинсинтетаза) (56). L-Теанин может подвергаться гидролизу, превращаясь в этиламин и далее вновь использоваться как предшественник в биосинтезе катехинов, что отмечали при длительном воздействии солнечного света (54). Превращение глутамина и этиламина в L-теанин у C. sinensis осуществляет фермент теанинсинтетаза (TS), который имеет очень высокую степень гомологии с глутаматсинтетазой (GS) (54). Глутамин — прекурсор L-теанина синтезруется с участием глутамин-2-оксоглутаратаминотрансферазы и глутаматдегидрогеназы (105). Другой предшественник L-теанина — этиламин образуется при декарбоксилировании аланина (Ala), которое катализирует фермент аланиндекарбоксилаза AlaDC (106). Аланин и ацетальдегид могут быть прекурсорами этиламина в тканях растений (107, 108), при этом аланин предшествует ацетальдегиду в путях биосинтеза (109). Хотя ключевые гены биосинтеза L-теанина известны, их транскрипционная регуляция остается малоизученной (110). Сравнительно недавно были найдены более 90 транскрипционных факторов из семейств AP2-EREBP, bHLH, C2H2 и WRKY, bZIP, C3H, MADS и REM, которые участвуют врегуляции биосинтеза L-теанина (57). По данным P. Li с соавт. (88), гены факторов транскрипции CsMYB9 и CsMYB49 участвуют в контроле биосинтеза L-теанина, а экспрессия гена фактора транскрипции CsMYB73 отрицательно коррелировала с накоплением L-теанина в процессе созревания листьев. В листьях табака продукт гена CsMYB73 связывается с промоторными регионами генов CsGS1 и CsGS2 и подавляет их транскрипцию (110). Кроме того, транскрипционный фактор CsWRKY40 активировал ключевой ген гидролиза L-теанина CsPDX2.1 (пиридоксаль-5'-фосфатсинтаза). При увядании и потере влаги в листьях накапливалась абсцизовая кислота, а содержание L-теанина снижалось на фоне активации экспрессии CsWRKY40 и CsPDX2.1 (111).
Таким образом, в регуляции биосинтеза катехинов, кофеина и L-теанина, определяющих качество получаемой растительной продукции C. sinensis, в общей сложности участвуют 339 транскрипционных факторов, относящихся к 35 семействам (57). Важно отметить наличие 67 общих факторов транскрипции в регуляторных сетях биосинтеза катехинов и кофеина (57). Это указывает на положительную корреляцию между их накоплением (57), что представляет интерес как с точки зрения фундаментальных механизмов вторичного метаболизма растений, так и для решения практических задач селекции и оптимизации технологий возделывания культуры. Напротив, лишь два транскрипционных фактора оказались общими в регуляции экспрессии генов биосинтеза катехинов и L-теанина, что подтверждает обратную связь между их продукцией в растении. Тот факт, что на активность генов, ответственных за биосинтез катехинов, кофеина и L-теанина, влияют транскрипционные факторы из разных семейств, свидетельствует о сложной системе транскрипционного контроля при образовании рассматриваемых биологически активных вторичных метаболитов.
Биосинтез вторичных метаболитов при абиотическом стрессе. Стрессовые условия среды существенно изменяют содержание катехинов, кофеина и L-теанина в растениях чая (112, 113). Транскриптомные исследования выявили ключевые транскрипционные факторы, вовлеченные в ответ на абиотический стресс у чайного растения (44, 114-116). Установлено, что многие семейства факторов транскрипции (CBF,bHLH,WRKY) участвуют в ответах на разные абиотические стрессы (холод, засуху, засоление), то есть неспецифичны (117-121).
Летняя притенка чайных растений снижала содержание катехинов и подавляла экспрессию гена ANS (122), тогда как функциональная активность генов халконсинтазы(CHS), флавоноид-3-гидроксилазы (F3’H) и дигидрофлавонол-4-редуктазы (DFR,) не изменялась (52). Повышение уровня освещенности при культивировании каллусов чая in vitro способствовало накоплению катехинов (123). Кратковременное (30 мин) воздействие ультрафиолетового облучения (UV-B) на однолетние вегетативно размноженные саженцы C. sinensis сортов Yulan и Fudingdabai в горшечной культуре повышало, а продолжительное (360 мин) воздействие, наоборот, снижало содержание катехинов (124).
Накопление катехинов также зависело от водного статуса растений и применения удобрений (125-127), доступа углерода и гормонального баланса (78). Так, при продолжительном воздействии засухи отмечено кратковременное снижение, а затем повышение экспрессии генов CHS, DFR, LAR, ANS и ANR, что коррелировало с накоплением галлата эпикатехина, галлата эпигаллокатехина и галлата галлокатехина (128, 129). Сообщалось о снижении содержания полифенолов в листьях чая при засухе (125, 130). Однако в условиях кратковременной засухи (2 сут) уровень экспрессии генов FLS и FNS повышался, что сопровождалось увеличением накопления соединений из группы флавоноидов (128). У чайного растения активность основных выявленных генов биосинтеза кофеина подавлялись в ответ на засуху (128), а содержание кофеина в 3-листной флеши снижалось (в среднем на 1 %) в сравнении с показателем при нормальной влагообеспеченности растений (23, 30). Сообщалось (128), что содержание L-теанина в листьях C. sinensis и уровень экспрессии генов GOGAT, GDH, ADC и TS при засухе снижается, тогда как экспрессия гена ThYD (L-треанингидролаза), который кодирует ключевой фермент деградации L-теанина, повышалась.
В условиях азотного голодания у двух сортов чая был выявлен ген AlaDC (аннотированный как ген сериндекарбоксилазы), который может выполнять специфическую роль в накоплении L-теанина (128, 131). Известно, что азот относится к наиболее важным элементам для биосинтеза L-теанина, кофеина и катехинов (28, 132-134). При дефиците азота растения чая накапливали различные флавоноиды, тогда как на фоне обеспеченности этим элементом значительно усиливался синтез аминокислот, в том числе L-теанина (62, 135). Общее содержание катехинов также существенно зависело от количества и соотношения доступных форм азота, фосфора и калия в почве (136). При этом накопление простых катехинов (эпигаллокатехина, эпикатехина, галлокатехина и катехина) обратно коррелировало с количеством внесенных в почву N, P, K, а их галлированных форм — прямо коррелировало с величиной доз P и K (136). Также сообщалось, что повышенные дозы фосфора и калия, которые приводили к накоплению катехинов и углеводов в побегах чая, снижали относительное содержание свободных аминокислот, в частности L-теанина и глутаминовой кислоты (137).
