doi: 10.15389/agrobiology.2020.5.945rus
УДК 633.11:581.1:631.816.3:57.04
В исследованиях использовали приборную базу Центра коллективного пользования «Агидель» УФИЦ РАН. Работа выполнена при частичной поддержке государственного задания Минобрнауки России № 075-00326-19-00 по теме № АААА-А18-118022190099-6.
СОДЕРЖАНИЕ ФИТОГОРМОНОВ И РОСТ ПОБЕГОВ У ПШЕНИЦЫ (Triticum durum Desf.) ПОД ВЛИЯНИЕМ ГУМАТОВ НАТРИЯ В СОСТАВЕ ГРАНУЛИРОВАННЫХ ОРГАНОМИНЕРАЛЬНЫХ УДОБРЕНИЙ
А.М. НАЗАРОВ1 ✉, И.Н. ГАРАНЬКОВ1, И.О. ТУКТАРОВА1, Э.Р. САЛЬМАНОВА1, Т.Н. АРХИПОВА2, И.И. ИВАНОВ2,
А.В. ФЕОКТИСТОВА2, З.Г. ПРОСТЯКОВА2, Г.Р. КУДОЯРОВА2
Гуминовые вещества образуются в почве в процессе разложения органических остатков и способны стимулировать рост и повышать продуктивность растений. В растениеводстве широко применяют наиболее растворимые препараты на основе гуматов натрия и калия, которые предположительно обладают гормоноподобной активностью. Однако механизм влияния гуминовых веществ на физиологические и биохимические процессы в растениях до сих пор не до конца понятен. Гуминовые вещества могут оказывать стимулирующее действие в очень низких концентрациях, что определяет требования к адресности и режиму их поступления в растения. Этим требованиям соответствует размещение удобрения в непосредственной близости от семян. Ранее было показано положительное влияние инкрустации гранул нитрофоски гуматом натрия (ГН) на урожайность яровой твердой пшеницы. В настоящем исследовании впервые представлены данные об изменении ростовых процессов и гормонального баланса у растений пшеницы (сорт Башкирская 27) при применении ГН в составе органоминерального удобрения (ОМУ). Гранулы удобрений без ГН и содержащие 1,25×10-2 %, 2,5×10-2 % и 5×10-2 % гуминового препарата (по массе гранул) размещали на расстоянии 2-3 см от высаженных в почву 1-суточных проростков пшеницы. ОМУ получали на основе отхода птицеводства (куриный помет) и доломита, ГН — экстракцией из бурого угля. Контролем служили растения, которые не получали ни гуматов, ни дополнительного питания в виде ОМУ. С 3-х сут после посадки измеряли длину листьев и эвапотранспирацию (по уменьшению массы сосудов с растениями). Образцы листьев для определения фитогормонов с помощью иммуноферментного анализа собирали на 9-е сут, площадь листьев, массу растений и содержание общего азота определяли на 21-е сут после посадки. Органоминеральные гранулы без ГН стимулировали удлинение листьев, что проявлялось в достоверно большей их длине по сравнению с контролем (42, 156, 187 и 274 против 47, 167, 199 и 294 мм, р ≤ 0,05), зарегистрированной для первого листа через 3, 6 и 8, для второго — через 14 сут после посадки. При этом увеличение длины листьев не зависело от дозы ОМУ, повышение которой не приводило к достоверному изменению анализируемого показателя. Добавление гуминовых веществ к гранулам ОМУ дополнительно стимулировало удлинение листьев, что наиболее ярко проявлялось в варианте с промежуточной концентрации ГН (2,5×10-2 %), при которой за время наблюдения (через 3, 6, 8 и 14 сут после посадки) была выявлена достоверная прибавка длины листьев по сравнению с эффектом от применения ОМУ без ГН (6,0; 11,0; 13,0 мм прибавки по первому листу и 9,4; 9,0; 22,0 — по второму, р ≤ 0,05). В отличие от ОМУ без ГН, применение которых не изменяло массу побега и площадь листьев, сочетание ОМУ с ГН в промежуточной концентрации достоверно увеличивало массу побегов (с 538 до 583 мг, р ≤ 0,05). Площадь листьев достоверно превышала контроль в вариантах с промежуточной и максимальной концентрациями ГН (на 385 и 283 мм2, р ≤ 0,05). Таким образом, добавление ГН к ОМУ повышало эффективность воздействия органоминерального удобрения на ростовые показатели. Накопление азота в побегах растений пшеницы, которые получали только ОМУ без гумата, не отличалось от контроля, а при сочетании удобрения с ГН — было достоверно (на 8-15 %, р ≤ 0,05) выше, чем в контроле. Внесение ОМУ без ГН не отражалось на содержании изученных фитогормонов (ауксинов, цитокининов и абсцизовой кислоты — АБК), тогда как добавление гумата в гранулы удобрения приводило к достоверному повышению содержания АБК и цитокининов в побегах — соответственно на 1,6-2,8 нг · г-1 (в 1,5-1,8 раза) и на 3,8-4,9 нг · г-1 (в 1,5-1,7 раза) (р ≤ 0,05). Содержание индолилуксусной кислоты было выше, чем в контроле (40 против 15 нг · г-1, р ≤ 0,05) при применении промежуточной концентрации ГН. Полученные нами результаты позволяют объяснить бóльшую эффективность удобрений в случае добавления гуматов их влиянием на количество фитогормонов. Предполагается, что повышение содержания гормонов со стимулирующим типом действия обеспечивает активацию роста растений, в то время как накопление АБК ограничивает транспирационные потери воды. Результаты этих лабораторных опытов свидетельствуют о перспективности практического применения гранул ОМУ с включением гумата натрия по предлагаемой нами технологии. Кроме того, полученные данные важны для выяснения фундаментальных механизмов, посредством которых проявляется воздействия гуминовых веществ, в частности на уровне гормонального баланса растений.
Ключевые слова: Triticum durum, органоминеральное удобрение, гумат натрия, ауксины, цитокинины, абсцизовая кислота, рост растений.
