doi: 10.15389/agrobiology.2021.5.924rus
УДК 615.32:576.3/.7.086.83
Работа финансировалась Министерством науки и технологий Вьетнама в рамках проекта «Оптимизация протокола микроразмножения женьшеня Ngoc Linh (Panax vietnamensis) с использованием биореакторной системы культивирования» (2018-2019).
ВЫСОКОЭФФЕКТИВНОЕ МИКРОРАЗМНОЖЕНИЕ ВЬЕТНАМСКОГО
ЖЕНЬШЕНЯ Panax vietnamensis Ha et Grushv. С ИСПОЛЬЗОВАНИЕМ
КУЛЬТУРЫ in vitro НЕЗРЕЛЫХ ЗИГОТИЧЕСКИХ ЗАРОДЫШЕЙ
T.X. DINH1, M.L. NGUYEN2 ✉, D.D. VU1, G.T. HOANG3 ✉
Несмотря на значительную экономическую выгоду при выращивании вьетнамского женьшеня (Panax vietnamensis Ha et Grushv.), или женьшеня Ngoc Linh, в качестве лекарственного растения, масштабы его культивирования не расширяются из-за ограниченного количества посадочного материала. Предприняты значительные исследовательские усилия по разработке систем культуры тканей у вьетнамского женьшеня, однако его микроразмножение все еще сопряжено с трудностями. Нами предложен протокол микроразмножения вьетнамского женьшеня посредством соматического эмбриогенеза in vitro в культуре незрелых зиготических зародышей. Частота прямого соматического эмбриогенеза из незрелых зиготических зародышей на основной среде Schenk и Hildebrandt (SH) с 7 % сахарозы составила 44,33 %. Вторичный соматический эмбриогенез с коэффициентом пролиферации 26,71 индуцировали в жидкой культуре с встряхиванием в течение 4 нед в среде 1/2 SH с кинетином (0,1 мг/л) и тидиазуроном (0,1 мг/л). Среда SH с добавлением α-нафтилуксусной кислоты (НУК, 1 мг/л), 6-бензиладенина (6-БАП, 0,5 мг/л) и гибберелловой кислоты (5 мг/л) стимулировала прорастание соматических зародышей и раннее образование микроризомов. Далее у двух 10-недельных культур на среде SH с НУК (0,5 мг/л), 6-БАП (1 мг/л) и 4 % сахарозы в результате микроразмножения были получены проростки с хорошо развитой корневой системой и утолщенным микроризомом со спящей почкой. Акклиматизация микроразмноженных проростков, высаженных почву, состоящую из смеси лесного перегноя с песком в соотношении 1:2, прошла успешно с выживаемостью 91,67 %. Таким образом, для женьшеня Ngoc Linh разработан эффективный протокол микроразмножения на основе культуры in vitro незрелых зиготических зародышей, который состоит из пяти этапов, включая индукцию эмбриогенеза, пролиферацию соматических зародышей, прорастание соматических зародышей, развитие проростков и акклиматизацию проростков.
Ключевые слова: Panax vietnamensis Ha et Grushv., вьетнамский женьшень, женьшень Нгок Линь, культура in vitro, зиготический зародыш, соматический эмбриогенез, прорастание соматических зародышей, микроризом, микроразмножение.
Вьетнамский женьшень, или женьшень Ngoc Linh (Panax vietnamensis Ha et Grushv.), принадлежащий к семейству Araliaceae, — ценное эндемичное лекарственное растение. В природе вьетнамский женьшень растет только на высоте более 1200 м над уровнем моря и под пологом влажных тропических лесов с дневной температурой от 20 до 25 °С и ночью от 15 до 18 °С. Вьетнамский женьшень — медленнорастущее и долгоживущее травянистое многолетнее растение высотой от 40 до 100 см. На его корневищах ежегодно образуются рубцы от опавших надземных стеблей. Корневище вьетнамского женьшеня содержит 52 сапонина, 24 из которых не встречаются у других видов женьшеня (1). Содержание сапонинов в корневище женьшеня Ngoc Linh выше, чем у других видов рода Panax L. (2, 3).
Несмотря на высокую экономическую ценность вьетнамского женьшеня, его выращивание в крупных масштабах пока затруднено. С 1983 года многие исследования были направлены на разработку технологий выращивания и применения женьшеня Ngoc Linh в качестве лекарственного растения (4).
