doi: 10.15389/agrobiology.2020.5.981rus
УДК 633.491:631.527:577.2:632
Работа выполнялась при поддержке подпрограммы «Развитие селекции и семеноводства картофеля в РФ». ДНК пробы сортов и гибридов картофеля, которые были использованы в MAS, пополнили банк ДНК препаратов образцов картофеля, поддерживаемый в рамках программы 0662-2019-0004.
ИСПОЛЬЗОВАНИЕ В ПРАКТИЧЕСКОЙ СЕЛЕКЦИИ КАРТОФЕЛЯ РЕЗУЛЬТАТОВ ДНК-МАРКИРОВАНИЯ ИСХОДНЫХ РОДИТЕЛЬСКИХФОРМ И МЕЖСОРТОВЫХ ГИБРИДОВ
Н.М. ГАДЖИЕВ1, В.А. ЛЕБЕДЕВА1, Д.А. РЫБАКОВ2, А.В. ИВАНОВ1, В.В. ЖЕЛТОВА2, Н.А. ФОМИНА2,
О.Ю. АНТОНОВА2, Т.А. ГАВРИЛЕНКО2 ✉
Успех селекционных исследований во многом определяется удачным подбором родительских форм для гибридизации. В последние годы при подборе пар в комбинациях скрещиваний наряду с традиционными подходами активно используются результаты маркер-опосредованного отбора (marker assisted selection, MAS) для комбинирования ценных аллелей родительских генотипов. Такой методический прием широко распространен в селекционных программах для различных сельскохозяйственных растений во многих странах, в том числе в России. Применение MAS перспективно как на начальном этапе при подборе родительских сортов для скрещиваний, так и при оценке расщепляющихся гибридных популяций. В представленной работе пары для межсортовых скрещиваний подбирали таким образом, чтобы они дополняли друг друга, объединяя у гибридов доминантные аллели разных R-генов устойчивости к различным болезням и вредителям и хозяйственно ценные признаки родительских сортов. Для повышения результативности отбора перспективных гибридных генотипов использовали комплексный подход, сочетающий MAS c маркерами R-генов устойчивости к разным болезням и вредителям с традиционными методами оценки хозяйственно ценных признаков гибридных популяций. Полученные межсортовые гибриды трех комбинаций (Гусар × Чароит), (Гусар × Алый парус), (Гусар × Сиреневый туман) также участвовали в MAS с 8 маркерами шести R-генов: устойчивости к Y-вирусу картофеля PVY (Rysto) и X-вирусу картофеля PVX (Rx1), к золотистой Globodera rostochiensis (Wollenweber) Behrens (H1) и бледной G. pallida (Stone) Behrens (Gpa2) картофельным нематодам, расоспецифической устойчивости к фитофторозу (R1, R3а) (возбудитель Phytophthora infestans Mont. de Bary). Фактически все полученные гибриды обладали различными комбинациями ДНК маркеров R-генов. Для выявления аллельного состава R-генов у родительских форм был изучен характер наследования ДНК маркеров в каждой комбинации, что позволило определить уровень гетерозиготности маркированных локусов родительских сортов. По результатам оценки 144 гибридов в полевых и лабораторных условиях выделили 31 генотип с теми или иными хозяйственно ценными признаками (продуктивность, товарность, выравненность гнезд и клубней в гнезде, крахмалистость, полевая устойчивость к фитофторозу), 113 гибридов были забракованы. В отобранной группе, включающей 31 генотип, 23 гибридных генотипа имели относительно высокую продуктивность, которая варьировала от 600 до 1525 г/растение, и 12 из них характеризовались комплексом хозяйственно ценных признаков (высокие показатели продуктивности и товарности клубней, выравненность гнезд и клубней в гнезде), а также обладали различными комбинациями маркеров генов Rysto, Rx1, H1, Gpa2, R1 и R3а. Таким образом, использование комплексного подхода, сочетающего традиционные методы селекции и MAS, повышает результативность отбора перспективных межсортовых гибридов с заданным набором признаков. Отобранные гибридные генотипы представляют интерес для создания конкурентоспособных по продуктивности и качеству сортов с комплексной устойчивостью к различным возбудителям заболеваний и вредителям (в том числе с групповой устойчивостью к вирусам PVY и PVX, с групповой устойчивостью к разным видам нематод), что позволит сократить число химических обработок для защиты урожая.
Ключевые слова: картофель, сорта, гибриды, хозяйственно ценные признаки, устойчивость, R-гены, ДНК маркеры, MAS, PVY, PVX,вирусы картофеля, Globodera rostochiensis, золотистая картофельная нематода, Globodera pallida, бледная картофельная нематода, фитофтороз, Phytophthora infestans.
При выборе пар для гибридизации селекционеры используют разные подходы и методы, включая подбор родительских пар на основе эколого-географических особенностей, различий по параметрам продуктивности, продолжительности фаз вегетации или неодинаковой устойчивости к болезням и вредителям, а также по результатам оценки комбинационной способности родительских форм (1, 2). Однако, несмотря на многолетнюю историю исследований по разработке и совершенствованию методов подбора пар для скрещиваний, успех создания удачных гибридных комбинаций во многом остается мало предсказуемым (3). В последние годы при подборе пар для скрещиваний наряду с традиционными подходами активно используются результаты маркер-опосредованного отбора (marker assisted selection, MAS) для комбинирования ценных аллелей родительских генотипов. В селекции картофеля при подборе пар для скрещивания чаще всего применяют принцип комплементарности исходных форм в сочетании с MAS по ДНК маркерам разных генов устойчивости к болезням и вредителям (4, 5). Сочетание MAS с маркерами R-генов устойчивости с традиционными методами отбора гибридных генотипов по урожайности и другим агрономически ценным признакам повышает эффективность селекционного процесса, что показали зарубежные программы по созданию нематодоустойчивого (6) и вирусоустойчивого (7, 8) селекционного материала, а также по отбору новых перспективных клонов картофеля с комплексной устойчивостью к разным патогенам (9-13). Исследования в рамках аналогичных программ ведутся и в России (14-16).
Поскольку сорта картофеля представлены высокогетерозиготными тетраплоидными генотипами, у межсортовых гибридов отмечается расщепление по изучаемым признакам, в том числе по наличию/отсутствию ДНК маркеров определенного локуса. Данные о наследовании ДНК маркеров доминантных аллелей R-генов устойчивости у межсортовых гибридов служат основой для отбора перспективных гибридных генотипов в потомстве. Кроме того, информация об аллельном составе и степени гетерозиготности маркированных локусов у родительских сортов дает возможность прогнозировать частоту появления в потомстве гибридов, устойчивых к определенному патогену, и планировать объемы анализируемых расщепляющихся популяций (17-21). В зависимости от степени гетерозиготности маркированного R-локуса у исходного сорта, частота, с которой в гибридных популяциях появляются устойчивые генотипы, несущие хотя бы один доминантный аллель, будет составлять 100 % в случаях квадриплекса (RRRR) или триплекса (RRRr), 83,4 % — в варианте дуплекса (RRrr) и 50 % — для симплекса (Rrrr) (17).