Транскрипционные факторы и гены метаболизма, вовлеченные в биосинтез катехинов, кофеина и L-теанина в чайном растении, представлены на рисунке.
Итак, за последние годы выявлены основные гены метаболизма, вовлеченные в биосинтез катехинов, кофеина и L-теанина у чайного растения и их роль в каскаде биохимических реакций, идентифицированы некоторые транскрипционные факторы, участвующие в регуляции экспрессии этих генов. Предполагается, что выявленные транскрипционные факторы могут быть связаны с регуляторами стрессовых ответов, в частности через путь ответа, опосредованный абсцизовой кислотой. Однако все еще недостаточно знаний о функциональной роли регуляторов биосинтеза катехинов, кофеина и L-теанина во взаимосвязи с ключевыми транскрипционными факторами стрессовых ответов. Это направление представляется перспективным для дальнейших исследований.
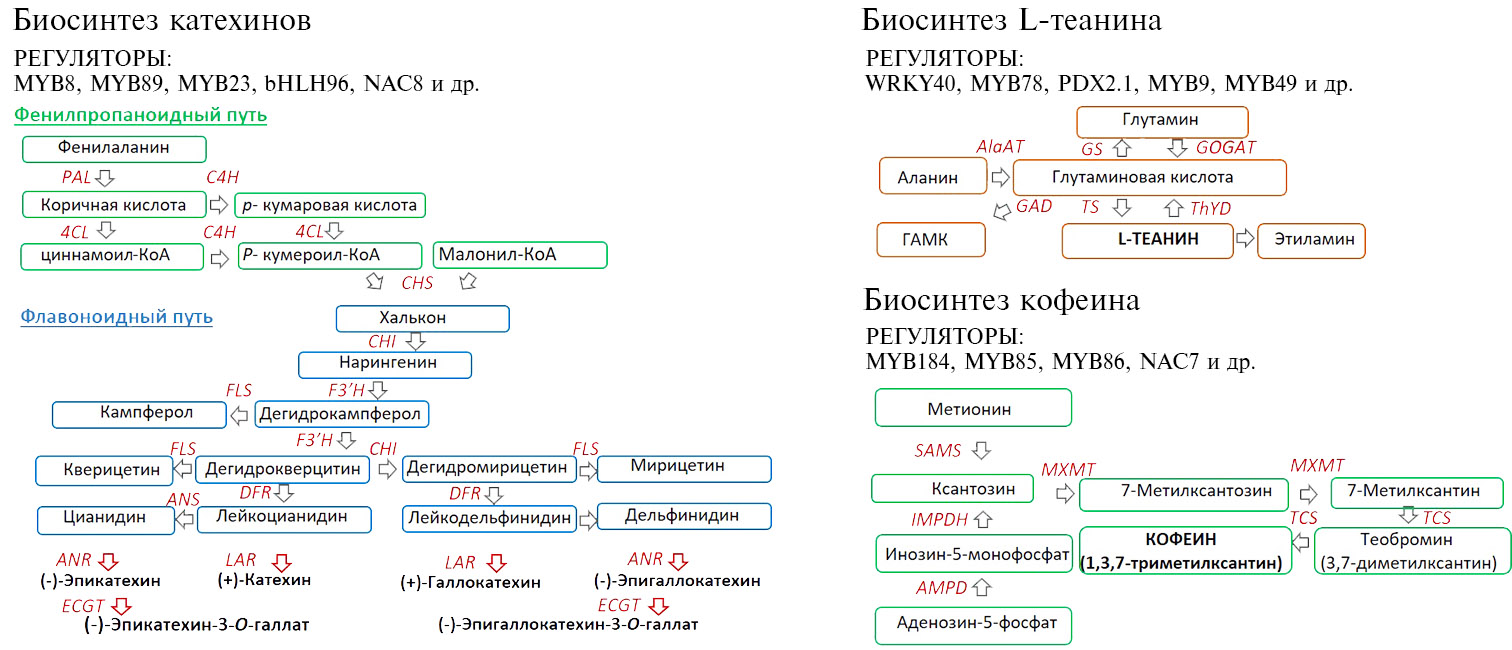
Транскрипционные факторы и гены биосинтез катехинов, кофеина и L-теанина в чайном растении Camellia sinensis (L.) Kuntze, представленные в настоящем обзоре.
ЛИТЕРАТУРА
- Zhao J., Li P., Xia T., Wan X. Exploring plant metabolic genomics: chemical diversity, metabolic complexity in the biosynthesis and transport of specialized metabolites with the tea plant as a model. Crit. Rev. Biotechnol., 2020, 40(5): 667-688 (doi: 10.1080/07388551.2020.1752617).
- Sharangi A.B. Medicinal and therapeutic potentialities of tea (Camellia sinensis L.) — a review. Food Research International, 2009, 42 (5-6): 529-535 (doi: 10.1016/j.foodres.2009.01.007).
- Frei B., Higdon J.V. Antioxidant activity of tea polyphenols in vivo: evidence from animal studies. Journal of Nutrition, 2003, 133(10): 3275-3284 (doi: 10.1093/jn/133.10.3275S).
- Boschmann M., Thielecke, F. The effects of epigallocatechin-3-gallate on thermogenesis and fat oxidation in obese men: a pilot study. Journal of the American College of Nutrition, 2007, 26(4): 389-395 (doi: 10.1080/07315724.2007.10719627).
- Khan N., Mukhtar H. Tea polyphenols for health promotion. Life Sciences, 2007, 81(7): 519-533 (doi: 10.1016/j.lfs.2007.06.011).
- Velayutham P., Babu A., Liu D. Green tea catechins and cardiovascular health: An update. Current medicinal chemistry,2008, 15(18): 1840-1850 (doi: 10.2174/092986708785132979).
- Lorenzo J.M., Munekata P.E.S. Phenolic compounds of green tea: health benefits and technological application in food. Asian Pacific Journal of Tropical Biomedicine, 2016, 6(8): 709-719 (doi: 10.1016/j.apjtb.2016.06.010).
- Yan Z., Zhong Y., Duan Y., Chen Q., Li F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Animal Nutrition, 2020, 6(2): 115-123 (doi: 10.1016/j.aninu.2020.01.001).
- Vuong Q.V., Bowyer M.C., Roach P.D. L-Theanine: properties, synthesis and isolation from tea. Journal of the Science of Food and Agriculture, 2011, 91(11): 1931-1939 (doi: 10.1002/jsfa.4373).
- Khalesi S., Sun J., Buys N., Jamshidi A., Nikbakht-Nasrabadi Е., Khosravi-boroujeni H. Green tea catechins and blood pressure: a systematic review and meta-analysis of randomized controlled trials. European Journal of Nutrition, 2014, 53(6): 1299-1311 (doi: 10.1007/s00394-014-0720-1).
