doi: 10.15389/agrobiology.2020.5.1018rus
УДК 631.4:579:577.2
При выполнении работы было использовано оборудование ресурсного центра «Методы изучения состава вещества» Научного парка СПбГУ. Исследование проведено при финансовой поддержке РНФ (проект № 19-16-00049).
ПОЧВЫ ЧЕРНЕВОЙ ТАЙГИ ЗАПАДНОЙ СИБИРИ — МОРФОЛОГИЯ, АГРОХИМИЧЕСКИЕ ОСОБЕННОСТИ, МИКРОБИОТА
Е.В. АБАКУМОВ1 ✉, С.В. ЛОЙКО2, 3, Г.И. ИСТИГЕЧЕВ2, А.И. КУЛЕМЗИНА4, Н.Н. ЛАЩИНСКИЙ5, Е.Е. АНДРОНОВ6, 7,
А.Л. ЛАПИДУС1, 8 ✉
Почвы черневой тайги Западной Сибири обладают исключительно высоким потенциальным плодородием, не затронутым негативными последствиями многолетней сельскохозяйственной практики и реализующимся за счет внутренних биотических и абиотических ресурсов. Анализ такого комплекса связанных свойств в рамках метагеномного исследования и выявление микробных драйверов плодородия могут стать основой инновационных технологий повышения продуктивности почв и сельскохозяйственных культур. В настоящей работе нами впервые получены данные о таксономической структуре и особенностях микробиоты почв черневой тайги и выявлены таксоны, численность которых достоверно увеличивается при переходе от фоновой зональной почвы к почве черневой тайги. Анализ почвенных образцов, собранных при экспедиционных обследованиях в 2019 году, показал, что почвы черневой тайги (Новосибирская, Томская и Кемеровская области, Алтайский край) представлены в основном подзолистыми, дерново-подзолистыми, серыми и (намного реже) темно-серыми почвами (тяжелосуглинистыми и легкоглинистыми разновидностями, приуроченными к делювиальным чехлам голоценового и позднеплейстоценового возраста). Эти почвы формируются при уникальных комбинациях геогенных и биоклиматических условий, при которых почвы не промерзают зимой и обеспечены влагой, что способствует быстрой минерализации подстилочного материала и закреплению минеральных элементов питания в верхней гумусированной толще почвенного профиля. Аккумуляция биофильных элементов является важнейшим свойством почв черневой тайги, с которым связан феномен гигантизма и крайне высокой продуктивности растений. Мы показали, что почвы черневой тайги содержат максимальное количество органических углерода по сравнению с почвами олиготрофных местообитаний (9,85 против 2,74 %). Актуальное почвенное плодородия в черневой тайге в несколько раз выше, чем в смежных биотопах (максимальное содержание доступных форм фосфора и калия — соответственно 702 и 470 мг/кг). Расположенные в смежных ландшафтах почвы относительно олиготрофных лесов бедны по агрохимическим показателям плодородия (максимальное содержание обменных форм фосфора и калия — соответственно 113 и 18 мг/кг), имеют маломощный гумусовый горизонт и относятся (согласно субстантивно-профильной классификации почв России) к органо-аккумулятивным и альфегумусовым стволам почвообразования. Разнообразие микроорганизмов в изученных почвах различается в зависимости от режима трофности экосистемы. Почвы черневой тайги характеризуются повышенным разнообразием микробного сообщества (оценка по индексу Шеннона), а также присутствием филумов Nitrospirae и Thaumarchaeota,которые, однако, не доминируют. Общие филы для всех изученных почв — Proteobacteria, Verrucomicrobia, Actinobacteria, Acidobacteria, Planctomycetes и Firmicutes.
Ключевые слова: экологические функции почв, черневая тайга, микробные сообщества, метагеномное секвенирование, факторы плодородия, Западная Сибирь.