На основе оригинальных многовидовых гибридов были созданы сорта картофеля, обладающие хозяйственно ценными признаками и адаптированные к условиям Северо-Западного региона России (22-24). Полученные нами ранее результаты MAS этих сортов (25, 26) детектировали разные ДНК маркеры, ассоциированные с генами устойчивости к возбудителю рака картофеля, наиболее распространенным и вредоносным Y и X вирусам картофеля (PVY и PVX), маркеры генов расоспецифической устойчивости к возбудителю фитофтороза Phytophthora infestans Mont. de Bary, а также маркеры генов устойчивости к разным видам цистообразующих нематод — золотистой картофельной нематоде (ЗКН) Globodera rostochiensis (Wollenweber) Behrens и бледной картофельной нематоде (БКН) G. pallida (Stone) Behrens. Отбор устойчивых к ЗКН и БКН генотипов с применением фитопатологических методов очень сложен, так как эти виды нематод относятся к объектам соответственно внутреннего и внешнего карантина. Информация о наличии в селекционном материале маркеров генов устойчивости к G. pallida представляет особую ценность: этот вид нематоды в Российской Федерации пока не обнаружен, хотя велика опасность его заноса извне (27).
Основываясь на данных MAS (26), в представляемой работе подобрали родительские пары и провели серию межсортовых скрещиваний с целью объединения в одном генотипе SCAR маркеров R-генов устойчивости к различным болезням и вредителям. Материнской формой в этих скрещиваниях служил сорт Гусар с маркерами генов устойчивости к PVY и ЗКН. Использовать указанный сорт в качестве опылителя невозможно из-за присущей этому сорту цитоплазматической мужской стерильности (26, 28). Отцовскими формами в трех комбинациях скрещиваний были сорта Чароит, Алый парус и Сиреневый туман, у которых выявлены ДНК маркеры генов устойчивости к PVX и к БКН. В рамках представлемого исследования расщепляющиеся гибридные популяции, полученные в трех комбинациях скрещиваний, вовлекались в MAS для отбора перспективных генотипов с различным сочетанием маркеров доминантных аллелей R-генов устойчивости к разным болезням и вредителям и поиска среди них перспективных клонов с комплексом агрономически ценных признаков (продуктивность, товарность, выравненная поверхность клубней, крахмалистость).
Цель работы состояла в повышении результативности отбора перспективных межсортовых гибридов, на основе использования комплексного подхода, сочетающего MAS и традиционные методы селекции.
Методика. Исследования проводили в 2017-2019 годах (Ленинградский НИИ сельского хозяйства «Белогорка», Гатчинский р-н, Ленинградская обл.).
Родительские пары для межсортовых скрещиваний подбирали с учетом сведений о хозяйственно ценных признаках сортов (продуктивность, качество клубней, устойчивость к наиболее распространенным болезням), созданных в разные годы и адаптированных к условиям Северо-Западного региона России (оригинаторы Ленинградский НИИ сельского хозяйства «Белогорка», ООО Селекционная фирма «ЛиГа», Ленинградская обл.) (29-31). При подборе также учитывали результаты выполненного нами ранее молекулярного скрининга этих сортов (26), в котором использовались ДНК маркеры, ассоциированные с R-генами устойчивости к вирусам PVY и PVX, ЗКН и БКН и с генами расоспецифической устойчивости к фитофторозу. В гибридизацию вовлекали сорта с маркерами разных R-генов. Скрещивания проводили в 2017 году в трех комбинациях — Гусар × Чароит, Гусар × Алый парус и Гусар × Сиреневый туман.
В 2018 году гибридные семена высевали в теплицу и сеянцы пикировали в поле. В июле 2018 года от каждого гибридного генотипа были взяты листья для выделения ДНК, а в сентябре 2018 года — собраны клубни от каждого гибрида индивидуально. Весной 2019 года по результатам хранения клубней были отобраны 144 гибрида трех комбинаций и проведена их посадка, каждый гибридный генотип высаживали на 8-клубневой делянке площадью 2,3 м2. Отбор лучших гибридных генотипов проводили в 2019 году на опытном поле (Ленинградский НИИ сельского хозяйства «Белогорка», Ленинградская обл.) во время вегетации гибридных растений на основании оценки фитофтороустойчивости и устойчивости к вирусным болезням, а также при уборке и в лабораторных условиях по результатам оценки комплекса хозяйственно ценных признаков (продуктивность, товарный вид клубней, выравненность гнезд и клубней в гнезде, содержание сухих веществ и крахмала в клубнях).
MAS проводили для 112 из 144 гибридных генотипов (27 гибридов из комбинации Гусар × Чароит, 30 — Гусар × Алый парус и 55 — Гусар × Сиреневый туман).
Листья индивидуальных гибридных растений, собранные летом 2018 года на экспериментальном поле (Ленинградский НИИ сельского хозяйства «Белогорка», Ленинградская обл.), фиксировали в жидком азоте с последующей транспортировкой в сухом льду. Тотальную ДНК выделяли методом СТАВ-экстракции в предложенной нами модификации (32).
MAS межсортовых гибридов выполняли с 8 SCAR (sequence chаr-acterized amplyfied region) маркерами, ассоциированными с шестью R-генами устойчивости: к вирусам PVY (Rysto) и PVX (Rx1), к цистообразующим картофельным нематодам — патотипу Ro1 G. rostochiensis (ген H1) и патотипам Pa2/Ра3 G. pallida (ген Gpa2), а также с маркерами генов расоспецифической устойчивости к возбудителю фитофтороза (R1, R3а).
ПЦР осуществляли в 20 мкл реакционной смеси, имеющей следующий состав: 40 нг тотальной ДНК, 1× реакционный буфер (ЗАО «Диалат Лтд», Россия), 2,5 мM MgCl2, 0,6 мM каждого из dNTPs, 0,25 мкM прямого и обратного праймеров и 1 ед. BioTaq-ДНК-полимеразы (ЗАО «Диалат Лтд», Москва). Использовали ДНК-амплификатор Mastercycler® nexus gradient, («Eppendorf», Германия). Протоколы ПЦР и температуры отжига в целом соответствовали приведенным в литературе для каждого из использованных маркеров (10, 33-38), в ряде случаев для увеличения специфичности в протоколы дополнительно вводили функцию TouchDown.