- Karuppusamy S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. Journal of Medicinal Plants Research, 2009, 3(13): 1222-1239 (doi: 10.5897/JMPR.9000026).
- Gaurav N., Juyal P., Tyagi M., Chauhan N., Kumar A. A review on in vitro propagation of medicinal plants. Journal of Pharmacognosy and Phytochemistry, 2018, 7(6): 2228-2231.
- Zhang X.B. Differences of polyphenols content in Anxi TieGuanYin tea among different seasons and relationship between polyphenols and tea quality. Agricultural Science & Technology, 2014, 15(7): 1191-1195.
- Fang R., Redfern S.P., Kirkup D., Porter E.A., Kite G.C., Terry L.A., Berry M.J., Simmonds M.S.J. Variation of theanine, phenolic, and methylxanthine compounds in 21 cultivars of Camellia sinensis harvested in different seasons. Food Chemistry, 2017, 220: 517-526 (doi: 10.1016/j.foodchem.2016.09.047).
- Owuor P.O., Kamau D.M., Jondiko E.O. The influence of geographical area of production and nitrogenous fertilizer on yields and quality parameters of clonal tea. Journal of Food, Agriculture and Environment, 2010, 8: 682-690 (doi: 10.20425/IJTS14116).
- Малюкова Л.С., Цюпко Т.Г., Притула З.В., Воронова О.Б., Гущаева К.С., Великий А.В. Состав и содержание флавоноидов в готовом чае, выращенном в условиях Черноморского побережья Краснодарского края. В сб.: Фенольные соединения: функциональная роль в растениях. М., 2018: 272-277.
- Belous O.G, Platonova N.B. Biologically active substances of Сamellia sinensis in a humid subtropical climate of Russia. Slovak Journal of Food Sciences, 2021, 15: 360-368 (doi: 10.5219/1440).
- Гущаева К.С., Цюпко Т.Г., Воронова О.Б., Малюкова Л.С. Определение кофеина, катехинов и галловой кислоты в черном чае различного происхождения. Заводская лаборатория. Диагностика материалов, 2021, 87(9): 12-19 (doi: 10.26896/1028-6861-2021-87-9-12-19).
- Ercisli S., Orhan E., Ozdemir O., Sengul M., Gungor N. Seasonal variation of total phenolic, antioxidant activity, plant nutritional elements and fatty acids in tea leaves (Camellia sinensis var. sinensis clone Derepazari 7) grown in Turkey. Pharmaceutical Biology, 2008, 46(10-11): 683-687 (doi: 10.1080/13880200802215818).
- Lee L.-S., Kim S.-H., Kim Y.-B., Kim Y.-C. Quantitative analysis of major constituents in green tea with different plucking periods and their antioxidant activity. Molecules, 2014, 19(7): 9173-9186 (doi: 10.3390/molecules19079173).
- Gong A.D., Lian S.B., Wu N.N., Zhou Y.J., Zhao S.Q., Zhang L.M., Cheng L., Yuan H.Y. Integrated transcriptomics and metabolomics analysis of catechins, caffeine and theanine biosynthesis in tea plant (Camellia sinensis) over the course of seasons. BMC Plant Biology, 2020, 20(1): 294 (doi: 10.1186/s12870-020-02443-y).
- Jamir T. Seasonal variations in antioxidant capacities and phenolic contents of tea leaf extracts. Asian Journal of Pharmaceutical and Clinical Research, 2020, 13(4): 108-112 (doi: 10.22159/ajpcr.2020.v13i4.36866).
- Belous O.G. Influence of microelements on biochemical parameters of tea. Potravinarstvo, 2013, 7(S): 149-152.
- Diniz P.H.G.D., Pistonesi M. F., Alvarez M.B., Band B.S.F., Araujo M.C.U. Simplified tea classification based on a reduced chemical composition profile via successive projections algorithm linear discriminant analysis (SPA-LDA). Journal of Food Composition and Analysis, 2015, 39: 103-110 (doi: 10.1016/J.JFCA.2014.11.012).
- Рындин А.В., Малюкова Л.С., Цюпко Т.Г., Воронова О.Б., Гущаева К.С. Особенности элементного состава Краснодарского чая сорта Колхида. Новые технологии, 2018, 4: 224-229.
- Притула З.В., Малюкова Л.С., Бехтерев В.Н. Диапазоны варьирования и корреляционная связь между основными биохимическими и минеральными компонентами 3-листной флеши чая сорта Колхида во влажно-субтропической зоне России. Плодоводство и ягодоводство России, 2019, 59: 92-98 (doi: 10.31676/2073-4948-2019-59-92-98).
- Зубова М.Ю., Николаева Т.Н., Нечаева Т.Л., Малюкова Л.С., Загоскина Н.В. О содержании пигментов, фенольных соединений и антирадикальной активности молодых побегов чая (Camellia sinensis l.). Химия растительного сырья, 2019, 4: 249-257 (doi: 10.14258/jcprm.2020016065).
- Sabhapondit S., Karak T., Bhuyan L.P., Goswami B.C., Hazarika M. Diversity of catechin in northeast Indian tea cultivars. The Scientific World Journal,2012, 1-8 (doi: 10.1100/2012/485193).
- Sari F., Velioglu Y.S. Changes in theanine and caffeine contents of black tea with different rolling methods and processing stages. European Food Research and Technology, 2013, 237(2): 229-236 (doi: 10.1007/s00217-013-1984-z).
- Притула З.В., Бехтерев В.Н., Малюкова Л.С. Влияние мезоудобрений (Ca, Mg) на содержание кофеина в чайном растении в условиях влажных субтропиков России. Субтропическое и декоративное садоводство, 2015,54: 185-192.
- Runa J., Haerdter R., Gerendás J. Impact of nitrogen supply on carbon/nitrogen allocation: a case study on amino acids and catechins in green tea [Camellia sinensis (L.) O. Kuntze] plants. Plant Biology, 2010: 12(5): 724-734 (doi: 10.1111/j.1438-8677.2009.00288.x).
- Hrishikesh U., Biman K, D., Lingaraj S., Sanjib K. P. Comparative effect of Ca, K, Mn and B on post-drought stress recovery in tea [Camellia sinensis (L.) O. Kuntze]. American Journal of Plant Sciences, 2012, 3(4): 443-460 (doi: 10.4236/ajps.2012.34054).
- Ruan J., Ma L., Shi Y. Potassium management in tea plantations: its uptake by field plants, status in soils, and efficacy on yields and quality of teas in China. Journal of Plant Nutrition and Soil Science, 2013, 176(3): 450-459 (doi: 10.1002/jpln.201200175).