Данные об успешном микроразмножении корейского женьшеня (Panax ginseng C.A. Meyer) и других видов женьшеня стимулировали исследования культуры тканей женьшеня Ngoc Linh, в частности изучение органогенеза (5), формирования искусственных семян (6), образования микроризомов (7), соматического эмбриогенеза (8-12). Некоторые работы были сосредоточены на культурах вторичных и придаточных корней (13) для производства биомассы женьшеня Ngoc Linh с целью получения гинсенозидов (14-18). Соматический эмбриогенез считается наиболее изученным методом микроразмножения применительно к женьшеню Ngoc Linh из-за большого количества продуцируемых соматических зародышей и высокой частоты регенерации растений. M. Truong с соавт. (11, 12) сообщили о потенциале соматического эмбриогенеза и морфогенеза побегов или корней из клеточной суспензионной культуры каллусов, полученных из листовых эксплантатов. В исследовании D. Nhut с соавт. (9), соматический эмбриогенез был индуцирован в культуре in vitro на основе тонкого слоя клеток основного корня. Однако, выживаемость проростков после переноса в питомник оказалась довольно низкой, поэтому считается, что методы in vitro все еще не обеспечивают стабильного размножения женьшеня Ngoc Linh.
На сегодняшний день не сообщалось об исследованиях по использованию незрелых семян женьшеня Ngoc Linh в качестве источника эксплантатов для получения незрелых зиготических зародышей (НЗЗ) и их последующего культивирования in vitro.
В этом исследовании мы оптимизировали протокол для микроразмножения женьшеня Ngoc Linh, используя соматический эмбриогенез в культуре НЗЗ. Этот протокол состоит из пяти этапов, включая индукцию соматического эмбриогенеза, пролиферацию соматических зародышей, прорастание соматических зародышей, развитие проростков и акклиматизацию проростков. Протокол будет полезен для крупномасштабного распространения женьшеня Ngoc Linh.
Методика. Незрелые семена женьшеня Ngoc Linh собирали из свежих плодов, собранных на экспериментальном поле в горах Ngoc Linh (14°58´34´´N и 107°54´41´´E). Семена извлекали из капсул, стерилизовали 1 % NaClO в течение 15 мин, затем обрабатывали раствором стрептомицина (500 мг/л) в течение 10 мин и кратковременно погружали в 70 % этанол. Далее семена тщательно промывали стерильной дистиллированной водой. После стерилизации удаляли оболочку семян и 2/3 энтодермы и собирали НЗЗ, которые использовали в качестве эксплантов для культивирования in vitro.
Собранные НЗЗ инкубировали в культуральной среде для индукции первичных соматических зародышей (СЗ). Основные среды Schenk-Hil-debrandt (SH) (19) и Murashige-Skoog (MS) (20) с разной (полной, половинной) концентрацией солей с добавлением 7 % сахарозы использовали отдельно или в сочетании с одним из регуляторов роста растений — 2,4-дихлорфеноксиуксусной кислотой (2,4-D, 0,2 и 0,5 мг/л), тидиазуроном (ТДЗ, 0,2 и 0,5 мг/л), α-нафтилуксусной кислотой (НУК, 1 мг/л) и индолил-3-масляной кислотой (ИМК, 3 мг/л). Долю эксплантов, индуцировавших первичные СЗ, оценивали через 10 нед культивирования.
Образовавшиеся первичные СЗ отделяли от эмбриогенных тканевых масс. Для этого эмбриогенные тканевые массы переносили в жидкую среду SH половинной концентрации с добавлением 3 % сахарозы в соотношении от 1/5 до 1/10 (масса/объем). Разделенные первичные СЗ использовали для пролиферации вторичных СЗ. Исследовали влияние двух типов питательных сред (жидких и плотных) на основе SH половинной концентрации с 3 % сахарозы и различными комбинациями кинетина (0,1 мг/л) и ТДЗ (0; 0,05; 0,1 и 0,2 мг/л). Содержание агара в плотной среде — 7 г/л. В жидкой среде экспланты (соотношение эксплант:среда 1:20) культивировали на шейкере (100 об/мин). Коэффициент пролиферации оценивали через 4 нед культивирования и рассчитывали, как отношение сырой массы вторичных СЗ через 4 нед культивирования к сырой массе инокулированных первичных СЗ.
Для прорастания вторичных СЗ использовали плотную среду SH с добавлением 1 мг/л НУК и 0,5 мг/л 6-БАП в сочетании с разными концентрациями гибберелловой кислоты (ГК3) (0, 1, 3, 5 и 7 мг/л). После 10-недельного культивирования регистрировали число проросших СЗ для расчета частоты прорастания.
После прорастания полученные небольшие проростки с корнями и побегами переносили на среду SH с 0,5 мг/л НУК, 1 мг/л 6-БАП и разными концентрациями сахарозы (2, 3, 4, 5 и 6 %) для индукции микроризомов. Через 10 нед проростки переносили на ту же среду еще на 10 нед для инициации спящих почек на микроризомах. Через 5 мес культивирования измеряли биомассу растений и длину побегов. Также учитывали наличие спящих почек.