Гуминовые вещества, представленные большей частью гуминовыми кислотами и фульвокислотами и образующиеся в почве в процессе разложения органических остатков, способны стимулировать рост растений и повышать их урожайность (1), что используют в растениеводстве, где все более широко применяют препараты гуминовых соединений на основе гуматов натрия и калия, полученных щелочной экстракцией каустобиолитов (бурого угля, торфа, сапропеля). Предполагается, что гуминовые вещества влияют на урожайность растений как непосредственно, так и опосредованно. Опосредованное воздействие связывают с модификацией структуры почвы в ризосфере и увеличением доступности для растений элементов минерального питания, в то время как непосредственный эффект гуминовых соединений проявляется в изменении метаболизма и развития растения (2, 3).
Механизм влияния гуминовых веществ на физиологические и биохимические процессы в растениях до сих пор не до конца понятен (4). Гуминовые вещества с относительно небольшой массой образуют надмолекулярные ассоциации, состоящие из гидрофильных и гидрофобных доменов (5). Гидрофобные компоненты гуминовых веществ, образующиеся в результате разрушения растительных остатков, захватывают гидрофильные компоненты почвы, защищая их от деградации (6). Захваченные таким образом соединения могут освобождаться из гуминовых ассоциаций в результате изменения их структуры под влиянием корневых выделений, имеющих низкие значения рН (4). Непосредственное влияние гуминовых кислот на рост и развитие растений представляет собой проявление их гормоноподобной активности (7-9). В препаратах гуминовых кислот были выявлены ауксины (в основном в форме индолилуксусной кислоты) (7, 10) и цитокинины (в форме изопентениладенина) (11). Гуминовые кислоты влияют на активность ферментов, экспрессию генов и активность протонной помпы таким же образом, как и гормон растений ауксин (7-10). Есть сообщения, что гуминовые вещества благодаря такой активности изменяют архитектуру и метаболизм корней (12). Гормоноподобные свойства гуминовых препаратов обусловливают одно из преимуществ органических удобрений по сравнению с химическими. Однако влияние гуминовых кислот на содержание ауксинов и цитокининов в растениях пока что не изучено.
Привлекательности гуминовых соединений для практики растениеводства способствует тот факт, что они могут оказывать стимулирующее действие на рост растений в очень низких концентрациях (от 20 мг/л-1) (7). Это может обеспечить потенциально высокую рентабельность их применения. Вместе с тем при использовании действующего вещества в низких концентрациях возникает проблема адресности его применения и поступления в растения. Такую возможность открывает технология полосной обработки (strip till), когда с помощью специальных сеялок удобрения размещаются в непосредственной близости от семян.
Ранее мы показали положительное действие гумата натрия, включенного в гранулы нитрофоски, на урожайность яровой твердой пшеницы (13). В представляемой работе нами впервые выявлено влияние разных доз гумата натрия в составе органоминеральных удобрений (ОМУ) на рост и гормональный баланс растений пшеницы.
Цель исследования состояла в проверке гипотезы о том, что эффективность сочетания органоминеральных удобрений и гумата натрия связана с его действием на гормональную систему растений.
Методика. Гумат натрия получали по методике, аналогичной описанной (14, 15): к 5 г бурого угля (Кумертауское месторождение, Республика Башкортастан) добавляли 100 мл 3 % NaOH и перемешивали в течение 2 ч при 60 °С, нерастворенный осадок (гумин) отделяли центрифугированием, промывали дистиллированной водой. К полученному раствору добавляли 1 % HCl до pH = 2, осадок гуминовых кислот отделяли центрифугированием (10 мин , 4000 об/мин-1, 2325 g, центрифуга ПЭ 6910, «ООО ЭкросХим», Россия), промывали осадок дистиллированной водой, высушивали на воздухе и растворяли 0,1 г в 10 мл 1 % NaOH.
Концентрацию 0,01-0,001 % раствора гуминовых кислот в 0,4 % растворе NaOH определяли по величине оптической плотности (OD) методом УФ-спектрометрии (Shimadzu UV 2600, «Shimadzu», Япония, λ = 465 нм) согласно описанию (14, 15) .
Выделенный ГН вводили в состав органоминеральных удобрений (ОМУ), содержащих 60-65 % свежего куриного помета, 54-33 % доломита (карбонаты кальция и магния). В процессе получения гранул ОМУ в их состав включали ГН из расчета 1,25×10-2 %, 2,5×10-2 % и 5×10-2 % (соответственно ГН1, ГН2 и ГН3) от массы гранулы ОМУ. К части гранул гумат натрия не добавляли.
Гранулы ОМУ массой около 80 мг с разным содержанием гуминовых кислот размещали в вегетационных сосудах с землей (около 0,82 кг; на дне сосудов находился слой гальки высотой 2 см для дренажа) на глубине около 5-6 см от поверхности почвы из расчета одна или 3 гранулы на семя (80 или 240 мг). Проросшие за 1 сут на фильтровальной бумаге семена твердой яровой пшеницы (Triticum durum Desf.) сорта Башкирская 27 располагали на глубине 3 см от поверхности почвы (3 см от гранул) по 10 семян на сосуд, по 8 сосудов в каждом варианте опыта. В качестве контроля служили растения, которые не получали дополнительного питания в виде ОМУ. Растения выращивали на светоплощадке (ФАР 400 мкмол/м-2, 14 ч световой день, 20/24 °C). Влажность почвы поддерживали на уровне 60-80 % от полной влагоемкости с поливом 2-3 раза в неделю. Начиная с 3-х сут после высадки проростков, измеряли длину 1-го и 2-го листьев. Через 21 сут после посадки измеряли массу побегов растений, а также площадь сканированных изображений листьев с помощью программы ImageJ (National Institute of Health, США; https://imagej.nih.gov).