Глобальное доминирование человечества в биосфере определяется прежде всего развитием сельского хозяйства, охватившего более половины суши Земли. Это приводит к кардинальным изменениям в окружающей среде и климате (1), к нехватке продовольствия и энергии, потере биоразнообразия и устойчивости экосистем, вызывает загрязнение природных вод (2-5). Более 60 % всех изменений земельных угодий в период с 1982 по 2016 год связано с деятельностью человека, что указывает на управляющую роль человечества в развитии Земли (6). Проблемы, вызванные антропогенной трансформацией почвенного покрова и экологических функций почв, относят к наиболее важным в текущем столетии (7). Современное человечество обязано заниматься сохранением и защитой почв, поскольку почвенные ресурсы составляют основу продовольственной и экологической безопасности наций. Длительное использование почв в сельскохозяйственном производстве имеет множество негативных последствий для почвенных свойств, включая деградацию органического вещества (в том числе дегумификацию), увеличение эмиссии парниковых газов и диверсификацию ее продуктов эмиссии), изменение кислотно-основного состава почвы, истощение почвы и т.д. (8-10). Применение агротехнологий (вспашка и рыхление, внесение минеральных и органических удобрений и т.д.) значительно изменяет биогеохимические циклы, в том числе те, в которых участвуют почвенные микроорганизмы. Общий тренд этих изменений проявляется в снижении способности почвы обеспечивать сельскохозяйственные растения необходимым количеством элементов минерального питания и азотом. В связи с этим агроэкосистема не способна эффективно функционировать без внесения дополнительных доз питательных веществ в виде удобрений (10).
Внесение удобрений в агроэкосистему сопровождается появлением новых проблем, а именно существенным выносом элементов из агроэкосистемы с поверхностным и внутрипочвенным стоком воды (11-13). Более того, со временем отдача агрокультуры от внесения удобрений уменьшается (14), а при увеличении доз внесенного азота снижается эффективность его удержания агроэкосистемой (13). Эти неизбежные процессы можно замедлить, если создать в почве достаточный пул микробной биомассы, способной аккумулировать элементы минерального питания и азот органического происхождения в периоды между сбором урожая и посадкой агрокультуры. Более того, известны биогеохимические механизмы, которые позволяют удерживать в экосистемах часть внесенного с удобрениями азота на протяжении десятилетий даже в условиях существующей сельскохозяйственной практики (15). Это значит, что проблему, вызванную снижением эффективности удобрений при их длительном применении, помогут решить технологии, способствующие поддержанию и реализации потенциала почвенного плодородия.
По нашему мнению, необходимый пул микробной биомассы можно сформировать бионически, используя в качестве основы модели природных микробиот, способных в гумидных автономных ландшафтах поддерживать максимально высокую биологическую продуктивность растительности за счет достаточной замкнутости биологического круговорота веществ в системе. В условиях промывного водного режима это обеспечивается сохранением объема круговорота элементов минерального питания. Состав такой микробиоты должен складываться эволюционно при постепенном увеличении объема биологического круговорота за счет постоянного экзогенного поступления элементов питания в случае их выветривания и азота в процессе азотфиксации. К факторам, которые нарушают эти события, «размыкая» эволюционные циклы, относятся глобальные природные и техногенные катаклизмы, катастрофические пожары и распашка. Использование микробиоты могло бы стать одним из направлений биологической мелиорации почв, что крайне актуально для агроландшафтов России (16).
Почвы черневой тайги — наглядный пример проявления ключевых экологических функций эдафотопа, связанных с плодородием и лесорастительными свойствами. Его изучение важно для понимания феномена высокого плодородия почв. Уникальное сочетание факторов почвообразования в черневой тайге запускает драйверы интенсивного почвообразования и круговорота веществ. Отметим, что в настоящее время подобных экосистем в умеренном климате практически не осталось. Как правило, лесные экосистемы Европы либо находятся на месте бывших сельскохозяйственных угодий, либо располагались на их периферии и испытали мощное влияние пирогенного фактора (17). Первичные ненарушенные экосистемы, представляющие собой чрезвычайно продуктивные и максимально биогеохимически интенсивные биокосные образования, стоит искать в пределах барьерно-дождевых ландшафтов Сибири.