Хорошо развитые всходы с микроризомами и спящими почками переносили на экспериментальное поле на высоту 1800 м. Был проведен двухэтапный эксперимент. Перед пересадкой проростки помещали в 3 % раствор хитозана на 10 мин. После этого их высаживали в полиэтиленовые пакеты, содержащие различные типы почвенных смесей: 1) лесной перегной, 2) 2/3 лесного перегноя + 1/3 песка, 3) 1/3 лесного перегноя + 2/3 песка, 4) 1/5 лесного перегноя + 4/5 песка, 5) 1/3 лесного перегноя + 1/3 кокосового волокна + 1/3 перлита. Через 3 нед после посадки подсчитывали процент проросших проростков, через 3 мес оценивали выживаемость и ростовые параметры растений.
Все эксперименты проводили в 3 повторностях с 20 эксплантами для каждой обработки. Все культуры поддерживали в культуральной комнате при 23 °С при 14-часовом фотопериоде. Для сравнения результатов был применен критерий Тьюки (Tukey’s multiple range test) с использованием Statgraphics Centurion XVI (https://www.statgraphics.com/download-statgraphics-centurion-xvi).
Результаты. Мы выявили влияние различных основных сред и различных регуляторов роста растений на индукцию СЗ. Инициация соматического эмбриогенеза в культуре in vitro НЗЗ на всех исследованных средах начиналась после 5 нед культивирования (рис. 1, A). После 10 нед культивирования из 10 исследованных вариантов сред среда SH с 7 % сахарозы обеспечивала максимальную скорость инициации СЗ (44,33 %), которая была значительно выше, чем в других средах (1/2 MS, 1/2 SH и MS) и средах с регуляторами роста растений (2,4-D, ТДЗ, НУК и ИМК). После 10 нед культивирования на среде SH с 7 % сахарозой мы наблюдали образование соматических зародышей на нескольких стадиях развития (см. рис. 1, Б-Г). Кроме того, также на средах MS с высокой частотой появлялись аномальные соматические зародыши.
После кратковременного встряхивания эмбриогенных тканевых масс в жидкой среде СЗ надежно отделялись от родительских эксплантатов, которые затем удаляли (см. рис. 1, Д). Для пролиферации полученных СЗ тестировали плотную и жидкую среду 1/2 SH с добавлением 0,1 мг/л кинетина и разных концентраций ТДЗ. После 3 нед культивирования вокруг основания первичных СЗ начали формироваться новые эмбрионы, образовавшие затем кластеры (по 4-12 в каждом), которые наблюдали на разных стадиях развития. Индуцированные эмбрионы оставались компактными и светло-желтыми в течение 4 нед культивирования (см. рис. 1, Е, Ж) и становились зеленовато-желтыми в том случае, если культивирование продолжали. Наименьшую эффективность вторичного эмбриогенеза отмечали на среде 1/2 SH с добавлением кинетина (0,1 мг/л) как в плотных, так и в жидких культурах (коэффициент пролиферации соответственно 8,63 и 8,27). На плотной среде коэффициент пролиферации СЗ постепенно увеличивался с 8,63 до 13,9 в присутствии ТДЗ и с увеличением его концентрации (особенно при 0,2 мг/л ТДЗ). Интересно, что вторичный эмбриогенез значительно улучшался в жидкой культуре со встряхиванием по сравнению с культурой на плотной среде того же состава. Наибольший коэффициент пролиферации — 26,71 отмечали в жидкой среде 1/2 SH с 0,1 мг/л кинетина и 0,1 мг/л ТДЗ.
На среде без ГК3 (рис. 2) прорастание СЗ происходило с очень низкой частотой (6,33 %). При обработке ГК3 соматические зародыши быстро прорастали с увеличением частоты до 72,28 % на среде с 5 мг/л ГК3. Однако добавление более высокой концентрации ГК3 (7 мг/л) снизило частоту прорастания СЗ на 28,67 %.
Полученные результаты показали существенное влияние сахарозы на рост проростков и утолщение микроризомов (табл. 1). Несмотря на то, что на среде с 2 % сахарозы, микроризомы были утолщены, спящие почки не образовывались. При повышении концентрации сахарозы отмечали интенсивный рост проростков, растения были выше и накапливали бóльшую биомассу. Тем не менее мы отмечали, что на средах с концентрацией сахарозы 5 и 6 % края листьев становились сухими и беловатыми после трех первых суток культивирования. В этих вариантах опыта растения далее росли медленно; через 10 нед культивирования происходила индукция соматических зародышей на микроризомах. Оптимальная концентрация сахарозы для роста проростков составила 4 %, при этом примерно после 5 мес культивирования растения в среднем достигали высоты 7,44 см при массе 1,60 г, имели видимый микроризом (длиной около 5 мм) и спящую почку.
Через 3 нед после посадки спящие почки начали прорастать с частотой от 10,00 до 61,67 % в зависимости от типа почвенной смеси (табл. 2).