- Kwach B.O., Owuor P., Kamau D., Msomba S., Uwimana M.A. Variations in the precursors of plain black tea quality parameters due to location of production and nitrogen fertilizer rates in eastern African clonal tea leaves. Experimental Agriculture, 2016: 52(2): 266-278 (doi: 10.1017/S0014479715000083).
- Malyukova L.S., Pritula Z.V., Kozlova N.V., Veliky A.V., Rogozhina E.V., Kerimzade V.V., Samarina L.S. Effects of calcium-containing natural fertilizer on Camellia sinensis (L.) Kuntze. Bangladesh Journal of Botany, 2021, 50(1): 179-187 (doi: 10.3329/bjb.v50i1.52686).
- Hernaendez I., Alegre L., Munne-Bosch S. Enhanced oxidation of flavan-3-ols and proanthocyanidin accumulation in water-stressed tea plants. Phytochemistry, 2006, 67(11): 1120-1126 (doi: 10.1016/j.phytochem.2006.04.002).
- Xu Z., Zhou G., Shimizu H. Plant responses to drought and rewatering. Plant Signaling & Behavior, 2010, 5(6): 649-654 (doi: 10.4161/psb.5.6.11398).
- Maritim T.K., Kamunya S.M., Mireji P., Mwendia C.M., Muoki R.C., Cheruiyot E.K., Wachira F.N. Physiological and biochemical response of tea (Camellia sinensis (L.) O. Kuntze) to water-deficit stress. The Journal of Horticultural Science and Biotechnology, 2015, 90(4): 395-400 (doi: 10.1080/14620316.2015.11513200).
- Samarina L.S., Bobrovskikh A.V., Doroshkov A.V., Malyukova L.S., Matskiv A.O., Rakhmangulov R.S., Koninskaya N.G., Malyarovskaya V.I., Tong W., Xia E, Manakhova K.M., Ryndin A.V., Orlov Y.L. Comparative expression analysis of stress-inducible candidate genes in response to cold and drought of tea plant (Camellia sinensis (L.) Kuntze). Frontiers in Genetics, 2020, 11: 1613 (doi: 10.3389/fgene.2020.611283).
- Agati G., Tattini M. Multiple functional roles of flavonoids in photo protection. New Phytolgist, 2010, 186(4): 786-793 (doi: 10.1111/j.1469-8137.2010.03269.x).
- Wang Y.S., Gao L.P., Wang Z.R., Liu Y.J., Sun M., Yang D., Wei C., Shan Y., Xia T. Light-induced expression of genes involved in phenylpropanoid biosynthetic pathways in callus of tea (Camellia sinensis (L.) O. Kuntze). Scientia Horticulturae, 2012, 133: 72-83 (doi: 10.1016/J.SCIENTA.2011.10.017).
- Малюкова Л.С., Самарина Л.С., Конинская Н.Г., Притула З.В., Гвасалия М.В., Цюпко Т.Г., Воронова О.Б. Образование и накопление компонентов катехиновой группы в микропобегах чая при осмотическом стрессе в условиях in vitro. АгроЭкоИнфо, 2019, 4: 30.
- Малюкова Л.С., Нечаева Т.Л., Зубова М.Ю., Гвасалия М.В., Конинская Н.Г., Загоскина Н.В. Физиолого-биохимические характеристики микропобегов чая (Camellia sinensis L.) в условиях in vitro: норма, осмотический стресс, кальций. Сельскохозяйственная биология, 2020, 55(5): 970-980 (doi: 10.15389/agrobiology.2020.5.970rus).
- Samarina L., Matskiv A., Simonyan T., Koninskaya N., Malyarovskaya V., Gvasaliya M., Malyukova L., Mytdyeva A., Martinez-Montero M., Choudhary R., Ryndin A. Biochemical and genetic responses of tea (Camellia sinensis (L.) Kuntze) microplants under the mannitol-induced osmotic stress in vitro. Plants, 2020, 9(12): 1795 (doi: 10.3390/plants9121795).
- Peng P., Xie Q., Li P., Hou Y., Hu X., Lin, Q. Studies on the allelopathy components of tea. Southwest China Journal of Agricultural Sciences, 2009, 22(1): 67-70.
- Zeng L., Zhou X., Liao Y., Yang Z. Roles of specialized metabolites in biological function and environmental adaptability of tea plant (Camellia sinensis) as a metabolite studying model. Journal of Advanced Research, 2021, 34: 159-171 (doi: 10.1016/j.jare.2020.11.004).
- Lillo C., Lea U., Ruoff P. Nutrient depletion as a key factor for manipulating gene expression and product formation in different branches of the flavonoid pathway. Plant Cell Environ., 2008, 31(5): 587-601 (doi: 10.1111/j.1365-3040.2007.01748.x).
- Akola R., Ravishankar G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signaling &Behavior , 2011, 6(11): 1720-1731 (doi: 10.4161/psb.6.11.17613).
- Xiong L.G., Lia J., Lia Y.H., Yuan L., Liu S., Huang J., Liu Z. Dynamic changes in catechin levels and catechin biosynthesis-related gene expression in albino tea plants (Camellia sinensis L.). Plant Physiology and Biochemistry,2013, 71: 132-143 (doi: 10.1016/j.plaphy.2013.06.019).
- Hong G.J., Wang J., Zhang Y., Hochstetter D., Zhang S., Pan Y., Shi Y., Xu P., Wang Y. Biosynthesis of catechin components is differentially regulated in dark-treated tea (Camellia sinensis L.). Plant physiology and biochemistry, 2014, 78: 49-52 (doi: 10.1016/j.plaphy.2014.02.017).
- Lin S., Chen Z., Chen T., Deng W., Wan X., Zhang Z. Theanine metabolism and transport in tea plants (Camellia sinensis L.): advances and perspectives. Critical Reviews in Biotechnology, 2022, 1-15 (doi: 10.1080/07388551.2022.2036692).
- Eungwanichayapant P.D., Popluechai S. Accumulation of catechins in tea in relation to accumulation of mRNA from genes involved in catechin biosynthesis. Plant Physiology and Biochemistry,2009, 47(2): 94-97 (doi: 10.1016/j.plaphy.2008.11.002).
- Jia S., Wang Y., Hu J., Ding Z., Liang Q., Zhang Y., Wang H. Mineral and metabolic profiles in tea leaves and flowers during flower development. Plant Physiology and Biochemistry, 2016, 106: 316-326 (doi: 10.1016/j.plaphy.2016.06.013).