Содержание гормонов определяли на 9-е сут после посадки растений. Фитогормоны экстрагировали из измельченных побегов 80 % этиловым спиртом при температуре 4 °C в течение ночи. Очистку и концентрирование индолилуксусной кислоты (ИУК) и абсцизовой кислоты (АБК) из аликвоты водного остатка после упаривания спиртового экстракта (из чашек Петри в токе воздуха) проводили с помощью эфирной экстракции по модифицированной схеме с уменьшением объема согласно описанию (16). ИУК и АБК дважды экстрагировали диэтиловым эфиром из подкисленной до рН 2-3 аликвоты водного остатка в соотношении 1:3, затем возвращали их в водную фазу (1 % раствор гидрокарбоната натрия, соотношение водной и органической фазы 1:2), снова подкисляли до рН 2-3 и после 2-кратной реэкстракции диэтиловым эфиром метилировали диазометаном, который получали из нитрозометилмочевины, и прибавляли к образцам. Образцы высушивали и непосредственно перед проведением иммуноанализа раство-ряли в небольшом количестве 80 % этанола, а перед проведением иммуноферментного анализа растворяли в 100 мкл 80 % этилового спирта. Цитокинины из аликвоты водного остатка концентрировали на картридже С18 («Waters Corporation», США). Для этого картридж С 18 уравновешивали дистиллированной водой и пропускали через него образец, предварительно очищенный центрифугированием . Колонку с нанесенным образцом промывали 20 мл дистиллированной воды. Цитокинины элюировали 70 % этиловым спиртом, затем спирт упаривали досуха и, растворив остаток в 20 мкл 80 % этанола, наносили образце на силуфоловую пластину для тонкослойной хроматографии (ТСХ). ТСХ проводили в системе растворителей бутанол: аммиак: вода (6:1:2) , как описано ранее (17). После детекции в УФ- свете (трансиллюминатор TCP-15.MC с лампами 4× 8 W 312 nm, «Vilber Lourmat», Франция) расположения метчиков зеатина, зеатиннуклеотида и зеатинрибозида содержимое зон каждого образца элюировали 0,1 М фосфатным буфером (рН 7,2-7,4). Для осаждения силуфола элюат центрифугировали 10 мин при 10000 об/мин-1 (Eppendorf MiniSpin, «Eppendorf», США). Количественное определение ИУК, АБК и цитокининов проводили с помощью иммуноферментного анализа (ИФА) с использованием специфических антител к гормонам (16-18). Антитела к зеатинрибозиду использовали также для определения зеатина и его нуклеотида, так как они обладают перекрестной реактивностью к этим цитокининам.
Иммуноферментный анализ проводили в лунках полистироловых планшетов марки Castar («Corning Incorparated», США). Конъюгат гормона с белком сорбировали на полистироле. Предварительно разбавленный конъюгат в 0,05 М посадочном буфере (9 % NaCl) разливали по 200 мкл в каждую лунку и выдерживали при температуре 4 °C в течение 18-20 ч в холодильнике или 2 ч в термостате при 37 °C . Трижды промывали планшеты физиологическим раствором (рН 7,2-7,4) с 0,05 % Tween 20 (раствор ФТ). На всех последующих стадиях промывки использовали этот же раствор. В часть лунок разливали стандарты гормона с 10-кратным разведением для получения калибровочной кривой, в остальные лунки — аликвоты спиртовых растворов мети лированных гормонов (ИУК, АБК) или аликвоты фосфатного буфера (рН 7,2-7,4), которым элюировали цитокинины из силуфола. Во все лунки вносили антисыворотки к соответствующему гормону (по 100 мкл на лунку), полученные, как описано (18, 19), и разбавленные физиологическим раствором с 0,3 % бычьего сывороточного альбумина и 0,05 % Tween 20 (ФТБ). Инкубировали в термостате при температуре 37 °C в течение 1 ч и промывали раствором ФТ. Для визуализации реакции и количественной оценки прореагировавших антител сыворотки с сорбированным в лунках белковым конъюгатом гормонов использовали препарат антикроличьих бычьих антител, меченных пероксидазой. Препарат разводили в ФТБ, разливали в лунки по 200 мкл, инкубировали 1 ч при 37 °C , затем промывали ФТ. Количество иммуносорбированных антител определяли по цветной реакции с субстратом — ортофенилендиамином (0,4 мг/мл в 0,06 М фосфатном буфере, pH 5,2, с 0,006 % перекиси водорода). Цветную реакцию через 15-30 мин останавливали 4 н. серной кислотой. После измерения оптической плотности ( фотометр АИФР-01 УНИПЛАН, ЗАО «ПИКОН », Россия; λ = 492 нм) рассчитывали содержание гормона по калибровочной кривой.
По окончании эксперимента (на 21-сут) определяли содержание общего азота по Кьельдалю. Эвапотранспирацию оценивали на протяжении всего эксперимента по уменьшению массы сосудов с растениями.
Статистические показатели рассчитывали стандартными методами с использованием программы MS Excel. Вычисляли средние (M) и их стандартные ошибки (±SEM). Различия оценивали по t-критерию Стьюдента и считали статистически значимыми при p ≤ 0,05. Число повторностей: 5 — при анализе содержания азота, 9 — гормонов, 30 — при учете водопотребления и 50 — при оценке массы растений, площади и длины листьев (указано в подрисуночных подписях).
Результаты. В процессе получения ОМУ при смешивании куриного помета с доломитом происходит образование магниевых и кальциевых солей мочевой и других органических кислот, содержащихся в курином помете, что снижает их растворимость и возможность негативного действия в высоких концентрациях на рост растений. Кроме того, нагревание продукта до 100 °С за счет трения (в процессе смешения и экструдирования) способствует уничтожению содержащихся в курином помете патогенных микроорганизмов.
Как видно из рисунка 1, внесение гранул органоминерального удобрения увеличивало длину листьев на 5-17 % по сравнению с контролем в зависимости от возраста листьев и варианта опыта. Через 3 сут после посадки семян листья растений были достоверно длиннее, чем в контроле (при р ≤ 0,05), только при меньшей дозе ОМУ (одна гранула вместо трех) и промежуточной концентрацией гуматов (ГН2) (см. рис. 1, А). По мере роста растений влияние ОМУ и ГН проявлялось и при других дозах и концентрациях: на 8-е сут — 1-е листья, на 14-е сут — 2-е листья были достоверно длиннее (при р ≤ 0,05), чем в контроле, во всех вариантах с внесением ОМУ (с гуматом и без него) (см. рис. 1). Увеличение дозы ОМУ не приводило к достоверному увеличению длины листьев по сравнению с минимальной дозой ОМУ (прибавка длины по сравнению с контролем в среднем составила 7,5 и 6,0 % при внесении соответственно одной и трех гранул на семя).