В течение следующих недель побеги продолжали формироваться, что служило индикатором для прогнозирования выживаемости микроклональных проростков женьшеня. В результате выживаемость, оцененная через 3 мес после посадки, была прямо пропорциональна проценту побегообразования через 3 нед после посадки. Мы обнаружили, что испытанные почвенные смеси влияют на выживаемость и рост проростков, полученных методом микроразмножения. После высадки в лесной перегной процент проростков, образующих побеги, через 3 нед после посадки был довольно низким (10 %), через 3 мес доля выживших проростков достигла 71,67 % при среднем числе корней 0,92 со средней длиной 0,49 см. Число корней было выше в почвенных смесях из 2/3 лесного перегноя + 1/3 кокосового волокна + 1/3 перлита (2,00 корня), из 1/5 лесного перегноя + 4/5 песка (2,52 корня) и из 2/3 лесного перегноя + 1/3 песка (2,58 корня). Мак-симальный рост корней (5,05 корня при средней длине 1,9 см), а также максимальную выживаемость (91,67 %) регистрировали в почвенной смеси, содержащей 1/3 лесного перегноя + 2/3 песка (рис. 3, Д, Е).
Настоящее сообщение представляет собой первое исследование культуры тканей на основе НЗЗ в качестве первичных эксплантатов для получения СЗ у женьшеня Ngoc Linh. В этом исследовании НЗЗ не были полностью препарированы от эндодермы, а отделены вместе с ее часть, потому что НЗЗ у женьшеня Ngoc Linh (и у женьшеней вообще) крошечные (21, 22) и легко повреждаются при отсечении от эндодермы. После 10 нед культивирования лучший результат для прямого соматического эмбриогенеза был получен на среде SH с 7 % сахарозой без каких-либо регуляторов роста растений. Сообщалось об использовании высокой концентрации сахарозы (5 %) для индукции СЗ у корейского женьшеня (результаты были получены на семядолях из незрелых или зрелых зиготических зародышей) (21-23). Как и в работах других авторов (24, 25), в нашем исследовании добавление в среду регуляторов роста растений привело к непрямому соматическому эмбриогенезу. Интересно, что самая высокая частота эксплантов (44,33 %), образующих СЗ, в этом исследовании была зарегистрирована на основной среде SH, а не на среде MS, как сообщалось (22). Более того, основная среда SH индуцировала образование рыхлой эмбриогенной массы, на которой появлялись одиночные соматические зародыши, в то время как большое количество соматических зародышей, появившихся на основной среде MS, были аномальными или не разделялись, что приводило к образованию дефектных проростков. Многие исследователи также сообщили о высокой частоте образования аномальных соматических эмбрионов в культуре тканей женьшеня (23, 24, 26-28). При размножении растений in vitro важно получать морфологически нормальные растения (в частности, в нашем эксперименте — из одиночных соматических зародышей). Мы собрали отделенные друг от друга первичные СЗ из сформированной эмбриогенной массы до пролиферации вторичных СЗ. Индивидуальные первичные СЗ легко отделялись от родительских эксплантатов при кратковременном встряхивании в жидкой культуре. Типы культур и регуляторы роста растений влияли на коэффициент пролиферации СЗ, который был выше в жидкой культуре на среде 1/2 SH с добавлением 0,1 мг/л кинетина и 0,1 мг/л ТДЗ. После 4 нед культивирования первичные СЗ генерировали множество новых эмбрионов, что рассматривается как вторичный эмбриогенез (см. рис. 1, Д-Ж). Поскольку формирование новых эмбрионов происходило асинхронно, эмбрионы, полученные после 4 нед культивирования, находились на разных стадиях развития — от глобулярных до семядольных.
Частота прорастания СЗ была намного выше на среде с добавлением ГК3, чем на среде без ГК3. Мы наблюдали, что на среде без ГК3 продолжается процесс вторичного соматического эмбриогенеза, что приводит к ингибированию развития эмбрионов. На средах с добавлением ГК3 развитие зародыша происходило нормально и включало глобулярную, сердцевидную, биполярную и семядольную стадии, затем происходило прорастание с образованием проростков (см. рис. 3, А, Б). Наилучший результат, который дал частоту прорастания 72,28 %, был получен при добавлении 5 мг/л ГК3 (см. рис. 3, В), как описано и J.Y. Zhang с соавт. (29). Следовательно, ГК3 стимулировала прорастание соматических зародышей у женьшеня Ngoc Linh. Это согласуется с результатами для других видов женьшеня (23, 28, 30, 31). После 10 нед культивирования на питательной среде для проращивания были получены проростки с высотой побегов 2-3 см и микроризомами диаметром 3 мм (см. рис. 3, Г). Это подтвердило эффективность использованной среды, в которую добавляли НУК и 6-БАП с целью стимулировать раннюю индукцию микроризомов.
