doi: 10.15389/agrobiology.2021.5.948rus
УДК 633.15:581.192:58.088
Исследования проводятся в Сербии и в Российской Федерации под руководством профессора C.N. Ra-denovich (г. Белград, Сербия) и профессора Г.В. Максимова (г. Москва, Россия) и финансируются национальными министерствами и ведомствами (РНФ для ГВМ № 19-79- 30062).
ИСПОЛЬЗОВАНИЕ ИНФРАКРАСНОЙ СПЕКТРОСКОПИИ И СПЕКТРОСКОПИИ КОМБИНАЦИОННОГО РАССЕЯНИЯ ДЛЯ АНАЛИЗА СОСТОЯНИЯ БИОМОЛЕКУЛ У ЛИНИЙ КУКУРУЗЫ Zea mays L.
C.N. RADENOVICH1, 5✉, Г.В. MAKСИМОВ2 ✉, В.В. ШУТОВА3, J. HAO4, N.S. DELICH1, M.D. SECHANSKY1, A.S. POPOVICH1
В настоящее время мало неинвазивных методов, которые позволяют контролировать содержание и конформацию молекул биоактивных органических соединений в клетках и тканях растений не только в лабораторных, но и в полевых условиях. Инфракрасная спектроскопия и спектроскопия комбинационного рассеяния (ИК- и КР-спектроскопия) активно используется для анализа роли молекул некоторых веществ в селекции сельскохозяйственных культур. С помощью методов вибронной спектроскопии мы исследовали изменения содержания и конформации молекул хлоропластов у различных линий кукурузы. Методом ИК-спектроскопии (область 3500-3000 см-1) установлено, что в воде, углеводах и белках хлоропластов доля колебаний ОН-групп и внутри- и межмолекулярных Н-связей максимальна у линии кукурузы ZPPL 186, а колебаний NH-групп амидов (белки) минимальна у линии ZPPL 225. Доказано, что для линии ZPPL 186 характерна максимальная доля колебаний от молекул алканов, карбоновых кислот (область 2920-2860 см-1) и деформационных колебаний ароматических структур (полоса 1000 см-1), находящихся в хлоропластах, тогда как для линии M1-3-3-sdms — доля валентных колебания O=C=O связей (полоса 2300 см-1). С использованием КР-спектроскопии (области 1250-500 см-1 и 1535-1400 см-1) обнаружено, что различия в спектрах между линиями кукурузы связаны с выявленными изменениями конформации молекул каротиноидов в хлоропластах, но не молекул целлюлозы. Установлено, что у всех образцов (кроме ZPPL 225) молекулы каротиноидов находятся в 15-trans форме с различной конформацией полиеновой цепи. Отметим, что у линии ZPPL 186 конформация каротиноидов, находящихся в хлоропластах, характеризуется минимальной величиной поворота вне плоскости полиеновой цепи и более выраженными колебаниями боковой СН3-группы. Высказывает предположение, что у изученных линий кукурузы каротиноиды хлоропластов листа не взаимодействуют с ароматическими аминокислотами белков. По мнению авторов, сочетание методов ИК- и КР-спектроскопии хлоропластов листа можно рекомендовать для контроля за содержанием и конформациоными изменениями биомолекул при селекции кукурузы.
Ключевые слова: Zea mays L., инбредная линия, лист, хлоропласты, спектры комбинационного рассеяния, инфракрасные спектры, конформационные изменения, углеводы, каротиноиды, белки, селекционное тестирование.
Рост населения на планете повышает требования к урожайности и рентабельности производства продуктов сельского хозяйства, что может быть достигнуто благодаря фундаментальным исследованиям, позволяющим разработать эффективные аналитические методы для применения в селекции растений (1-3). В настоящее время мало неинвазивных методов, которые позволяют контролировать содержание и изменение конформации молекул биоактивных органических соединений в клетках и тканях растений не только в лабораторных, но и в полевых условиях. ИК- и КР-спектроскопия активно используется для изучения роли молекул некоторых веществ в селекции сельскохозяйственных культур (3-6). В большинстве случаев измерения с помощью указанных методов можно проводить в полевых условиях непосредственно на тканях растений, а также на фракциях, выделенных из растительного материала. Оба метода вибронной спектроскопии позволяют не только регистрировать спектры, полосы которых характеризуют содержание и конформационные изменения определенных молекул (7-9), но и на основании этих данных различать генотипы растений (1, 2). Возможность быстрого мониторинга количества и физико-химического состояния биоактивных органических соединений и клеточных компартментов (например, каротиноидов и хлоропластов) в гомогенатах листьев растений и выявление корреляции этих показателей с хозяйственно значимыми признаками позволило бы использовать регистрируемые при спектроскопии параметры как маркеры для оценки агрономической эффективности исследуемых форм при селекции растений. Кроме того, эти методы могут применяться в обрабатывающей промышленности для быстрой проверки качества поступающего сырья, а также для контроля на непрерывных производствах (10-12). На основе анализа ИК-спектров у растений были получены дополнительные характеристики не только структуры различных молекул, но и модификации их конформации (по изменению доли характерных колебаний химических связей в молекулах) (13-15).