Добавление гуминовых веществ к гранулам ОМУ увеличивала его стимулирующее действие (по сравнению с контролем) на удлинение листьев, что наиболее ярко проявлялось на фоне ГН2 (2,5×10-2 % от массы гранулы). При этом содержании средняя прибавка длины листьев по сравнению с контролем за все время наблюдения (через 3, 6, 8 и 14 сут после посадки) составила в среднем 10 % (различия по сравнению с эффектом от применения ОМУ без ГН были достоверны при р ≤ 0,05, парный t-тест).
Через 3 нед после посадки применение ОМУ в сочетании с проме-жуточной концентрацией гумата натрия (ГН2) приводило к достоверному повышению сырой массы побегов по сравнению с контролем (рис. 2, А). При использовании ОМУ без ГН достоверного увеличения массы побега относительно контроля мы не выявили. По этому признаку растения на фоне ГН1 и ГН3 занимали промежуточное положение между контролем и вариантом со средней концентрацией гуматов (ГН2), то есть их показатели статистически значимо не отличались от таковых ни в первом, ни во втором случае. Таким образом, нами выявлена тенденция повышения биомассы растений пшеницы под влиянием сочетания ОМУ с ГН, которая была статистически достоверной при промежуточной из использованных концентраций ГН. В вариантах с ГН2 и ГН3 суммарная площадь листьев была достоверно больше , чем в контроле (2637 и 2535 против 2252 мм2, р ≤ 0,05), в то время как применение ОМУ без ГН и ОМУ в сочетании с минимальной концентрацией гуматов (ГН1) не имело существенного влияния на этот показатель (различия по сравнению с контролем недостоверны) (см. рис. 2, Б).
По накоплению общего азота в побегах растения пшеницы, которые получали органоминеральное удобрения без гумата, не отличались от контроля (см. рис. 2, В), а в вариантах с ГН накопление общего азота в побегах было достоверно (р ≤ 0,05) выше, чем в контроле.
По эвапотранспирации, которую регистрировали на протяжении всего эксперимента, растения, получавшие удобрения, не отличались от контроля, и во всех вариантах показатель составлял 1,6 г воды на растение в сутки (данные не приведены).
При внесении ОМУ без добавления ГН, содержание гормонов (ауксинов, цитокининов и АБК) у растений пшеницы было на уровне контроля (рис. 3), в то время как добавление гумата натрия приводило к повышению содержания гормонов. Для подтверждения результатов иммуноферментного анализа ранее проводили сравнение данных ИФА и хромато-масс-спектрометрии (17).
Суммарная концентрация цитокининов была стабильно выше, чем в контроле, при действии всех испытанных концентраций ГН (см. рис. 3, А , различия достоверны при р ≤ 0, 05), в то время как применение ОМУ без ГН не приводило к повышению уровня цитокининов по сравнению с контролем. Накопление ауксинов проявлялось при промежуточной концентрации гумата, при которой уровень ИУК был почти в 3 раза выше, чем в контроле (различия по сравнению с контролем достоверны при р ≤ 0,05) (см. рис. 3, Б). Содержание АБК было выше при действии всех испытанных концентраций ГН (различия по сравнению с контролем и вариантами с ОМУ без ГН достоверны при р ≤ 0,05) (см. рис. 3, В).
В целом результаты оценки длины листьев, их площади и массы побега показали, что добавление гумата к органоминеральным удобрениям усиливает их способность стимулировать рост растений пшеницы. Также установлено, что ОМУ действует в низких дозах (одна гранула на семя), и увеличение дозы до трех гранул не усиливает положительного эффекта.
Повышение накопления азота в побегах при внесении в почву гранул ОМУ с ГН свидетельствует об улучшении способности растений пшеницы поглощать элементы минерального питания под влиянием гумата, что соответствует некоторым данным литературы (20). Поскольку неоднократно показано свойство азота стимулировать синтез цитокининов (21), их аккумуляцию в побегах растений, получавших ГН, можно связать с ростом содержания азота. Вместе с тем количество общего азота в побегах под влиянием ГН возрастало менее значительно (максимум на 15 %), чем цитокининов, содержание которых было в 1,5 раза больше, чем в контроле, независимо от количества ГН в препарате. Отсутствие пропорциональности между накоплением азота и цитокининов позволяет предполагать существование дополнительных механизмов стимуляции накопления в растениях цитокининов при действии гуматов натрия (например, при их непосредственном влиянии на гормоны растений).
Альтернативное объяснение может заключаться в присутствии в препарате ГН гормоноподобных соединений, что соответствует данным литературы (1). Поскольку физиологически активные вещества, в том числе гормоны, по определению действуют на растения в низких концентрациях, именно присутствием гормоноподобных веществ можно объяснить способность столь малых доз ГН (как в нашем эксперименте) воздействовать на рост и развитие растений. В гуминовых препаратах обнаружены ауксино-подобные вещества (4), что соответствует результатам выполненных нами экспериментов, в которых выявлено повышение содержание ауксинов в растениях под влиянием ГН. Также было установлено, что применение гуминового препарата увеличивает доступность азота, фосфора и калия (11).
Содержание гуминовых кислот в свежем помете составляло 20-23 %, а в гранулированном ОМУ, полученном на основе куриного помета и доломита, — 15-17 %. Однако растворимость свободных гуминовых кислот очень низка, в то время как растворимость их натриевых солей достаточно большая. Поэтому высокая эффективность действия гранулированного ОМУ, содержащего хорошо растворимые гуматы натрия, на рост растений может быть обеспечена его высокой биоразлагаемостью.
В.В. Демин с соавт. (22) считают непосредственное проникновение гуматов в клетку маловероятным, поскольку этому должно препятствовать формирование ими водородных связей с компонентами клеточных стенок, но возможно поступление гуминовых веществ в клетку за счет эндоцитоза и их дальнейшего переваривания в лизосомах. Известно, что рецепторы многих гормонов находятся на поверхности клеток (23), что позволяет объяснить, как гормоноподобные компоненты, содержащиеся в гуматах, могут влиять на растения, взаимодействуя с рецепторами на поверхности клеток, не проникая в них.