Затем рост проростков и микроризомов усиливался при выращивании на средах с различными концентрациями сахарозы (от 2 до 6 %) в течение примерно 5 мес. Сообщалось о влиянии сахарозы на рост проростков и утолщение корней у женьшеня обыкновенного Panax ginseng (32). В настоящем исследовании наибольший успех был достигнут при использовании среды с 4 % сахарозы. Примерно через 5 мес в культурах с добавлением 3-6 % сахарозы образовывались спящие почки. Наличие спящей почки играет важную роль в выживании полученных in vitro проростков женьшеня при переносе в почву (32). Объяснение может заключаться в том, что в естественных условиях корни у растений женьшеня начинают расти после появления первого побега.
Перед посадкой в почву проростки обрабатывали 3 % хитозаном для борьбы с болезнями (33-35). В нашем исследовании без обработки хитозаном доля микроклональных проростков, пораженных корневой гнилью, достигала 90 % (данные не представлены), поэтому перед пересадкой проростки обрабатывали хитозаном. Мы также обнаружили влияние почвенных смесей на акклиматизацию проростков in vitro. Смесь лесного перегноя с песком в соотношении 1:2 значительно превосходила другие варианты из-за текстуры, подходящей для посадки рассады, полученной in vitro. Лесной перегной обогащен питательными веществами, а песок обеспечивает достаточную аэрацию и дренаж. На этой почвенной смеси выживаемость микроклональных проростков достигла 91,67 %. У растений старые листья пожелтели и опали через 5-6 нед после посадки, но растения хорошо росли, форируя новые листья и корни (см. рис. 3, Д, Е).
Итак, мы оптимизировали протокол для микроразмножения женьшеня Ngoc Linh, используя соматический эмбриогенез в культуре in vitro незрелых зиготических зародышей. Протокол включает индукцию соматического эмбриогенеза, пролиферацию соматических зародышей, прораста-ние соматических зародышей, развитие проростков и акклиматизацию проростков. Частота прямого соматического эмбриогенеза на среде Schenk и Hildebrandt (SH) с 7 % сахарозы составила 44,33 %. Вторичный соматический эмбриогенез с коэффициентом пролиферации 26,71 индуцировали в жидкой культуре в среде 1/2 SH с кинетином (0,1 мг/л) и тидиазуроном (0,1 мг/л). Среда SH с α-нафтилуксусной кислотой (НУК, 1 мг/л), 6-бензиладенином (6-БАП, 0,5 мг/л) и гибберелловой кислотой (ГК3, 5 мг/л) стимулировала прорастание соматических зародышей и раннее образование микроризомов. В результате микроразмножения на среде SH с НУК (0,5 мг/л), 6-БАП (1 мг/л) и 4 % сахарозы были получены проростки с хорошо развитой корневой системой и утолщенным микроризомом со спящей почкой. При акклиматизации микроразмноженных проростков на смеси лесного перегноя с песком в соотношении 1:2 выживаемость составила 91,67 %. Разработанный нами протокол позволяет ускорить процесс микроразмножения, а также значительно улучшает коэффициент пролиферации и выживаемость растений в полевых условиях. Его можно применять для крупномасштабного выращивания женьшеня Ngoc Linh.
1. Рост проростков женьшеня Ngoc Linh (Panax vietnamensis Ha et Grushv.), полученных в культуре in vitro, |
||||
Среда |
Концентрация сахарозы, % |
Биомасса растения, г |
Высота растения, см |
Наличие спящей почки |
SH + 0,5 мг/л НУК + 1 мг/л 6-БАП |
2 |
0,58±0,01a |
4,35±0,03a |
Нет |
SH + 0,5 мг/л НУК + 1 мг/л 6-БАП |
3 |
0,80±0,01b |
6,12±0,06b |
Да |
SH + 0,5 мг/л НУК + 1 мг/л 6-БАП |
4 |
1,60±0,02c |
7,44±0,09c |
Да |
SH + 0,5 мг/л НУК + 1 мг/л 6-БАП |
5 |
1,14±0,01d |
6,24±0,06b |
Да |
SH + 0,5 мг/л НУК + 1 мг/л 6-БАП |
6 |
1,33±0,00e |
5,62±0,05d |
Да |
Примечание. SH — среда Schenk-Hildebrandt, НУК — α-нафтилуксусная кислота, 6-БАП — 6-бензил-аденин. |
||||
2. Акклиматизация проростков женьшеня Ngoc Linh (Panax vietnamensis Ha et Grushv.), полученных методом |
||||
Состав |
Через 3 нед после посадки |
Через 3 мес после посадки |
||
доля проростков с побегами, % |
корни |
выживаемость, % |
||
число |
длина, см |
|||
Лесной перегной |
10,00±4,08a |
0,92±0,12a |
0,49±0,00a |
71,67±1,67a |
2/3 лесного перегноя + 1/3 песка |
38,33±6,39bc |
2,58±0,14b |
1,04±0,01b |
73,33±1,67a |
1/3 лесного перегноя + 2/3 песка |
61,67±7,26c |
5,05±0,20c |
1,90±0,03c |
91,67±1,67b |
1/5 лесного перегноя + 4/5 песка |
36,67±8,04abc |
2,52±0,12bd |
0,88±0,01d |
80,00±5,77ab |
1/3 лесного перегноя + 1/3 кокосового волокна + 1/3 перлита |
20,00±6,07ab |
2,00±0,10d |
0,55±0,01e |
70,00±2,89a |
abcd Различия между значениями, отмеченными разными буквами, статистически значимы по критерию Тьюки (Tukey’s multiple range test) при р < 0,05. |
||||

Рис. 1. Прямой соматический эмбриогенез в культуре in vitro незрелых зиготических зародышей (А-Г) и пролиферация вторичных соматических зародышей (Д-Ж) у женьшеня Ngoc Linh (Panax vietnamensis Ha et Grushv.): A — соматический эмбриогенез после 5 нед культивирования; Б — соматический эмбриогенез после 8 нед культивирования; В — соматический эмбриогенез после 10 нед культивирования; Г — первичные соматические зародыши на глобулярной и биполярной стадиях; Д — после изоляции первичных соматических зародышей; Е — после 2 нед пролиферации соматических эмбрионов; Ж — после 4 нед пролиферации соматических эмбрионов.