В отличие от ИК-спектроскопии, КР-спектроскопия позволяет контролировать изменения структуры молекул в клетках и тканях без высушивания препаратов (3, 12, 16-18). С помощью этого метода описаны особенности и различия в распределении молекул белков и пигментов в клетках растений у разных гибридов и линий кукурузы. Важно, что такой подход позволяет выявить наличие положительной корреляции между изменением флуоресценции хлорофилла в хлоропластах и содержанием и структурой молекул каротиноидов в целых листьях у линий и гибридов кукурузы (3, 18).
Очевидно, что формирование методологии исследования функционального состояния растения с помощью спектральных методов позволит выявить новые молекулярные механизмы, которые могут быть использованы в селекции кукурузы.
В настоящей работе доказано, что совместное использование ИК- и КР-спектроскопии листа растения можно рекомендовать в качестве малоинвазивного метода контроля за содержанием и возможными различиями в конформации биологических молекул при тестировании линий и гибридов кукурузы.
Цель работы заключалась в комплексном анализе изменений содержания и конформации молекул белков, углеводов и пигментов в хлоропластах листа у инбредных линий кукурузы методами ИК- и КР-спектроскопии.
Методика. Эксперименты выполняли на линиях кукурузы (Zea mays L.) ZPPL 186, ZPPL 225, M1-3-3-sdms, оригинатор — Maize Research Institute, Zemun Polje, г. Белград, Сербия), которые характеризуются высокими показателями качества, урожайности и адаптированы к технологиям возделывания (1, 3, 12). По 30 семян каждой линии (масса соответственно от 313±9, 382±17 и 196±5 мг) обрабатывали в течение 30 мин перекисью водорода, промывали водой и оставляли в чашке Петри (22 °С, постоянное бесконтактное смачивание водой) до появления корешков длиной не менее 5 мм, после чего проросшие семена помещали в грунт и выращивали до фазы 3-го листа (16-часовой световой день; лабораторный опыт). Качество используемых семян оценивали по всхожести (способность семян прорастать и давать нормально развитые проростки при определенных условиях за принятые сроки; ГОСТ 13056.6-75) и энергии прорастания (способность семян прорастать за более короткий срок — от 1 до 15 сут; ГОСТ 13056.6-75).
Для получения хлоропластов охлажденную (0-4 °С) навеску листьев (5 г, образцы находились в полиэтиленовом пакете или во влажной фильтровальной бумаге), отделяли от жилок, измельчали ножницами и гомогенизировали при 0-4 °С трижды (по 10 с) в охлажденном буфере (4 °С) (0,04 М сахароза, 20 мМ Tris-HCl, рН 7,8, 35 мМ NaCl, 1 мМ EDTA). Гомогенат фильтровали через четыре слоя капрона и центрифугировали (1500 об/мин, 5 мин, 0-4 °С). Супернатант переливали в предварительно охлажденные пробирки и снова центрифугировали (5000 об/мин, 44 °С, 10 мин). Супернатант сливали, полученный осадок ресуспендировали в 5 мл среды выделения. Добавляли глицерин (30 % от полученного объема) и замораживали при -73 °С. Перед регистрацией КР- и ИК-спектров суспензию размораживали и разводили буфером (15 мМ NaCl, 400 мМ сахароза, 50 мМ Mes-NaOH, рН 6,5) в соотношении 10 мкл суспензии на 5 мл буфера (19, 20).
Целлюлозу выделяли из листьев кукурузы согласно описанию (21) с использованием 3 % раствора гипохлорита натрия, 5 % раствора пероксида водорода и смеси 3 % раствора гипохлорита натрия и метанола.
Комбинационное рассеяние каротиноидов в хлоропластах листа регистрировали с помощью КР-спектрометра ДФС 24 (АО «ЛОМО», Россия) с лазером (Ciel, «Eurolaser GmbH», Германия) (длина волны 473 нм), системой регистрации МОРС 1/3648 (ООО «МОРС», г. Троицк, Россия) на базе линейной ПЗС-матрицы TCD1304DG («Toshiba», Япония) с фильтром LPO2-473RS-50 («Semrock», США). Мощность лазера на образце составляла 3 мВт, время регистрации сигнала — 10 с (23).
Регистрацию ИК-спектров в диапазоне 400-4000 см-1 с преобразованием Фурье (ИК-Фурье спектрометрия) проводили с помощью ИК-спектрометра IR-Prestig 21 («Shimadzu Corp.», Япония) с шагом измерения 4 см-1. Перед экспериментом суспензию гомогената листа (см. выше) размораживали и разводили буфером (15 мМ NaCl, 400 мМ сахароза и 50 мМ Mes-NaOH, рН 6,5; 10 мкл суспензии на 5 мл буфера), высушивали и закатывали в таблетку с бромидом калия (KBr) (соотношение компонентов 1:50) (12, 23).