Эти эксперименты не позволяют судить о том, связано ли повышенное содержание гормонов с их поступлением из препарата ГН (вероятность этого процесса невелика, учитывая большие размеры молекул гуматов) (24). Тем не менее сам факт накопления гормонов у растений, обработанных использованным нами препаратом, — это яркий индикатор физиологической активности ГН. Важно то, что при воздействии на растения ОМУ без гумата увеличения количества гормонов не регистрировали, то есть повышенное содержание гормонов у растений является характерной реакцией именно на присутствие ГН в препаратах. Хорошо известна способность цитокининов и ауксинов стимулировать рост побега, активируя процессы деления и растяжения клеток (25). Это свойство позволяет объяснить активацию роста побегов пшеницы под влиянием ГН, отмеченную в наших опытах, накоплением ауксинов и цитокининов.
Накопление АБК, обнаруженное нами при введении гранул с гуматом натрия в среду обитания корней растений, соответствует некоторым данным литературы (26). Авторы цитируемого сообщения связывают накопление АБК с особенностями водного обмена у обработанных гуматом растений. В наших опытах выявлено достоверно большее накопление АБК под влиянием гуматов, содержащихся в препаратах (почти 2-кратное по сравнению с контролем, различия с контролем и вариантом с внесением ОМУ без ГН достоверны при р ≤ 0,05). Как известно, испарение воды листьями возрастает с увеличением их площади. В нашем эксперименте площадь листьев была достоверно больше по сравнению с контролем при промежуточной и максимальной дозе ГН в ОМУ. Однако достоверного повышения транспирационных потерь (несмотря на большую площадь листьев) мы при этом не выявили, что указывает на снижение устьичной проводимости. Поскольку хорошо известна способность АБК закрывать устьица, можно объяснить ограничение транспирационных потерь повышением содержания АБК у растений под действием ГН. Ранее нами было показано, что активация роста листьев бактериями-фитостимуляторами сопровождалась накоплением АБК, что также ограничивало эвапотранспирацию (18). Таким образом, изменение накопления АБК под влиянием гумата можно рассматривать как адаптивную реакцию, направленную на оптимизацию водного обмена при его действии на растения.
Итак, нами показано, что включение гумата натрия (ГН) в гранулы органоминерального удобрения (ОМУ) активирует рост растений, что, скорее всего, обусловлено способностью ГН влиять на гормональный статус растений. Накопление гормонов стимулирующего действия обеспечивает активацию роста растений, в то время как накопление АБК нормализует их водный обмен. В нашем опыте размещением гранул ОМУ на расстоянии нескольких сантиметров от семян моделировалось применение оборудования для посева по технологии strip till. Полученные результаты лабораторных опытов позволяют прогнозировать перспективность применения гранул ОМУ с включением гумата натрия по предложенной технологии.
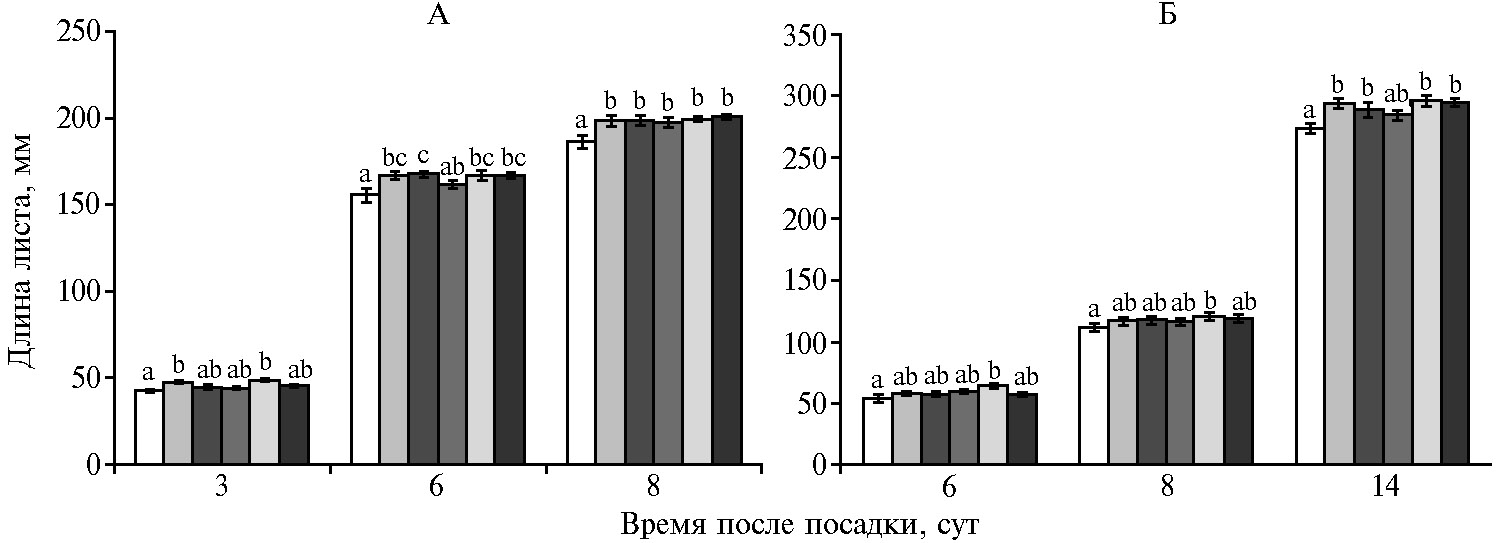
Рис. 1. Длина 1-го (А) и 2-го (Б) листьев у растущих побегов яровой твердой пшеницы (Triticum durum Desf.) сорта Башкирская 27 в зависимости от доз органоминерального удобрения (ОМУ, мг/семя) и их сочетания с разным количеством гумата натрия (ГН1 — 1,25×10-2 %, ГН2 — 2,5×10-2 %, ГН3 — 5×10-2 % от массы гранулы ОМУ). Столбики слева направо — контроль, ОМУ 80 мг, ОМУ 240 мг, ОМУ 80 мг + ГН1, ОМУ 80 мг + ГН2, ОМУ 80 мг + ГН3. Различия между показателями, для которых нет совпадения ни по одному буквенному обозначению, статистически значимы при р ≤ 0,05 (M±SEM, n = 50, вегетационный опыт).