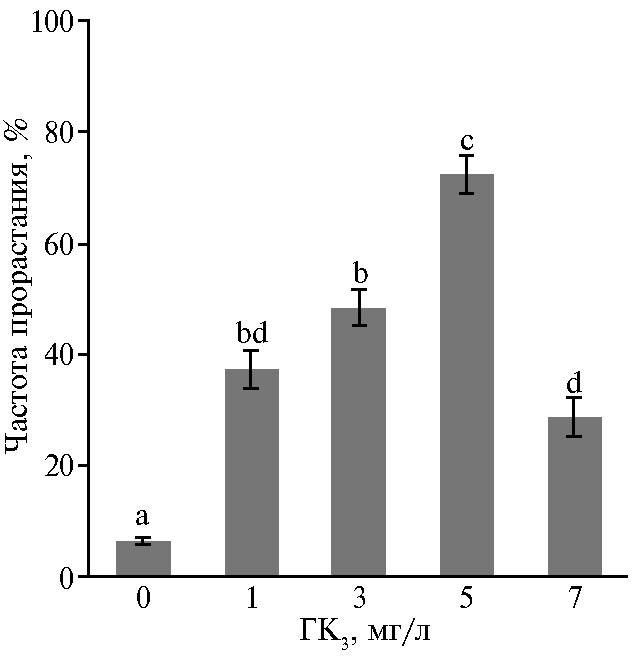
Рис. 2. Прорастание соматических зародышей женьшеня Ngoc Linh (Panax vietnamensis Ha et Grushv.), полученных в культуре in vitro незрелых зиготических зародышей, при разной концентрации гибберелловой кислоты (ГК3) в среде. Различия между значениями, отмеченными разными буквами, статистически значимы по критерию Тьюки (Tukey’s multiple range test) при р < 0,05.

Рис. 3. Прорастание соматических зародышей и развитие проростков женьшеня Ngoc Linh (Panax vietnamensis Ha et Grushv.) при микроразмножении: A — прорастающие соматические зародыши; Б — через 4 нед культивирования на среде для проращивания; В — через 10 нед культивирования на среде для проращивания; Г — растение с утолщенным микроризомом после культивирования на среде для проращивания; Д — растения через 8 нед после посадки; Е — растения через 3 мес после посадки.
ЛИТЕРАТУРА
- Nguyen M.D., Kasai R., Yamasaki K., Nham N.T., Tanaka O. New dammarane saponins from Vietnamese ginseng. Studies in Plant Science, 1999, 6: 77-82 (doi: 10.1016/S0928-3420(99)80010-X).
- Nguyen M., Nguyen T., Kasai R., Ito A., Yamasaki K., Tanaka O. Saponins from Vietnamese ginseng, Panax vietnamensis Ha et Grushv. collected in central Vietnam. I. Chem. Pharm. Bull. (Tokyo), 1993, 41(11): 2010-2014 (doi: 10.1248/cpb.41.2010).
- Yamasaki K. Bioactive saponins in Vietnamese ginseng, Panax vietnamensis. Pharmaceutical Biology, 2000, 38: 16-24 (doi: 10.1076/phbi.38.6.16.5956).
- Tran B.T., Nguyen T.H., Nguyen T.T.M., Truong T.C., Pham T.H., Pham H.S. Evaluation of growth and bioactive compositions of Vietnamese ginseng (Panax vietnamensis) cultivated in Quang Nam. VNU Journal of Science: Natural Sciences and Technology, 2017, 33(2S): 227-232 (doi: 10.25073/2588-1140/vnunst.4594).
- Nhut D., Nga L., Chien H., Huy N. Morphogenesis of in vitro main root transverse thin cell layers of Vietnamese ginseng (Panax vietnamensis Ha et Grushv.). African Journal of Biotechnology, 2012, 11: 6274-6289 (doi: 10.5897/AJB11.2884).