Использовали следующие ПЦР-программы. Для маркера YES3-3A: 3 мин 30 с при 94 °С; 45 с при 94 °С, 1 мин при 60 °С с понижением температуры отжига на 1 °С за цикл, 1 мин при 72 °С (5 циклов); 40 с при 94 °С, 40 с при 55 °С, 1 мин при 72 °С (35 циклов); в заключение 10 мин при 72 °С (TouchDown) (34). Для маркера YES3-3B: 3 мин 30 с при 94 °С; 45 с при 94 °С, 45 с при 58 °С с понижением температуры отжига на 1 °С за цикл, 1 мин при 72 °С (5 циклов); 40 с при 94 °С, 40 с при 53 °С, 1 мин при 72 °С (35 циклов); в заключение 10 мин при 72 °С (TouchDown) (34). Для маркеров 5Rx1: 3 мин 30 с при 94 °С; 45 с при 94 °С, 45 с при 55 °С, 1 мин при 72 °С (35 циклов); в заключение 10 мин при 72 °С (с увеличением продолжительности этапов денатурации и отжига) (33). Для маркера 57R: 3 мин 30 с при 94 °С; 45 с при 94 °С, 1 мин при 65 °С с понижением температуры отжига на 1 °С за цикл, 1 мин при 72 °С (5 циклов); 45 с при 94 °С, 45 с при 60 °С, 45 с при 72 °С (35 циклов); в заключение 10 мин при 72 °С, (TouchDown) (35). Для маркера N195: 3 мин 30 с при 94 °С; 45 с при 94° С, 45 с при 66 °С с понижением температуры отжига на 1 °С за цикл, 1 мин 30 с при 72 °С (8 циклов); 30 с при 94 °С, 30 с при 58 °С, 1 мин 30 с при 72 °С (35 циклов); в заключение 10 мин при 72 °С (TouchDown) (10). Для маркера Gpa2-2: 4 мин 30 с при 94 °С; 30 с при 94 °С, 30 с при 60 °С, 1 мин при 72 °С (35 циклов); в заключение 10 мин при 72 °С (36). Для маркера R1: 3 мин 30 с при 94 °С; 45 с при 94 °С, 1 мин при 65 °С с понижением температуры отжига на 1 °С за цикл, 1 мин 30 с при 72 °С (10 циклов); 45 с при 94 °С, 45 с при 55 °С, 1 мин 30 с при 72 °С (30 циклов); в заключение 10 мин при 72 °С (TouchDown) (37). Для маркера R3a: 3 мин 30 с при 94 °С; 45 с 94 °С, 45 с при 68 °С с понижением температуры отжига на 1 °С за цикл, 1 мин 30 с при 72 °С (10 циклов); 45 с при 94 °С, 45 с при 58 °С, 1 мин 30 с при 72 °С (35 циклов); в заключение 10 мин при 72 °С (TouchDown) (38). Препараты ДНК родительских сортов служили положительными контролями, дистиллированная вода — отрицательным контролем.
Амплифицированные фрагменты ДНК разделяли в 2 % агарозных гелях в буфере ТВЕ, гели окрашивали бромистым этидием с последующей визуализацией в проходящем УФ-свете.
Оценка хозяйственно ценных признаков межсортовых гибридов включала учет их продуктивности, товарного вида клубней, содержания сухих веществ и крахмала в клубнях. В исследовании участвовали 144 гибридных генотипа, для каждого из них определяли показатели по 8 растениям. Продуктивность определяли весовым методом, оценивая среднюю массу клубня, число клубней с растения и среднюю массу клубней для одного гнезда. Также учитывали число товарных и нетоварных клубней, их массу с каждого куста, выравненность гнезд и число клубней в гнезде. Товарность урожая оценивали согласно методике С.А. Банадысева с соавт. (39). Содержание сухих веществ и крахмала определяли по удельной массе клубней, которую переводили в процентное содержание сухих веществ и крахмала с помощью специальной таблицы (40). Вкус клубней определяли по 9-балльной шкале, где 9 баллов — отличный, 1 — плохой (горький, неприятный) (41). Кулинарный тип определялся в соответствии с международной классификацией столовых качеств клубней картофеля: A — салатный картофель, не разваривается, B — слабо разваривается, C — хорошо разваривается, D — сильно разваривается (41). Устойчивость растений к фитофторозу оценивали на естественном фоне развития заболевания (опытное поле Ленинградского НИИ сельского хозяйства «Белогорка», Ленинградская обл.) в условиях сильного распространения инфекции в 2019 году. Использовали шкалу от 1 (целиком пораженное растение) до 9 баллов (отсутствие симптомов болезни).
Статистическую обработку данных оценки агрономических признаков проводили общепринятыми методами (42). Критерий χ2 использовали для проверки достоверности отклонений от теоретически ожидаемого расщепления в гибридных популяциях (1:1 или 5:1) при различных уровнях гетерозиготности маркированных R-локусов у носителей доминантных аллелей, участвующих в скрещиваниях.
Результаты.Сорта с маркерами разных R-генов, вовлеченные в гибридизацию, представлены в таблице 1.
Молекулярный скрининг межсортовых гибридов на наличие маркеров R-генов устойчивости к болезням и вредителям. Всего в трех комбинациях скрещиваний было выращено 144 межсортовых гибрида, 112 из них проанализировали на наличие 8 SCAR маркеров, ассоциированных с шестью R-генами — Rysto и Rx1 (устойчивость к вирусам PVY и PVX), H1 и Gpa2 (устойчивость к золотистой и бледной картофельным нематодам), R1 и R3а (расоспецифическая устойчивость к фитофторозу).
Практически у всех 112 изученных гибридных генотипов была выявлена та или иная комбинация разного числа маркеров R-генов устойчивости (рис.). Исключение составил лишь один гибрид — 7.18-7 (Гусар × Сиреневый туман), у которого мы не обнаружили ни одного из 8 использованных в MAS маркеров. Из 112 гибридных генотипов потенциальную ценность для селекции сортов с групповой устойчивостью к двум видам цистообразующих нематод ЗКН и БКН могут представлять 58 гибридов, имеющих маркеры двух генов — H1 и Gpa2, и 42 гибрида, обладающие маркерами двух генов устойчивости к вирусам PVY и PVX— Rysto и Rx1. При этом у четырех из 112 изученных гибридных сеянцев (5.18-9 и 5.18-17 —из комбинации Гусар × Чароит, 6.18-12 и 6.18-37 —из комбинации Гусар × Алый парус) мы детектировали все восемь использованных в MAS маркеров шести генов устойчивости.
У межсортовых гибридов во всех трех комбинациях отмечалось расщепление по признаку наличие/отсутствие ДНК маркеров изученных шести R-локусов (табл. 2), таким образом, среди исходных сортов квадриплексов или триплексов по маркированным R-генам мы не выявили. Расщепление можно было отнести к двум вариантам скрещиваний с разным уровнем гетерозиготности носителей доминантных аллелей: первый — симплекс Rrrr × нуллиплекс rrrrс теоретически ожидаемым расщеплением 1 R-:1 rrrr, второй вариант — дуплекс RRrr × нуллиплекс rrrrс теоретически ожидаемым расщеплением 5 R-:1 rrrr.