- Bai P., Wei K., Wang L., Zhang F., Ruan L., Li H., Wu L., Cheng H. Identification of a novel gene encoding the specialized alanine decarboxylase in tea (Camellia sinensis). Molecules, 2019, 24(3): 540 (doi: 10.3390/molecules24030540).
- Liao Y., Zhou X., Zeng L. How does tea (Camellia sinensis) produce specialized metabolites which determine its unique quality and function: a review. Critical Reviews in Food Science and Nutrition, 2022, 62(14): 3751-3767 (doi: 10.1080/10408398.2020.1868970).
- Shi C.Y., Yang H., Wei C.L., Yu O., Zhang Z.Z., Jiang C. J., Sun J., Li Y.Y., Chen Q., Xia T., Wan X.C. Deep sequencing of the Camellia sinensis transcriptome revealed candidate genes for major metabolic pathways of tea-specific compounds. BMC Genomics,2011, 12(1): 131 (doi: 10.1186/1471-2164-12-131).
- Li C.-F., Zhu Y., Yu Y., Zhao Q.-Y., Wang S.-J., Wang X.-C., Yao M.-Z., Luo D., Li X., Chen L., Yang Y.-J. Global transcriptome and gene regulation network for secondary metabolite biosynthesis of tea plant (Camellia sinensis). BMC Genomics, 2015, 16(1): 560 (doi: 10.1186/s12864-015-1773-0).
- Wei C., Yang H., Wang S., Zhao J., Liu C., Gao L., Xia E., Lu Y., Tai Y., She G., Sun J., Cao H., Tong W., Gao Q., Li Y., Deng W., Jiang X., Wang W., Chen Q., Zhang Sh., Li Y., Wu J., Wang P., Li P., Shi Ch., Zheng F., Jian J., Huang B., Shan D., Shi M., Fang C., Yue Y., Li F., Li D., Wei Sh., Han B., Jiang Ch., Yin Y., Xia T., Zhang Z., Bennetzen J.L., Zhao Sh., Wan X. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proceedings of the National Academy of Sciences, 2018, 115(18): E4151-E4158 (doi: 10.1073/pnas.1719622115).
- Li Y., Wang X., Ban Q., Zhu X., Jiang C., Wei C., Bennetzen J.L. Comparative transcriptomic analysis reveals gene expression associated with cold adaptation in the tea plant Camellia sinensis. BMC Genomics, 2019, 20(1): 624. (doi: 10.1186/s12864-019-5988-3).
- Xia E.H., Li F.D., Tong W., Li P.H., Wu Q., Zhao H.J., Ge R.H., Li R.P., Li Y.Y., Zhang Z.Z., Wei C.L., Wan X.C. Tea plant information archive: a comprehensive genomics and bioinformatics platform for tea plant. Plant Biotechnology Journal, 2019, 17(10): 1938-1953 (doi: 10.1111/pbi.13111).
- Xia E.H., Tong W., Wu Q., Wei S., Zhao J., Zhang Z. Z., Wei C.L., Wan X. C. Tea plant genomics: achievements, challenges and perspectives. Horticulture Research, 2020, 7: 7 (doi: 10.1038/s41438-019-0225-4).
- Huang H., Yao Q., Xia E., Gao L. Metabolomics and transcriptomics analyses reveal nitrogen influences on the accumulation of flavonoids and amino acids in young shoots of tea plant (Camellia sinensis L.) associated with tea flavor. Journal of agricultural and food chemistry, 2018, 66(37): 9828-9838 (doi: 10.1021/acs.jafc.8b01995).
- Zhen Y.S. Antitumor activity of tea products. In: Tea: bioactivity and therapeutic potential /Y.S. Zhen, Z.M. Chen, S.J. Cheng, V.L. Chen (eds.). CRC Press, London. New York,2002 (doi: 10.1201/b12659).
- Guo Q., Zhao B., Shen S., Hou J., Hu J., Xin W. ESR study on the structure-antioxidant activity relationship of tea catechins and their epimers. Biochimica et Biophysica Acta (BBA)-General Subjects, 1999, 1427(1): 13-23 (doi: 10.1016/S0304-4165(98)00168-8).
- Farkas O., Jakus J., Héberger K. Quantitative structure-antioxidant activity relationships of flavonoid compounds. Molecules, 2004, 9(12): 1079-1088 (doi: 10.3390/91201079).
- Nakagawa T., Yokozawa T. Direct scavenging of nitric oxide and superoxide by green tea. Food and chemical Toxicology, 2002, 40(12): 1745-1750 (doi: 10.1016/s0278-6915(02)00169-2).
- Laura A., Moreno-Escamilla J.O., Rodrigo-García J., Alvarez-Parrilla E. Phenolic compounds. In: Postharvest physiology and biochemistry of fruits and vegetables /E.M.Yahia (ed.). Woodhead Publishing, 2019 (doi: 10.1016/B978-0-12-813278-4.00012-9).
- Alseekh S., Perez de Souza L., Benina M., Fernie A. R. The style and substance of plant flavonoid decoration; towards defining both structure and function. Phytocnemistry,2020, 174: 112347 (doi: 10.1016/j.phytochem.2020.112347).
- Kamiishi Y., Otani M., Takagi H., Han D.S., Mori S., Tatsuzawa F., Okuhara H., Kobayashi H., Nakano M. Flower color alteration in the liliaceous ornamental Tricyrtis sp. by RNA interference mediated suppression of the chalcone synthase gene. Molecular breeding, 2012, 30(2): 671-680 (doi: 10.1007/s11032-011-9653-z).
- Morita Y., Saito R., Ban Y., Tanikawa N., Kuchitsu K., Ando T., Yoshikawa M., Habu Y., Ozeki Y., Nakayama M. Tandemly arranged chalcone synthase A genes contribute to the spatially regulated expression of siRNA and the natural bicolor floral phenotype in Petunia hybrida. The Plant Journal, 2012, 70(5): 739-749 (doi: 10.1111/j.1365-313X.2012.04908.x).
- Dare A.P., Tomes S., Jones M., McGhie T.K., Stevenson D.E., Johnson R.A., Greenwood D.R., Hellens R.P. Phenotypic changes associated with RNA interference silencing of chalcone synthase in apple (Malus × domestica). The Plant Journal, 2013, 74(3): 398-410 (doi: 10.1111/tpj.12140).
- Jiang C., Schommer C.K., Kim S.Y., Suh D.Y. Cloning and characterization of chalcone synthase from the moss, Physcomitrella patens. Phytochemistry, 2006, 67(23): 2531-2540 (doi: 10.1016/j.phytochem.2006.09.030).
- She H., He S., Zhou Z., Zhang Q. Molecular cloning and sequences analysis of chalcone synthase gene from Fagopyrum tataricum. In: Informatics and management science I /W. Du (ed.). Springer, London, 2013 (doi: 10.1007/978-1-4471-4802-9_85).