- Trinh T.H., Duong T.N. In vitro germination somatic embryo-derived artificial seed of Ngoc Linh ginseng (Panax vietnamensis Ha et Grushv.). Vietnam Journal of Biotechnology, 2011, 9: 443-453.
- Hoang X.C., Ngo T.T., Nguyen B.T., Tran X.T., Lam B.T., Tran C.L. Effect of some factors to in vitro microrhizome formation (Panax vietnamensis Ha et Grushv.) and determination of plantlet saponin content in Ngoc Linh mountain. Vietnam Journal of Biotechnology, 2011, 9: 317-331.
- Vu T.H., Vu Q.L., Nguyen P.H., Nguyen B.N., Bui V.T.V., Thai X.D., Duong T.N. Direct somatic embryogenesis from leaf, petiole and rhizome explant of Panax vietnamensis Ha et Grushv. Academia Journal of Biology, 2014, 36: 277-282 (doi: 10.15625/0866-7160/v36n1se.4408).
- Nhut D., Vinh B., Hien T., Huy N., Nam N., Chien H. Effects of spermidine, proline and carbohydrate sources on somatic embryogenesis from main root transverse thin cell layers of Vietnamese ginseng (Panax vietnamensis Ha et. Grushv.). African Journal of Biotechnology, 2012, 11: 1084-1091 (doi: 10.5897/AJB11.3186).
- Trieu N.B., Tung N.T., Phuong T.T.B. In vitro culture of Ngoc Linh ginseng (Panax vietnamensis Ha et Grushv.). Hue University Journal of Science: Agriculture and Rural Development, 2013, 79: 107-115 (doi: 10.26459/jard.v79i1.3128).
- Truong M., Tran T.N.H., Phan T.L., Le T.D., Tran T.T., Do D.G., Bui D.T., Pham D.T., Nguyen D.M.H., Nguyen T.T., Nguyen V.K., Tran C.L., Nguyen H.H. Studies on induction and multiplication of somatic embryos of Ngoc Linh ginseng (Panax vietnamensis Ha et Grushv.) in liquid medium. Journal of Science and Development, 2014, 12(5): 1085-1095.
- Truong M., Tran T.N.H., Phan T.L., Le T.D., Tran T.T., Do D.G., Bui D.T., Pham D.T., Nguyen D.M.H., Nguyen T.T., Nguyen V.K., Tran C.L., Nguyen H.H. The study on in vitro culture of embryogenic calllus and somatic embryo tissue of Panax vietnamensis Ha et Grushv. Academia Journal of Biology, 2013, 35: 145-157 (doi: 10.15625/0866-7160/v35n3se.3855).
- Lieu N.T., Thanh N.T., Ket N.V. The adventitious root induce of Ngoc Linh ginseng (Panax vietnamensis, Ha et Grushv.) in vitro cultures. VNU Journal of Science: Natural Sciences and Technology, 2011, 27: 30-36.
- Huong T.T., Ngoc P.B., Ha C.H., Nhut D.T. Assessing the growth and saponin accumulation in hairy and adventitious roots of Vietnamese ginseng (Panax vietnamensis Ha et Grushv.). Vietnam Journal of Biotechnology, 2016, 14(2): 231-236 (doi: 10.15625/1811-4989/14/2/9335).
- Nhut D.T., Luan V.Q., Binh N.V., Phong P.T., Huy B.N., Ngoc Ha D.T., et al. The effects of some factors on in vitro biomass production of Vietnamese ginseng (Panax vietnamensis Ha et Grushv.) and preliminary analysis of saponin content. Vietnam Journal of Biotechnology, 2009, 7(3): 357-370 (doi: 10.15625/1811-4989/7/3/12453).
- Thanh N.T., Anh H.T., Yoeup P.K. Effects of macro elements on biomass and ginsenoside production in cell suspension culture of Ngoc Linh ginseng (Panax vietnamensis Ha et Grushv.). VNU Journal of Science: Natural Sciences and Technology, 2008, 24(3): 248-252.
- Thanh N.T., Ket N.V., Yoeup P.K. Effecting of medium composition on biomass and ginsenoside production in cell suspension culture of Panax vietnamensis Ha et Grushv. VNU Journal of Science, Natural Sciences and Technology, 2007, 23(4): 269-274.
- Trong T.T., Truong D.-H., Nguyen H.C., Tran D.-T., Nguyen Thi H.-T., Dang G.D., Huu H.N. Biomass accumulation of Panax vietnamensis in cell suspension cultures varies with addition of plant growth regulators and organic additives. Asian Pacific Journal of Tropical Medicine, 2017, 10(9): 907-915 (doi: 10.1016/j.apjtm.2017.08.012).
- Schenk R., Hildebrandt A. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell cultures. Canadian Journal of Botany, 1972, 50(1): 199-204 (doi: 10.1139/b72-026).