Статистическую обработку результатов проводили с помощью программы Microsoft Excel 2013 («Microsoft Corp.», США) и пакета Statistica v.10 («StatSoft, Inc.», США). Первичнаую обработку КР- и ИК-спектров проводили с помощью пакета Origin Pro 2017 («OriginLab Corp.», США). Проверка статистических гипотез осуществлялась непараметрическим H-критерием для множества независимых переменных Краскела-Уоллиса (Kruskal-Wallis test) (уровень значимости р = 0,05; n = 10).
Результаты. Задачи нашего исследования включали регистрацию ИК-спектров хлоропластов у трех линий кукурузы и анализ различий между ними для молекул белков и углеводов, а также проведение дополнительно КР-спектроскопии для выявления изменений каротиноидов, содержащихся в хлоропластах. В ИК-спектрах хлоропластов в диапазоне 4000-400 см-1 были обнаружены полосы, обусловленные колебаниями связей в молекулах ряда органических соединений, включая целлюлозу, белки, углеводы, эфиры, фенолы (рис. 1).
Было установлено, что в ИК-спектре хлоропластов в области 3700-3100 см-1 присутствовали полосы, соответствующие колебаниям гидроксильных групп, а в области 1500-900 см-1 — колебаниям С–Н, С–О и О–Н групп гликозидной связи и глюкопиранозного кольца молекул целлюлозы (13-15). Так, полоса 3340 см-1 ИК-спектра характеризует колебания О–Н и СН2–ОН связей целлюлозы, полосы 2900 и 1374 см-1 — деформационные колебания С–Н групп, а полосы 1170 и 1059 см-1 — колебания соответственно связей С–О–С и С–ОН. В ИК-спектре хлоропластов листа присутствовала полоса 3414 см-1, обусловленная валентными колебаниями гидроксильных групп, и полоса при 2904 см-1, характеризующая колебания С–Н метиленовых и метиновых групп молекул. Также в ИК-спектре выявили полосу 1654 см-1, обусловленную Н–О–Н колебаниям в кристал-лизованной воде, полосу 1375 см-1, вызванную деформационными колебаниями С–Н связи, и полосу 1317 см-1, которая характеризует деформационные колебания СН2-групп молекул. Полоса 1165 см-1 в ИК-спектре соответствует валентным колебаниям С–О–С связи, однако ее также относят к деформационным колебаниям С–О или О–Н в С–ОН группах молекул. Полоса ИК-спектра при 1085 см-1 соответствует колебаниям связи С–О–С в глюкопиранозном кольце, полоса 1058 см-1 характеризует валентное колебание С–О связи в С3Н–ОН группе, а полосы 796 и 777 см-1 отражают колебания глюкопиранозного кольца, связанные с колебаниями СН- и СН2-групп. Полосы, выявленные в ИК-спектрах в области 1500-1650 см-1, соответствуют белкам.
Мы обнаружили, что колебания свободной и связанной ОН-группы и колебания внутри- и межмолекулярных Н-связей в хлоропластах листа, а также валентные колебания N–H связей в первичных и вторичных амидах белков, колебания связей гидроксильной OH-группы (вода, углеводы, аминокислоты) и валентные колебания связей NH-группы (белки, аминокислоты и их производные) максимальны для линии ZPPL 186 и минимальны для линии ZPPL 225 (область 3500-3000 см-1). У линии ZPPL 186 для хлоропластов листа также характерен максимальный вклад валентных колебаний алканов, карбоновых кислот (область 2920-2860 см-1) и деформационных колебаний связей в ароматических соединениях (полоса 1000 см-1), у линии M1-3-3-sdms — колебаний связей O–C–O (полоса 2300 см-1). Таким образом, нами выявлены различия в составе молекул белков и углеводов в хлоропластах листа различных линий кукурузы. Эти различия могут быть обусловлены синтезом новых молекул целлюлозы, белков, углеводов, эфиров, фенолов. Например, большее содержание молекул алканов, карбоновых кислот, а также ароматических соединениях в хлоропластах листа было характерно для ZPPL 186.
Однако с помощью ИК-спектроскопии мы не смогли выявить различия в области от 1100 до 1600 см-1 (см. рис. 1). Известно, что эта область характерна для симметричных и антисимметричных валентных колебаний атомов, а также плоскостных и внеплоскостных деформационных колебаний молекул (14 15). Возможными кандидатами для таких различий могли быть молекулы целлюлозы и каротиноидов из хлоропластов листа. Для изученных нами линий кукурузы характерно высокое содержание крахмала, целлюлозы и каротиноидов в тканях листа (22). Поэтому в следующей серии экспериментов мы получили ИК-спектры целлюлозы, которые характеризовались полосами в области 4000-2500 см-1 (валентные колебания О–Н и С–Н связей) и полосами в области 1500-500 см-1 (область валентных колебаний С–С, С–О связей и деформационные колебания С–Н и О–Н связей). Область колебаний ОН-групп включала три полосы, соответствующие свободным гидроксильным группам (у атомов углерода С2 и С6) и ОН-группе, которые вовлечены в водородные связи (рис. 2).