- Takeuchi A., Matsumoto S., Hayatsu M. Chalcone synthase from Camellia sinensis: isolation of the cDNAs and the organ-specific and sugar-responsive expression of the genes. Plant and Cell Physiology, 1994, 35(7): 1011-1018 (doi: 10.1093/oxfordjournals.pcp.a078688).
- Mamati G.E., Liang Y., Lu J. Expression of basic genes involved in tea polyphenol synthesis in relation to accumulation of catechins and total tea polyphenols. Journal of the Science of Food and Agriculture, 2006, 86(3): 459-464 (doi: 10.1002/jsfa.2368).
- Wilmouth R.C., Turnbull J.J., Welford R.W., Clifton I.J., Prescott A.G., Schofield C.J. Structure and mechanism of anthocyanidin synthase from Arabidopsis thaliana. Structure, 2002, 10(1): 93-103 (doi: 10.1016/s0969-2126(01)00695-5).
- Bogs J., Downey M.O., Harvey J.S., Ashton A.R., Tanner G.J., Robinson S.P. Proanthocyanidin synthesis and expression of genes encoding leucoanthocyanidin reductase and anthocyanidin reductase in developing grapeberries and grapevine leaves. Plant Physiology, 2005, 139(2): 652-663 (doi: 10.1104/стр.105.064238).
- Tanner G.J., Francki K.T., Abrahams S., Watson J.M., Larkin P.J., Ashton A.R. Proanthocyanidin biosynthesis in plants: purification of legume leucoanthocyanidin reductase and molecular cloning of its cDNA. Journal of Biological Chemistry, 2003, 278(34): 31647-31656 (doi: 10.1074/jbc.M302783200).
- Saito K., Kobayashi M., Gong Z., Tanaka Y., Yamazaki M. Direct evidence for anthocyanidin synthase as a 2‐oxoglutarate‐dependent oxygenase: molecular cloning and functional expression of cDNA from a red forma of Perilla frutescens. The Plant Journal, 17(2): 181-189 (doi: 10.1046/j.1365-313x.1999.00365.x).
- Punyasin P.A.N., Abeysinghe I.S.B., Kumar V., Treutter D., Duy D., Gosch C., Martens S., Forkmann G., Fischer T.C. Flavonoid biosynthesis in the tea plant Camellia sinensis: properties of enzymes of the prominent epicatechin and catechin pathways. Archives of Biochemistry and Biophysics, 2004, 431(1): 22-30 (doi: 10.1016/j.abb.2004.08.003).
- Xie D.Y., Sharma S.B., Dixon R.A. Anthocyanidin reductases from Medicago truncatula and Arabidopsis thaliana. Archives of Biochemistry and Biophysics, 2004, 422(1): 91-102 (doi: 10.1016/j.abb.2003.12.011).
- Zhang L.Q., Wei K., Cheng,H., Wang L.Y., Zhang, C.C. Accumulation of catechins and expression of catechin synthetic genes in Camellia sinensis at different developmental stages. Botanical Studies, 2016, 57(1): 31 (doi: 10.1186/s40529-016-0143-9).
- Liu M., Tian H., Wu J., Cang R., Wang R., Qi X., Xu Q., Chen X. Relationship between gene expression and the accumulation of catechin during spring and autumn in tea plants (Camellia sinensis L.). Horticulture Research, 2015, 2: 15011 (doi: 10.1038/hortres.2015.11).
- Nesi N., Jond C., Debeaujon I., Caboche M., Lepiniec L. The Arabidopsis TT2 gene encodes an R2R3 MYB domain protein that acts as a key determinant for proanthocyanidin accumulation in developing seed. The Plant Cell, 2001, 13(9): 2099-2114 (doi: 10.1105/TPC.010098).
- Taylor L.P., Grotewold E. Flavonoids as developmental regulators. Current Opinion in Plant Biology, 2005, 8(3): 317-323 (doi: 10.1016/j.pbi.2005.03.005).
- Baudry A., Caboche M., Lepiniec L. TT8 controls its own expression in a feedback regulation involving TTG1 and homologous MYB and bHLH factors, allowing a strong and cell‐specific accumulation of flavonoids in Arabidopsis thaliana. The Plant Journal, 2006, 46(5): 768-779 (doi: 10.1111/j.1365-313X.2006.02733.x).
- Ravaglia D., Espley R.V., Henry-Kirk R.A., Andreotti С., Ziosi V., Hellens R.P., Costa G., Allan A.C. Transcriptional regulation of flavonoid biosynthesis in nectarine (Prunus persica) by a set of R2R3 MYB transcription factors. BMC Plant Biology, 2013, 13: 68 (doi: 10.1186/1471-2229-13-68).
- Li P., Xia E., Fu J., Xu Y., Zhao X., Tong W., Tang Q., Tadege M., Fernie A.R., Zhao J. Diverse roles of MYB transcription factors in regulating secondary metabolite biosynthesis, shoot development, and stress responses in tea plants (Camellia sinensis). The Plant Journal, 2022, 110(4): 1144-1165 (doi: 10.1111/tpj.15729).
- Song S., Tao Y., Gao L., Liang H., Tang D., Lin J., Wang Y., Gmitter F.G. Jr., Li C. An integrated metabolome and transcriptome analysis reveal the regulation mechanisms of flavonoid biosynthesis in a purple tea plant cultivar. Frontiers in Plant Science, 2022, 13: 880227 (doi: 10.3389/fpls.2022.880227).
- Mohanpuria P., Kumar V., Yadav S.K. Tea caffeine: metabolism, functions, and reduction strategies. Food Science and Biotechnology, 2010, 19(2): 275-287 (doi: 10.1007/s10068-010-0041-y).
- Ashihara H., Kubota H. Patterns of adenine metabolism and caffeine biosynthesis in different parts of tea seedlings. Physiologia Plantarum, 1986, 68(2): 275-281 (doi: 10.1111/j.1399-3054.1986.tb01926.x).
- Negishi O., Ozawa T., Imagawa H. Biosynthesis of caffeine from purine nucleotides in tea plant. Bioscience, biotechnology, and biochemistry, 1992, 56(3): 499-503 (doi: 10.1271/bbb.56.499).
- Yoneyama N., Morimoto H., Ye C.X., Ashihara H., Mizuno K., Kato M. Substrate specificity of N-methyltransferase involved in purine alkaloids synthesis is dependent upon one amino acid residue of the enzyme. Molecular Genetics and Genomics, 2006, 275(2): 125-135 (doi: 10.1007/s00438-005-0070-z).