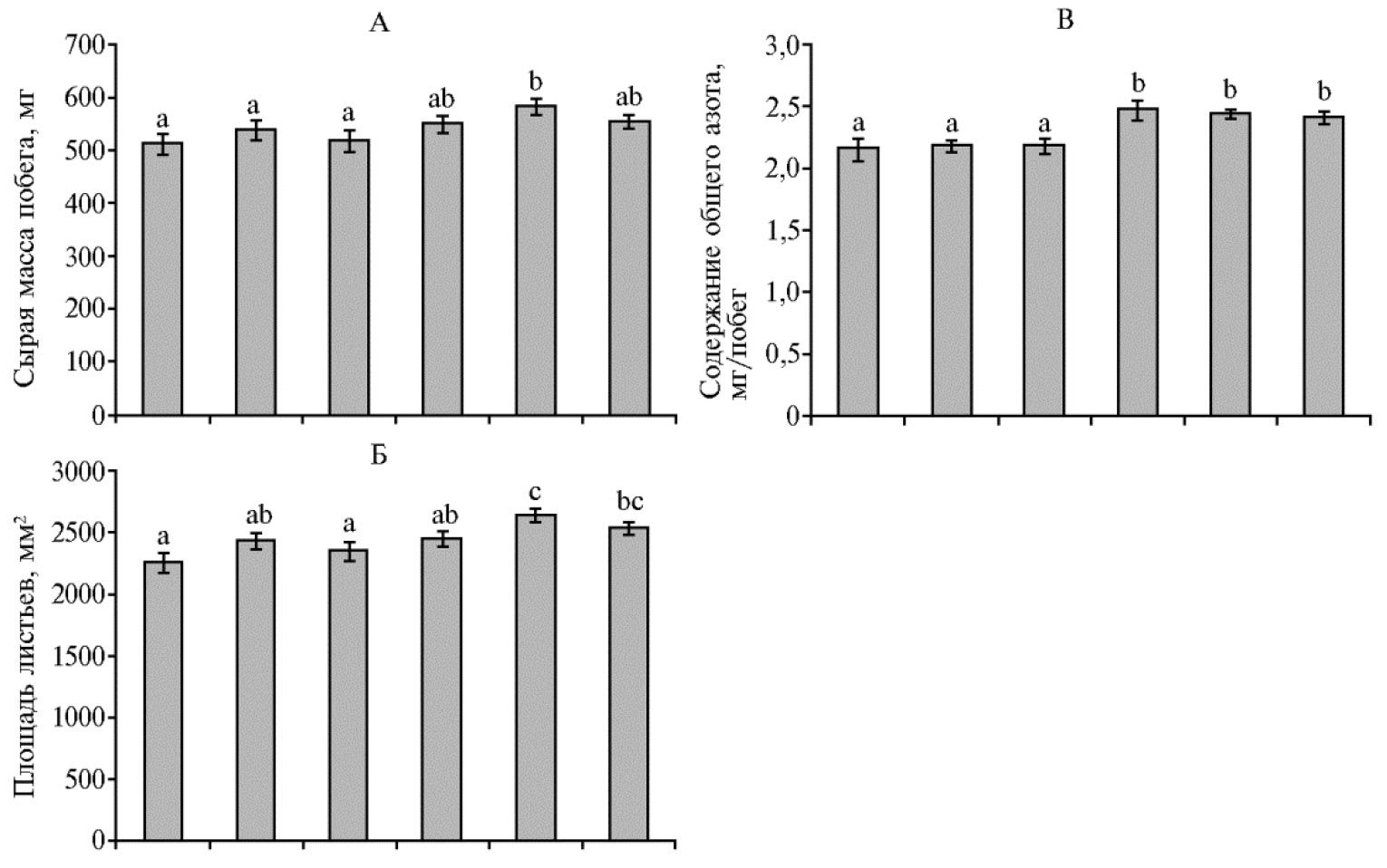
Рис. 2. Масса побега (А), площадь всех листьев (Б) и содержание общего азота (В) в побегах яровой твердой пшеницы (Triticum durum Desf.) сорта Башкирская 27 через 21 сут после посадки в зависимости от доз органоминерального удобрения (ОМУ, мг/семя) и их сочетания с разным количеством гумата натрия (ГН1 — 1,25×10-2 %, ГН2 — 2,5×10-2 %, ГН3 — 5×10-2 % от массы гранулы ОМУ). Столбики слева направо — контроль, ОМУ 80 мг, ОМУ 240 мг, ОМУ 80 мг + ГН1, ОМУ 80 мг + ГН2, ОМУ 80 мг + ГН3. Различия между показателями, для которых нет совпадения ни по одному буквенному обозначению, статистически значимы при р ≤ 0,05 (M±SEM, А и Б — n = 50, В — n = 5, вегетационный опыт).