Анализ характера наследования ДНК маркеров в каждой комбинации (см. табл. 2) позволил выявить аллельный состав R-генов у родительских форм и, соответственно, определить степень гетерозиготности маркированных локусов исходных сортов. Так, сорт Гусар имеет один доминантный аллель (симплекс) генов Rysto и R1 и два доминантных аллеля (дуплекс) гена H1; сорт Чароит — симплекс по гену R3a и дуплексы по генам Rx1 и Gpa2-2, сорт Алый парус — симплексы генов R3a и Н1 и дуплексы по генам Gpa2 и Rx1, у сорта Сиреневый туман выявлено по одному доминантному аллелю (симплексы) для трех — R3a, Rx1 и Gpa2-2.
Агрономические признаки полученных межсортовых гибридов.Отбор лучших гибридных генотипов проводили в 2019 году по продуктивности, форме и товарному виду клубней, а также по результатам оценки полевой устойчивости к фитофторозу и к вирусам; при этом также учитывались данные MAS. Показатели хозяйственно ценных признаков определяли по 8 растениям для каждого из 144 гибридных генотипов. В результате проведенных исследований из 144 гибридов был выделен 31 перспективный гибрид, обладающий теми или иными хозяйственно ценными признаками и различным сочетанием маркеров R-генов. Из них 23 гибридных генотипа проявили относительно высокую продуктивность (табл. 3); у оставшихся 8 выделенных гибридов продуктивность оказалась низкой, но они обладали высокой полевой устойчивостью к фитофторозу, поэтому их целесообразно вовлекать в дальнейшие скрещивания. Причиной выбраковки остальных 113 гибридов были излишне длинные столоны или поражение растений фитофторозом и вирусами.
Продуктивность 23 выделенных гибридов варьировала от 600 до 1525 г/растение, что соответствует урожайности 24,5-62,2 т/га; некоторые из них выделялись и по другим хозяйственно ценным признакам (высокая товарность клубней, хороший вкус, крахмалистость) (см. табл. 3). Например, отметим гибрид 7.18-3 (Гусар × Сиреневый туман) с маркерами генов устойчивости к вирусам PVY и PVХ, а также к обоим видам нематод; этот гибрид имел высокие показатели товарности клубней и продуктивность 1350 г/растение (соответствует урожайности 55,1 т/га) (см. табл. 3).
Наивысшую продуктивность можно было наблюдать у гибрида 5.18-7 — 1525 г/растение (62,2 т/га); этот гибрид также несет имеет маркеры YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, то есть потенциально обладает комплексной устойчивостью к вирусам PVY и PVХ и к двум видам нематод — ЗКН и БКН (см. табл. 3).
Из 23 выделенных нами гибридных генотипов с высокими показателями продуктивности примерно половина (12 гибридов) характеризовались выравненностью гнезд и клубней в гнезде, а также ровной и правильной формой клубней (их характеристики приведены в таблице 4).
Использование ДНК маркеров в селекционном процессе повышает эффективность отбора целевых генотипов с доминантными аллелями разных R-генов, что особенно важно при идентификации клонов с маркерами генов устойчивости к карантинным объектам, поскольку фитопатологические оценки устойчивости растений к таким патогенам очень сложны и трудоемки (27, 43). В настоящей работе мы выявили более 40 % гибридов с маркерами двух генов (H1 и Gpa2-2) устойчивости к патотипу Ro1 золотистой и патотипам Pa2/Pa3 бледной картофельных нематод. Высокая частота таких гибридов объясняется наличием дуплексов этих генов у родительских сортов — гена H1 у сорта Гусар и гена Gpa2-2 у сортов Алый парус и Чароит. Интересно отметить, что А.П. Ермишин с соавт. (20), анализируя расщепления по ДНК маркерам в 11 гибридных популяциях, пришли к выводу о том, что у межсортовых гибридов картофеля дуплексы наиболее часто выявляются по генам устойчивости к карантинным объектам. Дополнительно у сортов Алый парус и Чароит были обнаружены дуплексы гена Rx1, который тесно сцеплен с геном Gpa2-2 (44), поэтому в гибридных комбинациях, полученных с участием этих родительских сортов, наблюдалось совместное наследование ДНК маркеров генов устойчивости к БКН и к PVX.
Итак, на основе использования комплексного подхода, сочетающего традиционные методы селекции гибридов по хозяйственно ценным признакам и методы ДНК-маркирования, были отобраны 12 гибридных генотипов, которые имеют относительно высокие показатели продуктивности, ровную и правильную форму клубней и обладают маркерами генов устойчивости к цистообразующим нематодам; у семи из них были дополнительно детектированы маркеры генов устойчивости к Y- и X-вирусам картофеля Rysto и Rx1. Отобранные гибриды представляют интерес для создания конкурентоспособных сортов, устойчивых к разным группам вредителей и возбудителей болезней (в том числе с групповой устойчивостью к цистообразующим нематодам и с групповой устойчивостью к вирусам картофеля).