- Ping L., Ren Q., Kang X., Zhang Y., Lin X., Li B., Gao X., Chen Z. Isolation and functional analysis of promoter for N-methyltransferase gene associated with caffeine biosynthesis in tea plants (Camellia sinensis). Journal of Tea Science, 2018, 38(6): 569-579.
- Kato M., Mizuno K., Crozier A., Fujimura T., Ashihara H. Caffeine synthase gene from tea leaves. Nature, 2000, 406: 956-957 (doi: 10.1038/35023072).
- Li P., Ye Z., Fu J., Xu Y, Shen Y, Zhang Y., Tang D., Li P., Zuo H., Tong W., Wang S., Fernie A.R., Zhao J. CsMYB184 regulates caffeine biosynthesis in tea plants. Plant Biotechnology Journal, 2022, 20(6): 1012-1014 (doi: 10.1111/pbi.13814).
- Yamada Y., Sato F. Transcription factors in alkaloid biosynthesis. International review of cell and molecular biology, 2013, 305: 339-382 (doi: 10.1016/B978-0-12-407695-2.00008-1).
- Shi D., Winston J.H., Blackburn M.R., Datta S.K., Hanten G., Kellems R.E. Diverse genetic regulatory motifs required for murine adenosine deaminase gene expression in the placenta. Journal of Biological Chemistry, 1997, 272(4): 2334-2341 (doi: 10.1074/jbc.272.4.2334).
- Zhu B., Chen L.B., Lu M., Zhang J., Han J., Deng W.W., Zhang Z.Z. Caffeine content and related gene expression: novel insight into caffeine metabolism in camellia plants containing low, normal, and high caffeine concentrations. Agricultural and Food Chemistry, 2019, 67(12): 3400-3411 (doi: 10.1021/acs.jafc.9b00240).
- Ma W., Kang X., Liu P., She K., Zhang Y., Lin X., Li B., Chen Z. The NAC-like transcription factor CsNAC7 positively regulates the caffeine biosynthesis-related gene yhNMT1 in Camellia sinensis. Horticulture research, 2022, 9: uhab046 (doi: 10.1093/hr/uhab046).
- Hara Y., Luo S.J., Wikramasinghe R.L., Yamanishi T. Special issue on tea. Food Reviews International, 1995, 11: 371-545.
- Harbowy M.E., Balentine D.A., Davies A.P., Cai Y. Tea chemistry. Critical Reviews in Plant Sciences, 1997, 16(5): 415-480 (doi: 10.1080/07352689709701956).
- Thippeswamy R., Mallikarjun Gouda K.G., Rao D.H., Martin A., Gowda L.R. Determination of theanine in commercial tea by liquid chromatography with fluorescence and diode array ultraviolet detection. Journal of Agricultural and Food Chemistry, 2006, 54(19): 7014-7019 (doi: 10.1021/jf061715+).
- Mu W., Zhang T., Jiang B. An overview of biological production of L-theanine. Biotechnology Advances, 2015, 33(3-4): 335-342 (doi: 10.1016/j.biotechadv.2015.04.004).
- Chen Z., Wang Z., Yuan H., He N. From tea leaves to factories: a review of research progress in L-theanine biosynthesis and production. Journal of Agricultural and Food Chemistry, 2021, 69(4): 1187-1196 (doi: 10.1021/acs.jafc.0c06694).
- Lea P.J., Blackwell R.D., Chen F.L., Hecht U. Enzymes of ammonia assimilation. In: Methods in plant biochemistry / P.J. Lea (ed.). Academic Press, London, 1990 (doi: 10.1016/B978-0-12-461013-2.50022-8).
- Crocomo O., Fowden L. Amino acid decarboxylases of higher plants: the formation of ethylamine. Phytochemistry, 1970, 9(3): 537-540 (doi: 10.1016/S0031-9422(00)85685-X).
- Smith T. The occurrence, metabolism and functions of amines in plants. Biological Reviews, 1971, 46(2): 201-241 (doi: 10.1111/j.1469-185x.1971.tb01182.x).
- Takeo T. L-Alanine as a precursor of ethylamine in Camellia sinensis. Phytochemistry, 1974, 13(8): 1401-1406 (doi: 10.1016/0031-9422(74)80299-2).
- Wen B., Luo Y., Liu D., Zhang X., Peng Z., Wang K., Lia J., Huang J., Liu Zh. The R2R3-MYB transcription factor CsMYB73 negatively regulates L-theanine biosynthesis in tea plants (Camellia sinensis L.). Plant Science, 2020, 298: 110546 (doi: 10.1016/j.plantsci.2020.110546).
- Cheng H., Wu W., Liu X., Wang Y., Xu P. Transcription factor CsWRKY40 regulates L-theanine hydrolysis by activating the CsPDX2.1 promoter in tea leaves during withering. Horticulture Research, 2022, 9: uhac025 (doi: 10.1093/hr/uhac025).
- Duan X., Hu X., Chen F., Deng Z. Bioactive ingredient levels of tea leaves are associated with leaf Al level interactively influenced by acid rain intensity and soil Al supply. Journal of Food Agriculture and Environment, 2012, 10: 1197-1204.
- Ahmed S., Stepp J.R., Orians C., Griffin T., Matyas C., Robbat A., Cash S., Xue D., Long C., Unachukwu U., Buckley S., Small D., Kennelly E. Effects of extreme climate events on tea (Camellia sinensis) functional quality validate indigenous farmer knowledge and sensory preferences in tropical China. PLoS ONE, 2014, 9(10): e109126 (doi: 10.1371/journal.pone.0109126).
- Cao H., Wang L., Yue C., Hao X., Wang X., Yang Y. Isolation and expression analysis of 18 CsbZIP genes implicated in abiotic stress responses in the tea plant (Camellia sinensis). Plant Physiology and Biochemistry, 2015, 97: 432-442 (doi: 10.1016/j.plaphy.2015.10.030).
- Liu S.-C., Jin J.-Q., Ma J.-Q., Yao M.-J., Ma C.-L., Li C.-F., Ding Z.-T., Chen L. Transcriptomic analysis of tea plant responding to drought stress and recovery. PLoS ONE, 2016, 11(1): e0147306 (doi: 10.1371/journal.pone.0147306).
- Chen J., Gao T., Wan S., Zhang Y., Yang J., Yu Y., Wang W. Genome-wide identification, classification and expression analysis of the HSP gene superfamily in tea plant (Camellia sinensis). International Journal of Molecular Sciences, 2018, 19(9): 2633 (doi: 10.3390/ijms19092633).