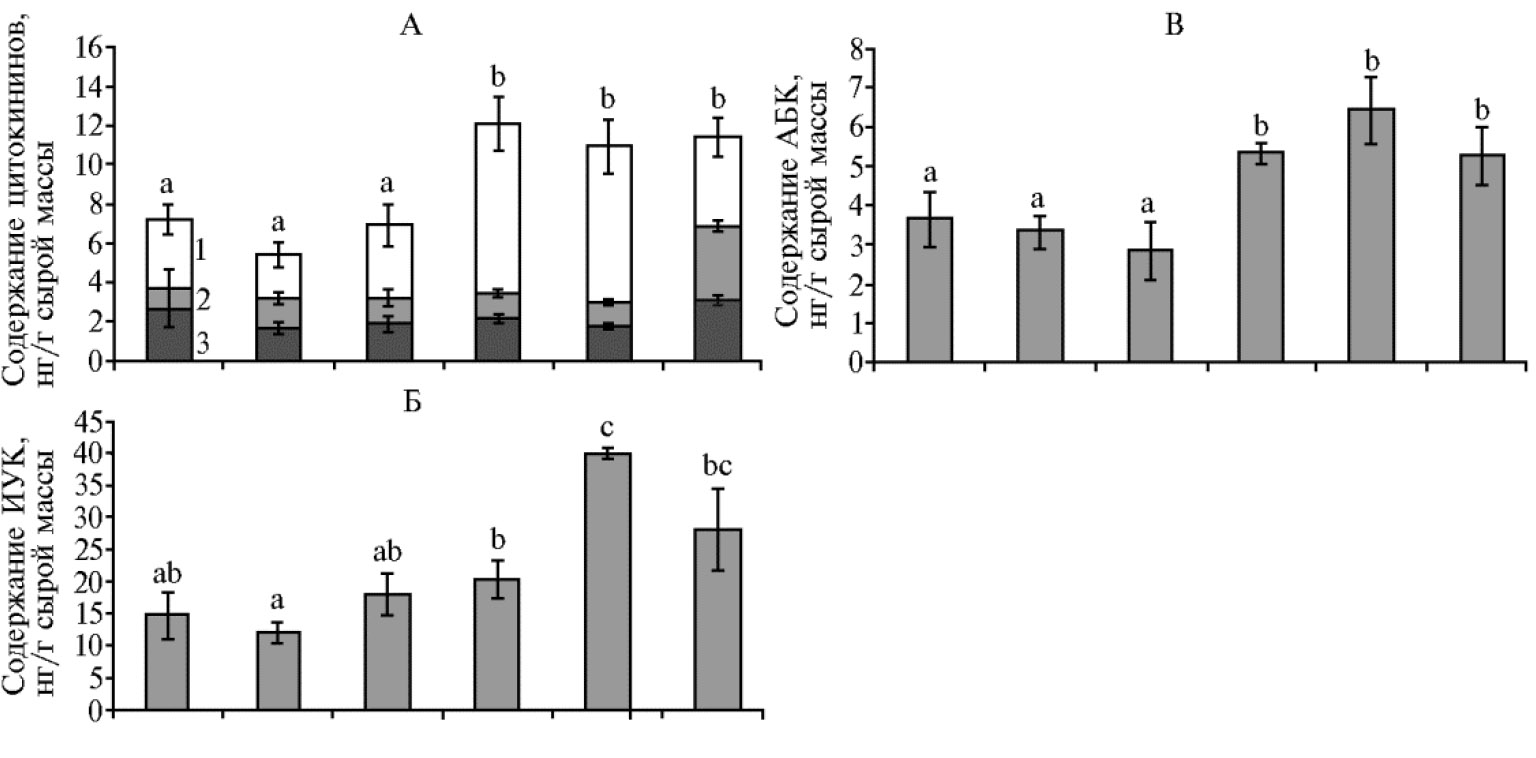
Рис. 3. Содержание цитокининов (А, 1 — зеатин, 2 — рибозид зеатина, 3 — риботид зеатина), ауксинов (Б, индолилуксусная кислота, ИУК) и абсцизовой кислоты (В, АБК) в побегах яровой твердой пшеницы (Triticum durum Desf.) сорта Башкирская 27 через 9 сут после посадки в зависимости от доз органоминерального удобрения (ОМУ, мг/семя) и их сочетания с разным количеством гумата натрия (ГН1 — 1,25×10-2 %, ГН2 — 2,5×10-2 %, ГН3 — 5×10-2 % от массы гранулы ОМУ). Столбики слева направо — контроль, ОМУ 80 мг, ОМУ 240 мг, ОМУ 80 мг + ГН1, ОМУ 80 мг + ГН2, ОМУ 80 мг + ГН3. Различия между показателями, для которых нет совпадения ни по одному буквенному обозначению, статистически значимы при р ≤ 0,05 (M±SEM, n = 9, вегетационный опыт).
ЛИТЕРАТУРА
- García A.C., Olaetxea M., Santos L.A., Mora V., Baigorri R., Fuentes M., Zamarreño A.M., Berbara R.L., Garcia-Mina J.M. Involvement of hormone- and ROS-signaling pathways in the beneficial action of humic substances on plants growing under normal and stressing conditions. BioMed Research International, 2016, 2016: 3747501 (doi: 10.1155/2016/3747501).
- Mora V., Olaetxea M., Bacaicoa E., Baigorri R., Fuentes M., Zamarreño A.M., Garcia-Mina J.M. Abiotic stress tolerance in plants: exploring the role of nitric oxideand humic substances. In: Nitric oxide in plants: metabolism and role in stress physiology /M.N. Khan, M. Mobin, F. Mohammad, F.J. Corpas (eds.). Springer, Cham, 2014: 243-264 (doi: 10.1007/978-3-319-06710-0_15).
- Chen Y., Aviad T. Effects of humic substances on plant growth. In: Humic substances in soil and crop sciences: selected readings /P. MacCarthy, C.E. Clapp, R.L. Malcolm and P.R. Bloom (eds.). American Society of Agronomy and Soil Sciences, Madison, 1990: 161-186 (doi: 10.2136/1990.humicsubstances.c7).
- Canellas L.P., Dantas D.J., Aguiar N.O., Peres L.E.P., Zsogon A., Olivares F.L., Dobbss L.B., Facanha A.R., Nebbioso A., Piccolo A. Probing the hormonal activity of fractionated molecular humic components in tomato auxin mutants. Annals of Applied Biology, 2011, 159(2): 202-211 (doi: 10.1111/j.1744-7348.2011.00487.x).
- Piccolo A. The supramolecular structure of humic substances. A novel understanding of humus chemistry and implications in soil science. Advances in Agronomy, 2002, 75: 57-134 (doi: 10.1016/S0065-2113(02)75003-7).
- Spaccini R., Piccolo A., Conte P., Haberhauer G., Gerzabek M.H. Increased soil organic carbon sequestration through hydrophobic protection by humic substances. Soil Biology and Biochemistry, 2002, 34: 1839-1851 (doi: 10.1016/S0038-0717(02)00197-9).