1. Основные характеристики родительских форм картофеля, отобранных для межсортовых скрещиваний в 2017 году (оригинаторы Ленинградский НИИ сельского хозяйства «Белогорка», ООО Селекционная фирма «ЛиГа», Ленинградская обл.) |
||||
Признак |
Сорт Гусар |
Сорт Алый парус |
Сорт Сиреневый туман |
Сорт Чароит |
Маркеры R-генов устойчивости (26): |
|
|
|
|
к вирусам картофеля |
|
|
|
|
YES3-3A/YES3-3B (к PVY, Rysto) |
+/+ |
-/- |
-/- |
-/- |
5Rx1 (к PVX, Rx1) |
- |
+ |
+ |
+ |
к возбудителю фитофтороза |
|
|
|
|
R1 (R1) |
+ |
- |
- |
- |
RT-R3a(R3a) |
- |
+ |
+ |
+ |
к картофельным нематодам |
|
|
|
|
57R/N195 (к золотистой, H1) |
+/+ |
+/+ |
-/- |
-/- |
Gpa2-2 (к бледной, Gpa2) |
- |
+ |
+ |
+ |
Хозяйственно ценные признаки и устойчивость к болезням (фитопатологическая оценка) (29-31) |
Среднеспелый, продуктивность до 60 т/га, содержание крахмала 15-19 %, продолжительный период покоя клубней; устойчив к золотистой картофельной нематоде, к возбудителю рака картофеля, фитофторозу, парше обыкновенной, ризоктониозу |
Среднеранний, продуктивность до 50 т/га, содержание крахмала 18-23 %, устойчив к ЗКН, вирусам, возбудителю рака картофеля |
Среднеспелый, урожайность до 60 т/га, содержание крахмала 14-17%, устойчив к возбудителю рака картофеля, фитофторозу, вирусам |
Ультраранний, урожайность до 55 т/га, устойчив к возбудителю рака картофеля, ризоктониозу, парше обыкновенной |
Примечание. Маркер RT-R3aдетектирован у сорта Алый парус при использовании материала, полученного непосредственно от авторов сорта (Н.М. Гаджиев, В.А. Лебедева). У растения сорта Алый парус, полученного нами из других источников, указанный маркер ранее диагностирован не был (26). |
||||
2. Расщепление по ДНК маркерам генов устойчивости к болезням и вредителям (соотношение числа гибридов с маркером и без маркера соответствующего R-гена) у межсортовых гибридов картофеля в трех комбинациях скрещиваний с участием сорта Гусар (Ленинградский НИИ сельского хозяйства «Белогорка», Ленинградская обл., 2018 год) |
||||||
Комбинация скрещивания |
YES3-3A/YES3-3В (ген Rysto) |
5Rx1 (ген Rx1) |
57R/N195 (ген H1) |
Gpa2-2 (ген Gpa2) |
R1 (ген R1) |
RT-R3a (ген R3a) |
Гусар × Чароит |
16:11a |
22:5b |
23:4b |
22:5b |
11:16а |
10:17а |
Гусар × Алый парус |
15:15а |
22:8b |
27:3с |
21:9b |
15:15а |
14:16а |
Гусар × Сиреневый туман |
33:22а |
28:27а |
46:9b |
26:29а |
18:37 |
22:33а |
Примечание. Использованы маркеры YES3-3A/YES3-3В(Rysto — ген устойчивости к PVY), 5Rx1 (Rx1 — ген устойчивости к PVX), 57R (H1 — ген устойчивости к патотипу Ro1 золотистой картофельной нематоды), N195 (H1 — ген устойчивости к патотипу Ro1 золотистой картофельной нематоды), R1 (R1 — ген устойчивости к Phytophthora infestans) и RT-R3a(R3a — ген устойчивости к Ph. infestans). Верхние индексы: а — расщепление для указанного R-локуса в указанной комбинации при р05 соответствует теоретически ожидаемому (1:1) для скрещивания симплекс × нуллиплекс (Rrrr× rrrr); b — расщепление при р05 соответствует теоретически ожидаемому (5:1) для скрещивания дуплекс × нуллиплекс (RRrr× rrrr); с — расщепление при р05 соответствует теоретически ожидаемому (11:1) для скрещивания дуплекс × симплекс (RRrr× Rrrr). |
||||||
3. Характеристика 23 гибридных генотипов картофеля, отобранных в трех межсортовых комбинациях по маркерам R-генов устойчивости к болезням и вредителям и по хозяйственно ценным признакам (опытное поле Ленинградского НИИ сельского хозяйства «Белогорка», Ленинградская обл., 2019 год; изучено по 8 растений каждого гибрида) |
|||||||
Сорт, гибрид |
Продуктивность, г/куст (min-max) |
Товарность, % |
Содержание в клубнях, % |
Вкус клубней, балл |
Кулинарный тип |
Наличие ДНК маркеров генов устойчивости к вирусам Y и Х картофеля, фитофторозу, золотистой картофельной нематоде и бледной картофельной нематоде |
|
крахмала |
сухого вещества |
||||||
Невский (st) |
700 (560-810) |
92 |
14,2 |
20,0 |
5,5 |
А |
RT-R3a |
Десять гибридов комбинации Гусар × Чароит |
|||||||
5.18-3 |
787 (690-855) |
97 |
15,4 |
21,1 |
7,5 |
B |
5Rx1, N195, 57R, Gpa2-2, RT-R3a |
5.18-7вп |
1525 (1300-1740) |
97 |
10,0 |
15,0 |
7,0 |
А |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2 |
5.18-15 |
643 (590-735) |
98 |
12,9 |
18,6 |
8,0 |
AB |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, R1 |
5.18-17 |
600 (430-810) |
95 |
14,2 |
20,0 |
7,0 |
A |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, R1, RT-R3a |
5.18-19вп |
925 (780-1060) |
97 |
11,7 |
17,4 |
5,0 |
A |
YES3-3А/3B, 5Rx1, Gpa2-2, R1 |
5.18-24вп |
1253 (1010-1470) |
94 |
14,7 |
20,4 |
6,0 |