Внутримолекулярные водородные связи между гидроксильными группами в положениях С2 и С6 (равно как и в С3) образуются даже в присутствии малых количеств ОН-групп. В ИК-спектре в области 1500-900 см-1 мы регистрировали суммарную интенсивность колебаний трех гидроксильных групп, входящих в состав каждого глюкопиранозного звена мо-лекулы целлюлозы (колебания С–Н, С–О и О–Н связей), а полосы при 1375 и 1319 см-1 характеризовали наличие и проявление деформационных колебаний соответственно С–Н- и СН2-групп в молекуле целлюлозы (8, 14). Однако с помощью ИК-спектроскопии нам не удалось обнаружить различия в исследуемой области волновых чисел для молекул целлюлозы.
В следующей серии экспериментов для уточнения результатов ИК-спектроскопии в области 800-1800 см-1 мы использовали КР-спектроскопию. Важно, что именно КР-спектры хлоропластов листа кукурузы в указанной области характеризуются высокоамплитудными полосами каротиноидов и низкоамплитудными или совмещенными полосами молекул целлюлозы (полоса 1095 см-1 соответствует колебаниям связей С–О–С, а 1477 см-1 — колебаниям Н–С–Н групп атомов молекулы целлюлозы). Таким образом, с помощью КР-спектроскопии в области 800-1800 см-1 можно дополнительно исследовать конформацию молекул каротиноидов в хлоропластах (см. рис. 2, табл.) (24-27). В КР-спектрах полоса 1523 см-1 характеризует колебания связей С=С в молекуле каротиноида, полоса 1155 см-1 —колебания C–C связей. В случае транс-конформации двойных связей в молекуле каротиноида полоса при 1155 см-1 изменяется и приобретает два выраженных плеча — при 1190-1193 и 1210 см-1. Полоса 1004 см-1 соответствует колебаниям боковой метильной группы C–СН3. Полоса 960 см-1 характеризует внеплоскостные колебания С–Н в области С–С связи, причем увеличение интенсивности этой полосы наблюдается при нарушении плоской конфигурации молекулы: чем большее количество пигмента связано с белком, тем менее выражены внеплоскостные кручения между атомами углерода С11 и С12 и, как следствие, интенсивность указанной полосы в КР-спектре.
При анализе КР-спектров каротиноидов для нормировки вклада каждого типа связей используют соотношения величины пиков, выбирая, как правило, постоянную по амплитуде полосу КР, изменения которой при изучаемом процессе минимальны (внутримолекулярный маркер) (23). Полученные нами результаты указывают на то, что в различных линиях кукурузы каротиноиды в хлоропластах могут находиться в разных конформационных состояниях. Так, конформация, обусловленная длиной полиеновой цепи молекул каротиноида (величина пропорциональна I1523/I1155), в хлоро-пластах листа линий кукурузы может варьировать от 0,88 (M1-3-3-sdms) до 1,09 (ZPPL 186), и у двух образцов линий кукурузы (кроме ZPPL 225) молекулы каротиноидов находятся в 15-trans форме. При такой конформации молекулы полоса 1155 см-1 КР-спектра характеризуется одним выраженным плечом 1190 см-1. Отметим, что у линии ZPPL 186 каротиноиды в хлоропластах листа характеризовались минимальным соотношением I960/I1006, что указывает на незначительное изменение конформации, обусловленное поворотом молекулы каротиноида вне плоскости полиеновой цепи, или на отсутствие такого изменения. У линии ZPPL 186 молекула каротиноидов характеризовалась более выраженными колебаниями боковой СН3-группы. Известно, что каротиноид, связанный с белками светособирающего комплекса фотосистемы II, характеризуется интенсивной полосой при 960 см-1 в КР-спектре, которая по амплитуде практически не отличается от полос 1156 и 1004 см-1 (24-27). Отметим, что в наших экспериментах амплитуда полосы 960 см-1 КР-спектра каротиноидов была существенно меньше амплитуды полос при 1156 и 1004 см-1 и не различалась в КР-спектре хлоропластов у разных линий, что, вероятно, свидетельствует об отсутствии белок-липидных взаимодействий.
По мнению авторов, совместное использование ИК- и КР-спектроскопии фракции хлоропластов листа можно рекомендовать в качестве малоинвазивного для растения метода контроля за содержанием и возможными различиями в конформации биологических молекул при тестировании линий и гибридов кукурузы (28-30).
Итак, с помощью методов вибронной спектроскопии исследовали изменения содержания и конформации молекул хлоропластов у разных линий кукурузы. Инфракрасная (ИК) спектроскопия (область 3500-3000 см-1) показала, что в молекулах, содержащихся в хлоропластах листа (вода, углеводы, белки), доля колебаний ОН-групп и внутри- и межмолекулярных Н-связей максимальна у линии кукурузы ZPPL 186, а колебаний NH-групп амидов (белки) минимальна у ZPPL 225. Доказано, что для линии ZPPL 186 характерна максимальная доля колебаний от молекул алканов, карбоновых кислот (область 2920-2860 см-1) и деформационных колебаний ароматических структур (полоса 1000 см-1) хлоропластов, а для линии M1-3-3-sdms — доля колебания связей O=C=O (полоса 2300 см-1). С помощью спектроскопии комбинационного рассеяния (КР-спектроскопия) (области 1250-500 см-1 и 1535-1400 см-1) обнаружено, что у изученных линий кукурузы выявленные различия связаны с изменениями конформации молекул каротиноидов в хлоропластах, но не молекул целлюлозы. Установлено, что у двух образцов (кроме ZPPL 225) молекулы каротиноидов находятся в 15-trans форме с различной конформацией полиеновой цепи. Отметим, что конформация каротиноидов линии ZPPL 186 характеризуется минимальной величиной поворота вне плоскости полиеновой цепи, при этом проявляются более выраженные колебания боковой СН3-группы. Высказано предположение, что у изученных линий каротиноиды хлоропластов листа не взаимодействуют с ароматическими аминокислотами белков.