- Murashige T., Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum, 1962, 15(3): 473-497 (doi: 10.1111/j.1399-3054.1962.tb08052.x).
- Choi K.T., Choi Y.E., Yang D.C., Park J.C. Somatic embryogenesis in Korean ginseng (Panax ginseng C.A. Meyer). In: Plant biotechnology and in vitro biology in the 21st century, vol. 36 /A. Altman, M. Ziv, S. Izhar (eds.). Springer Netherlands, Dordrecht, 1999: 29-32 (doi: 10.1007/978-94-011-4661-6_6).
- Kim O.T., Kim T.S., In D.S., Bang K.H., Kim Y.C., Choi Y.E., Cha S.W., Seong N.S. Optimization of direct somatic embryogenesis from mature zygotic embryos of Panax ginseng C.A. Meyer. Journal of Plant Biology, 2006, 49: 348-552 (doi: 10.1007/BF03178810).
- Choi Y.E., Yang D.C., Park J.C., Soh W.Y., Choi K.T. Regenerative ability of somatic single and multiple embryos from cotyledons of Korean ginseng on hormone-free medium. Plant Cell Reports, 1998, 17(6-7): 544-551 (doi: 10.1007/s002990050439).
- Arya S., Arya I.D., Eriksson T. Rapid multiplication of adventitious somatic embryos of Panax ginseng. Plant Cell, Tissue and Organ Culture, 1993, 34: 157-162 (doi: 10.1007/BF00036096).
- Kim Y.J., Lee O.R., Kim K.T., Yang D.C. High frequency of plant regeneration through cyclic secondary somatic embryogenesis in Panax ginseng. Journal of Ginseng Research, 2012, 36(4): 442-448 (doi: 10.5142/jgr.2012.36.4.442).
- Chang W., Hsing Y. Plant regeneration through somatic embryogenesis in root-derived callus of ginseng (Panax ginseng C.A. Meyer). Theoretical and Applied Genetics, 1980, 57(3): 133-135 (doi: 10.1007/BF00253888).
- Lee H.S., Liu J.R., Yang S.G., Lee Y.H., Lee K.W. In vitro flowering of plantlets regenerated from zygotic embryo-derived somatic embryos of ginseng. HortScience, 1990, 25(12): 1652-1654 (doi: 10.21273/HORTSCI.25.12.1652).
- Zhou S., Brown D.C.W. High efficiency plant production of North American ginseng via somatic embryogenesis from cotyledon explants. Plant Cell Reports, 2006, 25(3): 166-173 (doi: ).
- Zhang J.Y., Sun H.J., Song I.J., Bae T.W., Kang H.G., Ko S.M., Kwon Y.I., Kim I.-W., Lee J., Park S.-Y., Lim P.-O., Kim Y., Lee H.-Y. Plant regeneration of Korean wild ginseng (Panax ginseng Meyer) mutant lines induced by γ-irradiation (60Co) of adventitious roots. Journal of Ginseng Research, 2014, 38(3): 220-225 (doi: 10.1016/j.jgr.2014.04.001).
- You X.L., Han J.Y., Choi Y.E. Plant regeneration via direct somatic embryogenesis in Panax japonicus. Plant Biotechnology Reports, 2007, 1: 5-9 (doi: 10.1007/s11816-007-0009-4).
- Zhao S.J., Wang J.H., Liang Y.L., Xu L.X. Somatic embryogenesis and plantlet regeneration from hairy roots transformed by Agrobacterium rhizogenes in Panax quinquefolium L. International Journal of Biomedical and Pharmaceutical Science, 2012, 6(Special Issue 1): 97-100.
- Kim J.Y., Kim D.H., Kim Y.C., Kim K.H., Han J.Y., Choi Y.E. In vitro grown thickened taproots, a new type of soil transplanting source in Panax ginseng. Journal of Ginseng Research, 2016, 40(4): 409-414 (doi: 10.1016/j.jgr.2016.05.003).
- El-Mougy N.S., El-Gamal N.G., Fotouh Y.O., Abd-El-Kareem F. Evaluation of different application methods of chitin and chitosan for controlling tomato root rot disease under greenhouse and field conditions. Research Journal of Agriculture and Biological Sciences, 2006, 2(5): 190-195.
- Kołodziej B. The effect of chitosan in American ginseng (Panax quinquefolium L.) protection. In: Polish Chitin Society, Monograph XI. Łódź, Poland, 2006: 183-192.
- Vasyukova N.I., Zinoveva S.V., Ilinskaya L.I., Perekhod E.A., Chalenko G.I., Gerasimova N.G., Il'ina A.V., Varlamov V.P., Ozeretskovskaya O.L. Modulation of plant resistance to diseases by water-soluble chitosan. Applied Biochemistry and Microbiology, 2001, 37: 103-109 (doi: 10.1023/A:1002865029994).