A |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, RT-R3a |
5.18-25 |
860 (620-910) |
99 |
11,7 |
17,4 |
5,5 |
A |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, R1 |
5.18-27 |
800 (680-865) |
91 |
13,4 |
19,1 |
6,7 |
A |
Нет данных |
5.18-32 |
630 (520-700) |
86 |
14,4 |
20,1 |
6,0 |
A |
YES3-3А/3B, 5Rx1, Gpa2-2, R1 |
5.18-34вп |
1340 (1100-1480) |
94 |
15,7 |
21,5 |
5,5 |
A |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, |
Восемь гибридов комбинации Гусар × Алый парус |
|||||||
6.18-2 |
685 (583-770) |
99 |
14,2 |
20,0 |
7,5 |
A |
5Rx1, N195, 57R, Gpa2-2, R1, RT-R3a |
6.18-9 |
735 (615-842) |
95 |
15,2 |
21,0 |
6,5 |
A |
YES3-3А/3B, N195, 57R |
6.18-13вп |
1340 (1200-1420) |
94 |
12,9 |
18,6 |
5,5 |
A |
5Rx1, Gpa2-2 |
6.18-17 |
770 (630-865) |
97 |
11,0 |
16,7 |
6,5 |
A |
5Rx1, N195, 57R, Gpa2-2 |
6.18-24вп |
1015 (810-1360) |
94 |
11,0 |
16,7 |
6,0 |
A |
YES3-3А/3B, N195, 57R, RT-R3a |
6.18-26вп |
1155 (900-1320) |
88 |
14,2 |
20,0 |
6,0 |
A |
YES3-3А/3B, N195, 57R, R1 |
6.18-36вп |
870 (820-945) |
93 |
13,4 |
19,1 |
7,5 |
AB |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, |
6.18-42 |
765 (630-880) |
96 |
14,7 |
20,5 |
6,0 |
A |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, RT-R3a |
Пять гибридов комбинации Гусар × Сиреневый туман |
|||||||
7.18-3вп |
1350 (1210-1545) |
96 |
11,0 |
16,8 |
5,5 |
A |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, RT-R3a |
7.18-4 |
780 (660-863) |
97 |
12,9 |
18,6 |
6,5 |
AB |
YES3-3А/3B, R1, RT-R3a |
7.18-42вп |
1010 (820-1220) |
92 |
12,2 |
18,0 |
6,0 |
A |
5Rx1, N195, 57R |
7.18-55вп |
1255 (960-1320) |
96 |
13,4 |
19,1 |
8,0 |
B |
N195, 57R, R1, RT-R3a |
7.18-56вп |
1280 (1000-1380) |
91 |
12,2 |
18,0 |
7,0 |
AB |
YES3-3А/3B, 5Rx1, Gpa2-2, R1, RT-R3a |
Примечание. Сорт Невский (st) используется в Ленинградском НИИ сельского хозяйства «Белогорка» в качестве стандарта при оценке агрономических признаков. У гибрида 5.18-17 детектированы все изученные маркеры генов устойчивости; вп — 12 гибридных генотипов с высокой продуктивностью (их дополнительные характеристики приведены в таблице 4). |
|||||||
4. Некоторые хозяйственно значимые признаки 12 гибридов, выделившихся по продуктивности (n = 8, опытное поле Ленинградского НИИ сельского хозяйства «Белогорка», Ленинградская обл., 2019 год) |
||||||
Сорт, гибрид |
Наличие ДНК-маркеров R-генов устойчивости |
Окраска кожуры клубней |
Форма клубней |
Глубина и окраска глазков |
Выравненность, балл (от 9 до 1) |
|
гнезд |
клубней в гнезде |
|||||
Невский (st) |
RT-R3a |
Белая |
Овальная |
Мелкие, малиновые |
8 |
8 |
5.18-7 |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2 |
Белая |
Овальная |
Мелкие |
7 |
8 |
5.18-19 |
YES3-3А/3B, 5Rx1, Gpa2-2, R1 |
Белая |
удлиненная |
Мелкие |
7 |
7 |
5.18-24 |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, RT-R3a |
Желтая |
Овальная |
Мелкие |
8 |
8 |
5.18-34 |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, |
Желтая |
Овальная |
Мелкие |
8 |
8 |
6.18-13 |
5Rx1, Gpa2-2 |
Розовая |
Удлиненно-овальная |
Мелкие, малиновые |
8 |
8 |
6.18-24 |
YES3-3А/3B, N195, 57R, RT-R3a |
Белая |
Удлиненно-овальная |
Мелкие |
8 |
8 |
6.18-26 |
YES3-3А/3B, N195, 57R, R1 |
Розовая |
Овальная |
Мелкие, малиновые |
8 |
8 |
6.18-36 |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, |
Белая |
Овальная |
Мелкие |
7 |
8 |
7.18-3 |
YES3-3А/3B, 5Rx1, N195, 57R, Gpa2-2, RT-R3a |
Белая |
Округло-овальная |
Мелкие |
8 |
8 |
7.18-42 |
5Rx1, N195, 57R |
Розовая |
Овальная |
Мелкие, малиновые |
8 |
8 |
7.18-55 |
N195, 57R, R1, RT-R3a |
Розовая |
Овальная |
Мелкие |
8 |
8 |
7.18-56 |
YES3-3А/3B, 5Rx1, Gpa2-2, R1, RT-R3a |
Желтая |
Удлиненная |
Мелкие |
8 |
7 |
Примечание. Сорт Невский (st) используется в Ленинградском НИИ сельского хозяйства «Белогорка» в качестве стандарта при оценке агрономических признаков. |
||||||
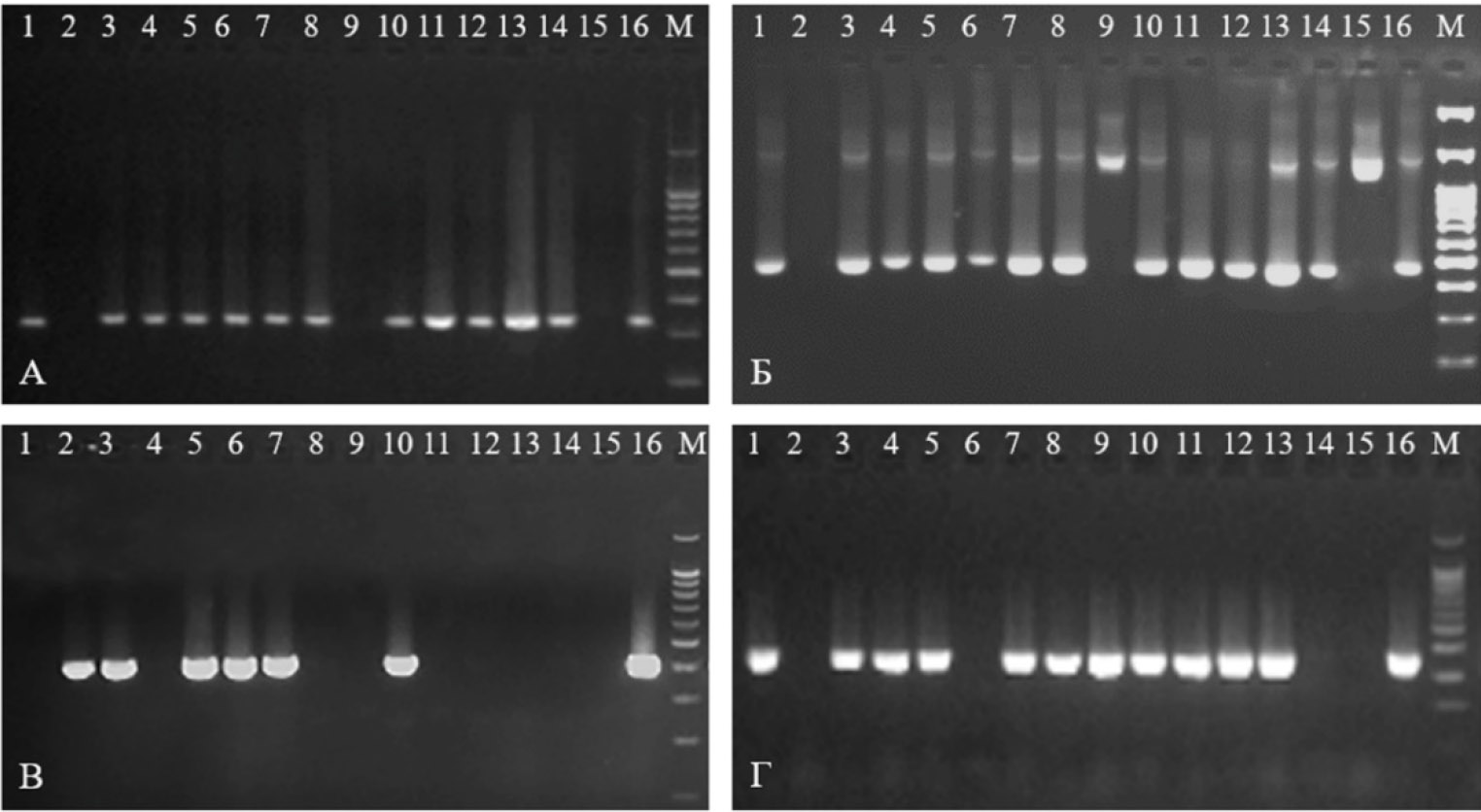
Скрининг межсортовых гибридов картофеля (комбинация сортов Гусар × Сиреневый туман) на наличие ДНК маркеров генов устойчивости к золотистой картофельной нематоде N195 (А) и 57R (Б), к бледной картофельной нематоде Globodera pallida Gpa2-2 (В) и Y-вирусу картофеля (PVY) YES3-3a (Г): 1 — Гусар, 2 — Сиреневый туман, 3 — гибрид 7.18-1, 4 — гибрид 7.18-8, 5 — гибрид 7.18-2, 6 — гибрид 7.18-9, 7 — гибрид 7.18-3, 8 — гибрид 7.18-10, 9 — гибрид 7.18-4, 10 — гибрид 7.18-11, 11 — гибрид 7.18-5, 12 — гибрид 7.18-12, 13 — гибрид 7.18-6, 14 — гибрид 7.18-13, 15 — гибрид 7.18-7, 16 — гибрид 7.18-14; M — маркер молекулярных масс 100 bp + 1500 + 3000 (НПО «СибЭнзим», Россия).