Интенсивность (I) и соотношение интенсивности полос в спектрах комбинационного рассеяния каротиноидов в хлоропластах листьев у изученных линий кукурузы (Zea mays L.) в фазу 3-го листа (n = 10) |
||||
Частотный сдвиг, см-1 (M±SD) |
Среднее значение I (M) |
Cтандартное отклонение (±SD) |
Характерные полосы |
|
положение |
соотношение интенсивности |
|||
Линия M1-3-3-sdms |
||||
1523,6±0,1 |
132,5 |
9,6 |
962/1006 |
0,45 |
1155,6±0,9 |
148,9 |
10,9 |
1006/1157 |
0,34 |
1191,3±0,4 |
46,7 |
3,7 |
1157/1190 |
3,18 |
1004,9±0,9 |
51,9 |
4,2 |
1006/1525 |
0,39 |
960,5±1,3 |
23,5 |
1,9 |
1526/1157 |
0,88 |
Линия ZPPL 186 |
||||
1523,0±0,2 |
213,6 |
26,9 |
962/1007 |
0,38 |
1155,8±0,7 |
195,7 |
23,5 |
1007/1158 |
0,28 |
1190,4±0,4 |
55,4 |
7,3 |
1158/1190 |
3,53 |
1004,0±0,6 |
55,3 |
4,5 |
1007/1526 |
0,25 |
960,1±0,5 |
21,5 |
2,2 |
1526/1158 |
1,09 |
Линия ZPPL 225 |
||||
1523,8±0,3 |
118,2 |
13,6 |
960/1005 |
0,48 |
1155,2±0,6 |
132,1 |
21,2 |