- Canellas L.P., Olivares F.L., Okorokova-Façanha A.L., Façanha A.R. Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence, and plasma membrane H+-ATPase activity in maize roots. Plant Physiology, 2002, 130: 1951-1957 (doi: 10.1104/pp.007088).
- Zandonadi D.B., Santos M.P., Busato J.G., Peres L.E.P., Façanha A.R. Plant physiology as affected by humified organic matter. Theoretical and Experimental Plant Physiology, 2013, 25(1): 12-25 (doi: 10.1590/S2197-00252013000100003).
- Aguirre E., Leménager D., Bacaicoa E., Fuentes M., Baigorri R., Zamarreño A.M., García-Mina J.M. The root application of a purified leonardite humic acid modifies the transcriptional regulation of the main physiological root responses to Fe deficiency in Fe-sufficient cucumber plants. Plant Physiol. Biochem., 2009, 47(3): 215-223 (doi: 10.1016/j.plaphy.2008.11.013).
- Jindo K., Martim S.A., Navarro E.C., Perez-Alfocea F., Hernandez T., Garcia C., Aguiar N.O., Canellas L.P. Root growth promoting by humic acids from composted and non-composted urban organic wastes. Plant and Soil, 2012, 353: 209-220 (doi: 10.1007/s11104-011-1024-3).
- Pizzeghello D., Francioso O., Ertani A., Muscolo A., Nardi S. Isopentenyladenosine and cytokinin-like activity of different humic substances. Journal of Geochemical Exploration, 2013, 129: 70-75 (doi: 10.1016/j.gexplo.2012.10.007).
- Trevisan S., Pizzeghello D., Ruperti B., Francioso O., Sassi A., Palme K., Quaggiotti S., Nardi S. Humic substances induce lateral root formation and expression of the early auxin-responsive IAA19 gene and DR5 synthetic element in Arabidopsis. PlantBiology, 2010, 12: 604-614 (doi: 10.1111/j.1438-8677.2009.00248.x).
- Трапезников В.К., Иванов И.И., Тальвинская Н.Г., Анохина Н.Л. Влияние инкрустации гранул нитрофоски гуматом натрия на урожай и качество зерна твердой пшеницы. Агрохимия, 2000, 9: 48-49.
- Орлов Д.С., Гришина Л.А. Практикум по химии гумуса. М., 1981.
- Hofrichter M., Steinbüchel A. Biopolymers, lignin, humic substances and coal. Wiley VCH, 2001.
- Юрков А.П., Веселова С.В., Якоби Л.М., Степанова Г.В., Кудоярова Г.Р., Шишова М.Ф. Содержание ауксина у люцерны хмелевидной (Medicago lupulina L.) при инокуляции грибом Rhizophagus irregularis на фоне низкой обеспеченности доступным фосфором. Сельскохозяйственнаябиология, 2017, 52(4): 830-838 (doi: 10.15389/agrobiology.2017.4.830rus).
- Kudoyarova G.R., Korobova A.V., Akhiyarova G.R., Arkhipova T.N., Zaytsev D.Yu., Prinsen E., Egutkin N.L., Medvedev S.S., Veselov S.Yu. Accumulation of cytokinins in roots and their export to the shoots of durum wheat plants treated with the protonophore carbonyl cyanide m-chlorophenylhydrazone (CCCP). Journal of Experimental Botany, 2014, 65(9): 2287-2294 (doi: 10.1093/jxb/eru113).
- Arkhipova T.N., Prinsen E., Veselov A.Yu., Martinenko E.V., Melentiev A.I., Kudoyarova G.R. Cytokinin producing bacteria enhance plant growth in drying soil. Plant and Soil, 2007, 292(1-2): 305-315 (doi: 10.1007/s11104-007-9233-5).
- Veselov S.Yu., Kudoyarova G.R., Egutkin N.L., Gyuli-Zade V.Z., Mustafina A.R., Kof E.M. Modified solvent partitioning scheme providing increased specificity and rapidity of immunoassay for indole- 3- acetic acid. Physiologia Plantarum, 1992, 86(1): 93-96 (doi: 10.1111/j.1399-3054.1992.tb01316.x).
- Calvo P., Nelson L., Kloepper J.W. Agricultural uses of plant biostimulants. Plant and Soil, 2014, 383(1-2): 3-41 (doi: 10.1007/s11104-014-2131-8).
- Kudoyarova G.R., Dodd I.C., Veselov D.S., Rothwell S.A., Veselov S.Y. Common and specific responses to availability of mineral nutrients and water. Journal of Experimental Botany, 2015, 66(8): 2133-2144 (doi: 10.1093/jxb/erv017).
- Демин В.В., Бирюков М.В., Семенов А.А., Завгородняя Ю.А. Природа биологического действия гуминовых веществ. Часть 1. Основные гипотезы. Доклады по экологическому почвоведению, 2006, 1(1): 72-79.
- Spartz A.K., Gray W.M. Plant hormone receptors: new perceptions. Genes and Development, 2008, 22(16): 2139-2148 (doi: 10.1101/gad.1693208).
- Baalousha M., Motelica-Heino M., Le Coustumer P. Conformation and size of humic substances: Effects of major cation concentration and type, pH, salinity, and residence time. Colloids and Surfaces A: Physicochemical and Engineering Aspects, 2006, 272(1-2): 48-55 (doi: 10.1016/j.colsurfa.2005.07.010).
- Schaller G.E., Bishopp A., Kieberc J.J. The Yin-Yang of hormones: cytokinin and auxin interactions in plant development. The Plant Cell, 2015, 27: 44-63 (doi: 10.1105/tpc.114.133595).
- Olaetxea M., Mora V., Bacaico E., Garnic M., Fuentes M., Casanova E., Zamarreño A.M., Iriarte J.C., Etayo D., Ederra I., Gonzalo R., Baigorri R., García-Mina J.M. Abscisic acid regulation of root hydraulic conductivity and aquaporin gene expression is crucial to the plant shoot growth enhancement caused by rhizosphere humic acids. Plant Physiology, 2015, 169: 2587-2596 (doi: 10.1104/pp.15.00596).