ЛИТЕРАТУРА
- Зыкин В.А. Системный анализ проблемы подбора пар для гибридизации. В сб.: Селекция и семеноводство сельскохозяйственных культур в Западной Сибири. Новосибирск, 1984: 3-12.
- Сюков В.В. Методы подбора родительских пар для гибридизации у самоопыляющихся растений. Самара, 2014.
- Лепехов С.Б. Методы подбора пар для скрещивания в селекции на урожайность у самоопыляющихся культур. Труды по прикладной ботанике, генетике и селекции, 2017, 178(4): 76-89 (doi: 10.30901/2227-8834-2017-4-76-89).
- Simko I., Jansky S., Stephenson S., Spooner D. Genetics of resistance to pests and disease. In: Potato biology and biotechnology: advances and perspectives /D. Vreugdenhil, J. Bradshaw, C. Gebhardt, F. Govers, M. Taylor, D. MacKerron, H. Ross (eds.). Elsevier, St. Louis, MO, 2007: 117-155.
- Gebhardt C. Bridging the gap between genome analysis and precision breeding in potato. Trends Genet., 2013, 29(4): 248-256 (doi: 10.1016/j.tig.2012.11.006).
- Milczarek D., Przetakiewicz A., Kamiński P., Flis B. Early selection of potato clones with the H1 resistance gene — the relation of nematode resistance to quality characteristics. Czech J. Genet. Plant Breed., 2014, 50(4): 278-284 (doi: 10.17221/114/2014-CJGPB).
- Ottoman R.J., Hane D.C., Brown C.R., Yilma S., James S.R., Mosley A.R., James M.C., Vales M.I. Validation and implementation of marker-assisted selection (MAS) for PVY resistance (Ryadg gene) in a tetraploid potato breeding program. American Journal of Potato Research, 2009, 86: 304-314 (doi: 10.1007/s12230-009-9084-0).
- Nie X., Chen H., Zhang J., Zhang Y., Yang J., Pan H., Song W.X., Murad F., He Y.Q. Bian K. Rutaecarpine ameliorates hyperlipidemia and hyperglycemia in fat-fed, streptozotocin-treated rats via regulating the IRS-1/PI3K/Akt and AMPK/ACC2 signaling pathways. Acta Pharmacologica Sinica, 2016, 37(4): 483-496 (doi: 10.1038/aps.2015.167).
- Gebhardt C., Bellin D., Henselewski H., Lehmann W., Schwarzfischer J., Valkonen J.P.T. Marker-assisted combination of major genes for pathogen resistance in potato. Theoretical and Applied Genetics, 2006, 112: 1458-1464 (doi: 10.1007/s00122-006-0248-8).
- Mori K., Sakamoto Y., Mukojima N., Tamiya S., Nakao T., Ishii T., Hosaka K. Development of a multiplex PCR method for simultaneous detection of diagnostic DNA markers of five disease and pest resistance genes in potato. Euphytica, 2011, 180: 347-355 (doi: 10.1007/s10681-011-0381-6).
- Mori K., Asano K., Tamiya S., Nakao T., Mori M. Challenges of breeding potato cultivars to grow in various environments and to meet different demands. Breeding Science, 2015, 65: 3-16 (doi: 10.1270/jsbbs.65.3).
- Asano K., Tamiya S. Breeding of pest and disease resistant potato cultivars in Japan by using classical and molecular approaches. The Japan Agricultural Research Quarterly, 2016, 50(1): 1-6 (doi: 10.6090/jarq.50.1).
- Milczarek D., Plich J., Tatarowska B., Flis B. Early selection of potato clones with resistance genes: the relationship between combined resistance and agronomical characteristics. Breeding Science, 2017, 67: 416-420 (doi: 10.1270/jsbbs.17035).
- Зотеева Н.М., Антонова О.Ю., Клименко Н.С., Апаликова О.В., Carlson-Nilsson U., Карабицина Ю.И., Ухатова Ю.В., Гавриленко Т.А. Использование молекулярных маркеров R генов и типов цитоплазмы при интрогрессивной гибридизации диких полиплоидных мексиканских видов картофеля. Сельскохозяйственная биология, 2017, 52(5): 964-975 (doi: 10.15389/agrobiology.2017.5.964rus).
- Сайнакова А.Б., Романова М.С., Красников С.Н., Литвинчук О.В., Алексеев Я.И., Никулин А.В., Терентьева Е.В. Исследование коллекционных образцов картофеля на наличие генетических маркеров устойчивости к фитопатогенам. Вавиловский журнал генетики и селекции, 2018, 22(1): 18-24 (doi: 10.18699/VJ18.326).
- Рогозина Е.В., Терентьева Е.В., Потокина Е.К., Юркина Е.Н., Никулин А.В., Алексеев Я.И. Идентификация родительских форм для селекции картофеля, устойчивого к болезням и вредителям, методом мультиплексного ПЦР-анализа. Сельскохозяйственная биология, 2019, 54(1): 19-30 (doi: 10.15389/agrobiology.2019.1.19rus).
- Bradshaw J.E., Mackay G.R. Breeding strategies for clonally propagated potatoes. In: Potato genetics /J.E. Bradshaw, G.R. Mackay (eds.). CABI, Wallingford (UK), 1994: 467-497.
- Wu R., Gallo-Meagher M., Littell R.C., Zeng Z.-B. A general polyploid model for analyzing gene segregation in outcrossing tetraploid species. Genetics, 2001, 159(2): 869-882.
- Skupinova S., Vejl P., Sedlak P., Domkarova J. Segregation of DNA markers of potato (Solanum tuberosum ssp. tuberosum L.) resistance against Ro1 pathotype Globodera rostochiensis in selected F1 progeny. Rostlinna vyroba, 2002, 48(11): 480-485.
- Ермишин А.П., Свиточ О.В., Воронкова Е.В., Гукасян О.Н., Лукша В.И. Определение состава и аллельного состояния генов устойчивости к болезням и вредителям у родительских линий картофеля с помощью ДНК маркеров. Генетика, 2016, 52(5): 569-578 (doi: 10.7868/S0016675816050052).