1005/1157 |
0,36 |
1189,7±0,9 |
43,4 |
6,7 |
1157/1189 |
3,04 |
1004,6±0,4 |
47,7 |
5,6 |
1005/1526 |
0,40 |
960,7±0,3 |
23,0 |
2,4 |
1526/1157 |
0,89 |
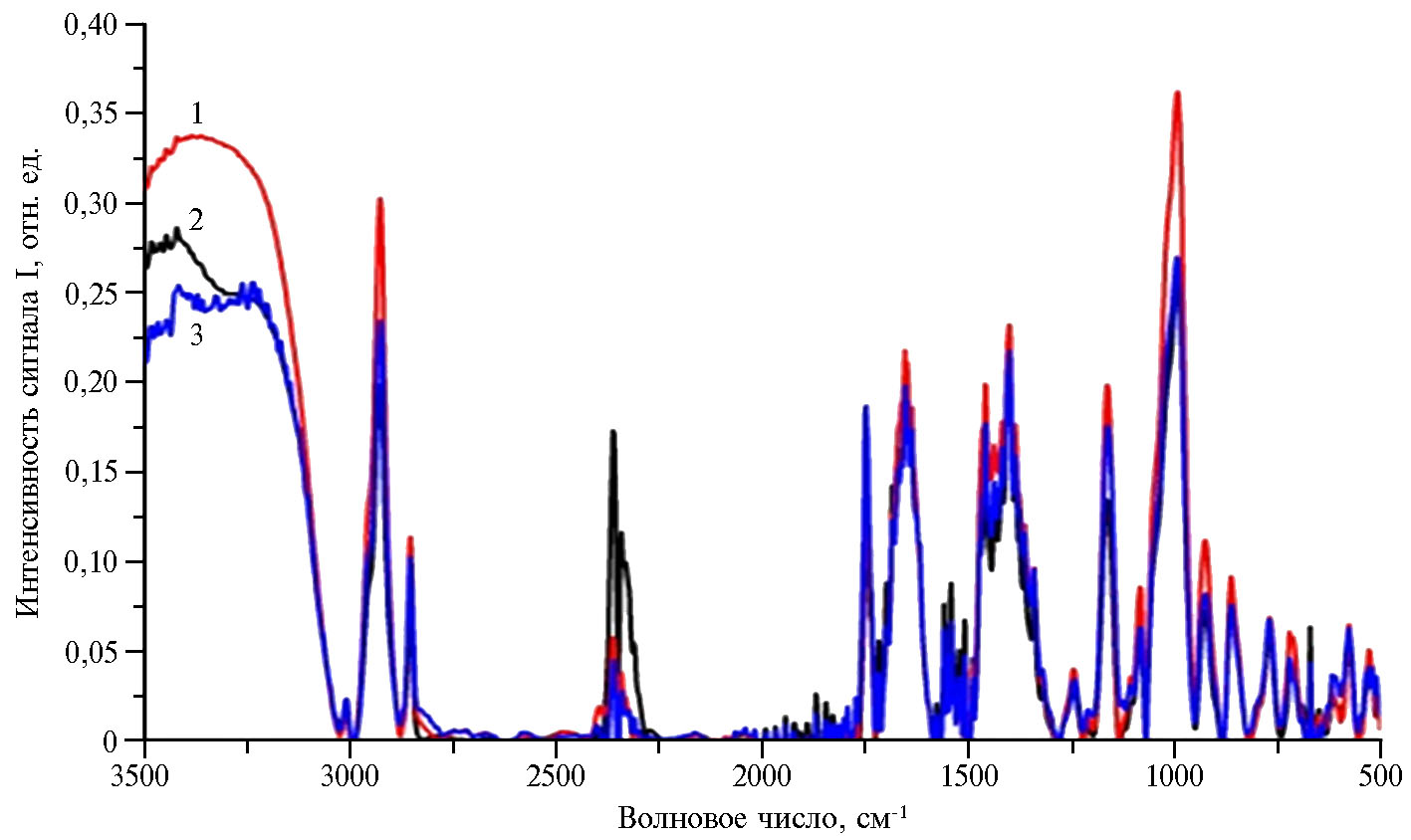
Рис. 1. ИК-спектры (инфракрасная спектроскопия с преобразованием Фурье) хлоропластов листа кукурузы (Zea mays L.) линий ZPPL 186(1), M1-3-3-sdms (2) и ZPPL 225 (3) (оригинатор — Maize Research Institute, Zemun Polje, г. Белград, Сербия) в фазу 3-го листа (лабораторный опыт). Представлены характерные нормированные ИК-спектры.
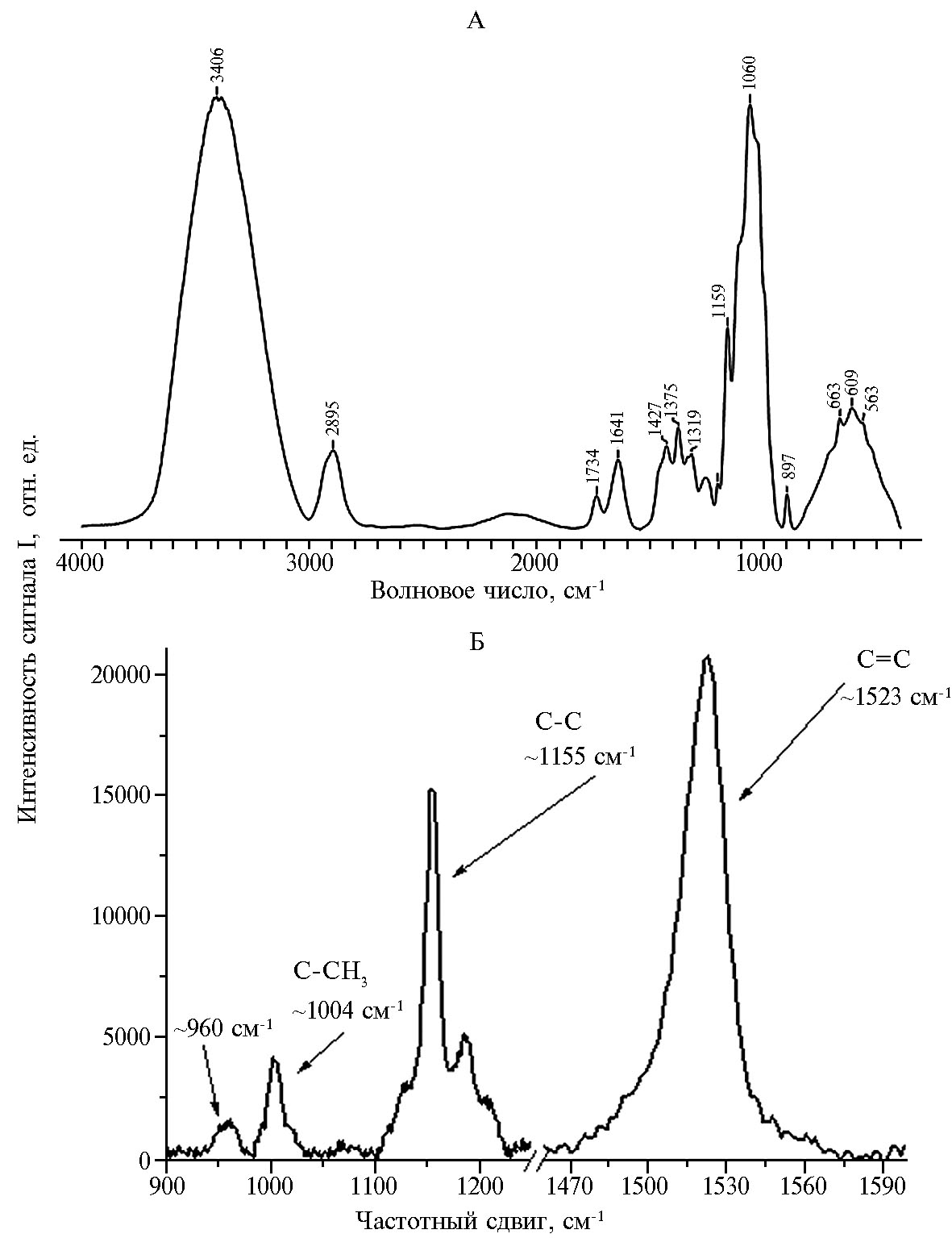
Рис. 2. ИК-спектры целюлозы из ткани листа (инфракрасная спектроскопия с преобразованием Фурье, А) и спектры комбинационного рассеяние (КР) каротиноидов в хлоропластах листьев (Б) у кукурузы (Zea mays L.) линии ZPPL 186 (оригинатор — Maize Research Institute, Zemun Polje, г. Белград, Сербия) в фазу 3-го листа (лабораторный опыт). Представлены характерные нормированные ИК- и КР-спектры.