- Cui X., Wang Y.X., Liu Z.W., Wang W.L., Li H., Zhuang J. Transcriptome-wide identification and expression profile analysis of the bHLH family genes in Camellia sinensis. Functional & Integrative Genomics,2018, 18(5): 489-503 (doi: 10.1007/s10142-018-0608-x).
- Zhu B., Chen L.B., Lu M., Zhang J., Han J., Deng W.W., Zhang Z.Z. Caffeine content and related gene expression: novel insight into caffeine metabolism in Camellia plants containing low, normal, and high caffeine concentrations. Journal of Agricultural and Food Chemistry, 2019, 67(12): 3400-3411 (doi: 10.1021/acs.jafc.9b00240).
- Hu Z., Ban Q., Hao J., Zhu X., Cheng Y., Mao J., Lin M., Xia E., Li Y. Genome-wide characterization of the C-repeat Binding Factor (CBF) gene family involved in the response to abiotic stresses in tea plant (Camellia sinensis). Frontiers in Plant Science, 2020, 11: 921 (doi: 10.3389/fpls.2020.00921).
- Maritim T., Kamunya S., Mwendia C. Mireji P., Muoki R., Wamalwa M., Francesca S., Schaack S., Kyalo M., Wachira F. Transcriptome-based identification of water-deficit stress responsive genes in the tea plant, Camellia sinensis. Journal of Plant Biotechnology, 2016, 43(3): 302-310 (doi: 10.5010/JPB.2016.43.3.302).
- Parmar R., Seth R., Singh P., Singh G., Kumar S., Sharma R.K. Transcriptional profiling of contrasting genotypes revealed key candidates and nucleotide variations for drought dissection in Camellia sinensis (L.) O. Kuntze. Scientific Reports, 2019, 9: 7487 (doi: 10.1038/s41598-019-43925-w).
- Zhang W.J., Liang Y.R., Zhang F.Z., Chen C.S., Zhang Y.G., Chen R.B., Weng B.Q. Effects on the yield and quality of oolong tea by covering with shading net. Journal of Tea Science, 2004, 4: 276-282 (doi: 10.13305/j.cnki.jts.2004.04.010).
- Wang R., Zhou W., Jiang X. Reaction kinetics of degradation and epimerization of epigallocatechin gallate (EGCG) in aqueous system over a wide temperature range. Journal of Agricultural & Food Chemistry,2008, 56(8): 2694-2701 (doi: 10.1021/jf0730338).
- Zheng X.Q., Jin J., Chen H., Du Y.Y., Ye J.H., Lu J.L., Lin C., Dong J.J., Sun Q.L., Wu L.Y., Liang Y.R. Effect of ultraviolet B irradiation on accumulation of catechins in tea (Camellia sinensis (L) O. Kuntze). African Journal of Biotechnology, 2008, 7(18): 3283-3287.
- Jeyaramraja P.R., Pius P.K., Kumar R.R., Jayakumar D. Soil moisture stress-induced alterations in bio constituents determining tea quality. Journal of the Science of Food and Agriculture, 2003, 83(12): 1187-1191 (doi: 10.1002/jsfa.1440).
- Притула З.В., Великий А.В., Малюкова Л.С. Влияние мезо- и микроудобрений на качество чайного сырья в условиях Черноморского побережья России. Плодоводство и ягодоводство России, 2014, 38(2): 52-58.
- Ding Z., Jia S., Wang Y., Xiao J., Zhang Y. Phosphate stresses affect ionome and metabolome in tea plants. Plant Physiology and Biochemistry, 2017, 120: 30-39 (doi: 10.1016/j.plaphy.2017.09.007).
- Wang W., Xin H., Wang M., Ma Q., Wang L., Kaleri N.A., Wang Y., Li X. Transcriptomic analysis reveals the molecular mechanisms of drought-stress-induced decreases in Camellia sinensis leaf quality. Frontiers in Plant Science, 2016, 7: 385 (doi: 10.3389/fpls.2016.00385).
- Wang Y.X., Liu Z.W., Wu Z.J., Li H., Zhuang J. Transcriptome-wide identification and expression analysis of the NAC gene family in tea plant [Camellia sinensis (L.) O. Kuntze]. PLoS ONE, 2016, 11(11): e0166727 (doi: 10.1371/journal.pone.0166727).
- Chen X.H., Zhuang C.G., He Y.F., Wang L., Han G.Q., Chen C., He H.Q. Photosynthesis, yield, and chemical composition of Tieguanyin tea plants (Camellia sinensis (L.) O. Kuntze) in response to irrigation treatments. Agricultural Water Management, 2010, 97(3): 419-425 (doi: 10.1016/j.agwat.2009.10.015).
- Bai P., Wei K., Wang L., Zhang F., Ruan L., Li H., Wu L., Cheng H. Identification of a novel gene encoding the specialized alanine decarboxylase in tea (Camellia sinensis). Molecules, 2019, 24(3): 540 (doi: 10.3390/molecules24030540).
- Dong F., Hu J., Shi Y., Liu M., Zhang Q., Ruan J. Effects of nitrogen supply on flavonol glycoside biosynthesis and accumulation in tea leaves (Camellia sinensis). Plant Physiology and Biochemistry, 2019, 128: 48-57 (doi: 10.1016/j.plaphy.2019.02.017).
- Kovalcik J., Klejdus B. Induction of phenolic metabolites and physiological changes in chamomile plants in relation to nitrogen nutrition. Food chemistry, 2014, 142: 334-341 (doi: 10.1016/j.foodchem.2013.07.074).
- Liu M.Y., Burgos A., Zhang Q., Tang D., Shi Y., Ma L., Yi X., Ruan J. Analyses of transcriptome profiles and selected metabolites unravel the metabolic response to NH4+ and NO3− as signaling molecules in tea plant (Camellia sinensis L.). Scientia Horticulturae, 2017, 218: 293-303 (doi: 10.1016/J.SCIENTA.2017.02.036).
- Lync J., Jonathan P., Clair S.B.S. Mineral stress: the missing link in understanding how global climate change will affect plants in real world soils. Field Crops Research, 2004, 90(1): 101-115 (doi: 10.1016/J.FCR.2004.07.008).
- Sun L., Liu Y., Wu L., Liao H. Comprehensive analysis revealed the close relationship between N/P/K status and secondary metabolites in tea leaves. ACS Omega, 2019, 4(1): 176-184 (doi: 10.1021/ACSOMEGA.8B02611).
- Wei K., Liu M., Shi Y., Zhang H., Ruan J., Zhang Q., Cao M. Metabolomics reveal that the high application of phosphorus and potassium in tea plantation inhibited amino-acid accumulation but promoted metabolism of flavonoid. Agronomy, 2022, 12(5): 1086 (doi: 10.3390/agronomy12051086).