- Kneib R., Kneib R., da Silva Pereira A., Castro C.M. Allele dosage of PVY resistance genes in potato clones using molecular markers. Crop Breeding and Applied Biotechnology, 2017, 17: 306-312 (doi: 10.1590/1984-70332017v17n4a47).
- Лебедева Н.А. Полиплоидия некоторых видов картофеля. Труды МОИП, отд. биол., 1962, 5: 215-221.
- Лебедева Н.А. Получение и использование полиплоидов в селекции картофеля. Тезисы докл. 2-го совещания по полиплоидии. Л., 1963: 28-29.
- Лебедева Н.А. Экспериментальная полиплоидия в селекции. Картофель и овощи, 1965, 4: 20-24.
- Антонова О.Ю., Швачко Н.А., Новикова Л.Ю., Шувалов О.Ю., Костина Л.И., Клименко Н.С., Шувалова А.Р., Гавриленко Т.А. Генетическое разнообразие сортов картофеля российской селекции и стран ближнего зарубежья по данным полиморфизма SSR-локусов и маркеров R-генов устойчивости. Вавиловский журнал генетики и селекции, 2016, 20(5): 596-606 (doi: 10.18699/VJ16.181).
- Гавриленко Т.А., Клименко Н.С., Антонова О.Ю., Лебедева В.А., Евдокимова З.З., Гаджиев Н.М., Апаликова О.В., Алпатьева Н.В., Костина Л.И., Зотеева Н.М., Мамадбокирова Ф.Т., Егорова К.В. Молекулярный скрининг сортов и гибридов картофеля северо-западной зоны Российской Федерации. Вавиловский журнал генетики и селекции, 2018, 22(1): 35-45 (doi: 10.18699/VJ18.329).
- Хютти А.В., Антонова О.Ю., Мироненко Н.В., Гавриленко Т.А., Афанасенко О.С. Устойчивость картофеля к карантинным болезням. Вавиловский журнал генетики и селекции, 2017, 21(1): 51-61 (doi: 10.18699/VJ17.223).
- Гавриленко Т.А., Клименко Н.С., Алпатьева Н.В., Костина Л.И., Лебедева В.А., Евдокимова З.З., Апаликова О.В., Новикова Л.Ю., Антонова О.Ю. Генетическое разнообразие сортов картофеля российской селекции и стран ближнего зарубежья по типам цитоплазм. Вавиловский журнал генетики и селекции, 2019, 23(6): 753-764 (doi: 10.18699/VJ19.534).
- Лебедева В.А. Селекция картофеля на основе межвидовой гибридизации. СПб, 2010.
- Лебедева В.А., Гаджиев Н.М. Два урожая сорта картофеля Чароит за один сезон. Мат. Международного конгресса «Агрорус». СПб, 2014: 19-20.
- Гаджиев Н.М., Лебедева В.А. Новый сорт картофеля Гусар. Защита картофеля, 2015, 2: 16.
- Gavrilenko T., Antonova O., Shuvalova A., Krylova E., Alpatyeva N., Spooner D.M., Novikova L. Genetic diversity and origin of cultivated potatoes based on plastid microsatellite polymorphism. Genetic Resources and Crop Evolution, 2013, 60(7): 1997-2015 (doi: 10.1007/s10722-013-9968-1).
- Ahmadvand R., Wolf I., Gorji A.M., Polgár Z., Taller J. Development of molecular tools for distinguishing between the highly similar Rx1 and Rx2 PVX extreme resistance genes in tetraploid potato. Potato Research, 2013, 56(4): 277-291 (doi: 10.1007/s11540-013-9244-y).
- Song Y.-S., Schwarzfischer A. Development of STS markers for selection of extreme resistance (Rysto) to PVY and maternal pedigree analysis of extremely resistant cultivars. American Journal of Potato Research, 2008, 85(2): 159-170 (doi: 10.1007/s12230-008-9012-8).
- Schultz L., Cogan N.О.I., Mclean K., Dale M.F.B., Bryan G.J., Forster J.N.W., Slater A.T. Evaluation and implementation of a potential diagnostic molecular marker for H1-conferred potato cyst nematode resistance in potato. Plant Breeding, 2012, 131: 315-321 (doi: 10.1111/j.1439-0523.2012.01949.x).
- Asano K., Kobayashi A., Tsuda S., Nishinaka M., Tamiya S. DNA marker-assisted evaluation of potato genotypes for potential resistance to potato cyst nematode pathotypes not yet invading into Japan. Breeding Science, 2012, 62(2): 142-150 (doi: 10.1270/jsbbs.62.142).
- Ballvora A., Ercolano M.R., Weiss J., Meksem K., Bormann C.A., Oberhagemann P., Salamini F., Gebhardt C. The R1 gene for potato resistance to late blight (Phytophthora infestans) belongs to the leucine zipper/NBS/LRR class of plant resistance genes. The Plant Journal, 2002, 30(3): 361-371 (doi: 10.1046/j.1365-313X.2001.01292.x).
- Huang S., van der Vossen E.A.G., Kuang H., Vleeshouwers V.G., Zhang N., Borm T.J.A., van Eck H.J., Baker B., Jacobsen E., Visser R.G.F. Comparative genomics enabled the isolation of the R3a late blight resistance gene in potato. The Plant Journal, 2005, 42(2): 251-261 (doi: 10.1111/j.1365-313X.2005.02365.x).
- Банадысев С.А., Старовойтов А.М., Колядко И.И., Маханько В.Л., Фандо В.В., Козлова Л.И., Колядко О.М., Незаконова Л.В., Гончарова Н.Н., Вологдина Л.Н., Стадников И.А., Грибко А.П. Методические рекомендации по специализированной оценке сортов картофеля. Минск, 2003.
- Симаков Е.А., Склярова Н.П., Яшина И.М. Методические указания по технологии селекционного процесса картофеля. М., 2006.
- Симаков Е.А. Анисимов Б.В., Шабанов А.Э., Зебрин С.Н., Юрлова С.М., Овэс Е.В., Зейрук В.Н., Усков А.И., Федотова Л.С., Филиппова Г.И. Методические положения по проведению оценки сортов картофеля на испытательных (тестовых) участках. М., 2013.
- Рокицкий П.Ф. Биологическая статистика. М., 1973.
- Dalamu V.B., Umamaheshwari R., Sharma R., Kaushik S., Joseph T., Singh B., Gebhardt C. Potato cyst nematode (PCN) resistance: genes, genotypes and markers. SABRAO Journal of Breeding and Genetics, 2012, 44(2): 202-228.
- Van der Vossen E.A.G., van der Voort J.R., Kanyuka K., Bendahmane A., Sandbrink H., Baulcombe D.C., Bakker J., Stiekema W.J., Klein-Lankhorst R.M. Homologues of a single resistance-gene cluster in potato confer resistance to distinct pathogens: a virus and a nematode. The Plant Juornal, 2000, 23(5): 567-576 (doi: 10.1046/j.1365-313x.2000.00814.x).