ЛИТЕРАТУРА
- Radosavljević M., Bekrić V., Božović I., Jakovljević J. Physical and chemical properties of various corn genotypes as a criterion of technological quality. Genetika, 2000, 32(3): 319-329.
- Konstantinov K., Mladeović Drinić S., Abđelkovič V., Babić M. Ethics in scientific results application: gene and life forms patenting. Genetika, 2010, 42(1): 195-208.
- Radenović C.N., Maksimov G.V., Tyutyaev E.V., Stanković G.J., Jovanović Z.V., Beljanski M.V. Detecting the phase transition in thylakoid membranes of maize inbred lines by means of delayed fluorescence. Plant Physiology and Biochemistry, 2014, 81: 208-211 (doi: 10.1016/j.plaphy.2014.03.013).
- Grubbs B.A., Etter N.P., Slaughter W.E., Pittsford A.M., Smith C.R., Schmitt P.D. A low-cost beam-scanning second harmonic generation microscope with application for agrochemical development and testing. Analytical Chemistry, 2019, 91(18): 11723-11730 (doi: 10.1021/acs.analchem.9b02304).
- Butler H.J., Martin F.L., Roberts M.R., Adams S., McAinsh M.R. Observation of nutrient uptake at the adaxial surface of leaves of tomato (Solanum lycopersicum) using Raman spectroscopy. Analytical Letters, 2020, 53(4): 536-562 (doi: 10.1080/00032719.2019.1658199).
- Hu F., Shi L., Min W. Biological imaging of chemical bonds by stimulated Raman scattering microscopy. Nature Methods, 2019, 16(9): 830-842 (doi: 10.1038/s41592-019-0538-0).
- Vlasov A.V., Maliar N.L., Bazhenov S.V., Nikelshparg E.I., Brazhe N.A., Vlasova A.D., Osipov S.D., Sudarev V.V., Ryzhykau Y.L., Bogorodskiy A.O., Zinovev E.V., Rogachev A.V., Manukhov I.V., Borshchevskiy V. I., Kuklin A. I., Pokorný J., Sosnovtseva O., Maksimov G.V., Gordeliy V.I. Raman scattering: from structural biology to medical applications. Crystals, 2020, 10(1): 38 (doi: 10.3390/cryst10010038).
- Schulz H., Baranska M.Identification and quantification of valuable plant substances by IR and Raman spectroscopy. Vibrational Spectroscopy, 2007, 43(1): 13-25 (doi: 10.1016/j.vibspec.2006.06.001).
- Еремина О.Е., Семенова А.А., Сергеева Е.А., Браже Н.А., Максимов Г.В., Шеховцова Т.Н., Гудилин Е.А., Веселова И.А. Спектроскопия гигантского комбинационного рассеяния в современном химическом анализе: достижения и перспективы использования. Успехи химии, 2018, 87(8): 741-770 (doi: 10.1070/RCR4804).
- Schiavon M., Pizzeghello D., Muscolo A., Vaccaro S., Francioso O., Nardi S. High molecular size humic substances enhance phenylpropanoid metabolism in maize (Zea mays L.). J. Chem. Ecol., 2010, 36(6): 662-669 (doi: 10.1007/s10886-010-9790-6).
- Ertani A., Francioso O., Tinti A., Schiavon M., Pizzeghello D., Nardi S. Evaluation of seaweed extracts from Laminaria and Ascophyllum nodosum spp. as biostimulants in Zea mays L. using a combination of chemical, biochemical and morphological approaches. Front Plant Sci., 2018, 9: 428 (doi: 10.3389/fpls.2018.00428).
- Radenović Č.N., Maksimov G.V., Shutova V.V., Slatinskaya O.V., Protopopov F.F., Delić N.S., Grchić N.M., Pavlov J.M., Čamdžija Z.F. Complete study of nature and importance of spectral bands contained in infrared spectra of leaves of maize inbred lines with significant breeding properties. Russian Agricultural Sciences, 2019, 45(4): 334-339 (doi: 10.3103/S106836741904013X).
- Vollhardt P.C., Schore N.E. Organic сhemistry. W.H. Freeman and Company, United States, 1996.
- Krimm S., Bandekar J. Vibrational spectroscopy and conformation of peptides, polypeptides, and proteins. Advances in Protein Chemistry, 1986, 38: 181-364 (doi: 10.1016/S0065-3233(08)60528-8).
- Свердлов Л.М., Ковнер М.А., Крайнов Е.П. Колебательные спектры многоатомных молекул. М., 1970.
- Merlin J. Resonance Raman spectroscopy of carotenoids and carotenoid-containing systems. Pure and Applied Chemistry, 1985, 57(5): 785-792 (doi: 10.1351/pac198557050785).
- Amir R.M., Anjum F.M., Khan M.I., Khan M.R., Pasha I., Nadeem M. Application of Fourier transform infrared (FTIR) spectroscopy for the identification of wheat. J. FoodSci. Technol., 2013, 50: 1018-1023 (doi: 10.1007/s13197-011-0424-y).
- Тютяев Е.В., Шутова В.В., Максимов Г.В., Раденович Ч., Гродзинский Д.М. Состояние фотосинтетических пигментов в листьях инбредных линий и гибридов кукурузы. Физиология растений и генетика, 2015, 47(2): 147-159.
- Berthold D.A., Babcock G.T., Yocum C. F. A highly resolved, oxygen evolving Photosystem II preparation from spinach thylakoid membranes. FEBS Lett., 1981, 134(2): 231-234 (doi: 10.1016/0014-5793(81)80608-4).
- Ловягина Е.Р., Давлетшина Л.Н., Култышева М.Ю., Тимофеев К.Н., Иванов И.И., Семин Б.К. Особенности процесса взаимодействия катионов Fe(II) с донорной стороной фотосистемы II, не содержащей марганец. Физиология растений, 2005, 52(1): 12-20.
- Shutova V., Yusipovich A., Parshina E., Zakharkin D., Revin V. Effect of particle size on the enzymatic hydrolysis of polysaccharides from ultrafine lignocellulose particles. Applied Biochemistry and Microbiology, 2012, 48(3): 312-317 (doi: 10.1134/S000368381203012X).
- Radenović Č., Grodzinskij D.M., Filipović M. The prestigious maize inbred lines and hybrids with erect top leaves are characterised by a property of an efficient photosynthetic model and a satisfactory base for the further progress in breeding and selection. Физиология и биохимия культурныхрастений, 2010, 42(3): 187-201.
- Shutova V.V., Tyutyaev E.V., Churin A.A., Ponomarev V.Yu, Belyakova G.A., Maksimov G.V. IR and Raman Spectroscopy in the study of carotenoids of Cladophora rivularis algae. Biophysics, 2016, 61(4): 601-605 (doi: 10.1134/S0006350916040217).
- Robert B. The electronic structure, stereochemistry and resonance Raman spectroscopy of carotenoids. In: The photochemistry of carotenoids. New York, Boston, Dordrecht, London, Moscow, Kluwer Academic Publishers, 2004: 189-201.
- Hudson S.D., Chumanov G. Bioanalytical applications of SERS (surface-enhanced Raman spectroscopy). Analytical and Bioanalytical Chemistry, 2009, 394(3): 679-686 (doi: 10.1007/s00216-009-2756-2).
- Ruban A.V., Berera R., Ilioaia C. van Stokkum IH, Kennis JT, Pascal AA, van Amerongen H, Robert B, Horton P, van Grondelle R. Identification of a mechanism of photoprotective energy dissipation in higher plants. Nature, 2007, 450: 575-578 (doi: 10.1038/nature06262).
- Gruszecki W.I., Gospodarek M., Grudzinski W. Mazur R, Gieczewska K, Garstka M. Light-induced change of configuration of the LHCII-Bound xanthophyll (tentatively assigned to violaxanthin): a resonance Raman study. J. Phys. Chem. B, 2009, 113: 2506-2512 (doi: 10.1021/jp8101755).
- Todorenko D.A., Hao J., Slatinskaya O.V., Allakhverdiev E.S., Khabatova V.V., Ivanov A.D., Radenovic C.N., Matorin D.N., Alwasel S., Maksimov G.V., Allakhverdiev S.I. Effect of thiamethoxam on photosynthetic pigments and primary photosynthetic reactions in two maize genotypes (Zea mays). Functional Plant Biology, 2021, 48(10): 994-1004 (doi: 10.1071/FP21134).
- Radenović Č., Maksimov G.V., Grodzinskij M.D. Identification of organic molecules in kernels of maize inbred lines displayed with infrared spectra. Plant Physiology and Genetics (Kiev, Ukraine), 2015, 47(1/273): 15-24.
- Radenović, Č., Filipović M., Selaković D. The prestigious maize inbred lines with erect top leaves. The priority performance of the efficient photosynthetic model in breeding. Genetika, 2009, 41(1): 49-58 (doi: 10.2298/GENSR0901041R).