Основной ареал черневой тайги расположен в диапазоне высот приблизительно от 200 до 700-800 м на западных, наветренных макросклонах гор и предгорий юга Западной Сибири. Черневая тайга относится к типу барьерно-дождевых ландшафтов и характеризуется комплексом признаков (26-31): в древостое доминирует пихта и осина; хорошо развит травяной ярус, образованный видами сибирского высокотравья; в подлеске присутствуют крупные кустарники; слабо представлена синузия напочвенных листостебельных мхов, эпифитная бриофлора довольно богата; во флоре представлен комплекс относительно теплолюбивых неморальных видов; мощно развита весенняя синузии эфемероидов; зимой формируется глубокий снежный покров (от 80 см до 2 м и более), благодаря которому почвы не промерзают, что не характерно для континентального климата тайги Западной Сибири; отсутствуют стратифицированные подстилки, время разложения опада составляет менее 2 лет; численность дождевых червей одна из самых высоких в России, в целом высока активность мезофауны, в том числе в зимнее время. Благодаря перечисленным факторам и почвенным особенностям черневой тайги ее биопродуктивность по сравнению с другими зональными типами экосистем умеренного пояса максимальна.
Высокотравная черневая тайга предгорий и гор юга Сибири — один из крупнейших сохранившихся массивов высокотравных лесов России. Высокотравные леса также типичны для Дальнего Востока. В целом в пределах таежной зоны высокотравные леса сохранились в виде изолированных участков, например в Предуралье и на западных склонах Урала, на равнинах Западной Сибири, в местах, наименее подвергшихся пожарам и воздействиям традиционного природопользования (21, 22). Наименее нарушенные участки высокотравных лесов соответствуют определению климаксных экосистем как по фитоценотическим, так и по почвенным критериям. Ежегодно поступающий опад высокотравья быстро разлагается, что с течением времени приводит к накоплению элементов минерального питания в гумусовом горизонте (18) и поддерживает активный биологический круговорот веществ, гетерогенность внутриценотической среды и высокое биоразнообразие. Закреплению элементов питания растений в почвенном профиле, вероятно, способствует водный и климатический режим и особенности гидрофизических характеристик соответствующих почв.
Выбор высокотравных лесов в качестве бионической модели связан с тем, что они наименее других подвергались в прошлом экзогенным нарушениям (19, 20). В биогеографических исследованиях показано, что экзогенные нарушения приводят к упрощению структуры (пространственной, видовой) сообществ (23). Существует точка зрения, что современные зональные экосистемы южной, средней и северной тайги представляют собой сукцессионные ряды восстановления растительности после экзогенных нарушений — пожаров, рубок, распашки, то есть в ходе демутационных смен (20). Почти все эти леса подстилочные, с преобладанием зеленых мхов, мелкотравных и кустарничковых видов. Среди них лишь бореально-неморально-высокотравные леса рассматриваются некоторыми авторами как относительно полно отвечающие заключительному этапу аутогенной сукцессии (24).
Приведенная точка зрения на природу бореальных лесов дискуссионная, однако она хорошо подчеркивает обособленность высокотравных лесов. Несмотря на название, черневая тайга в действительности существенно отличается от собственно таежных, бореальных экосистем по видовому и ценотическому составу, трофности и биогеохимии. Черневая тайга большинством исследователей рассматривается как субнеморальная, или гемибореальная, экосистема, то есть относится к другому классу экосистем, чем бореальный лес (собственно тайга) (25).
Уникальность почв черневой тайги состоит в исключительно высоком плодородии, реализованном за счет внутренних биотических и абиотических ресурсов, и сохранении микробиоты, не затронутой сельскохозяйственной практики. Почвы черневой тайги демонстрируют ряд особенностей, в частности высокую лесорастительную активность при общем низком накоплении гумуса (эффективное плодородие) (26), необычно высокую скорость разложения растительных остатков (27). Анализ такого комплекса связанных свойств в рамках метагеномного исследования представляет собой серьезную фундаментальную задачу. Итогом ее решения и выявления микробных драйверов плодородия могут стать инновационные технологии повышения продуктивности почв и сельскохозяйственных культур.
Для изучения почвенного микробиоты используется высокопроизводительное секвенирование (28, 29), позволяющее с недоступной ранее точностью выявлять компоненты микробных сообществ, в том числе некультивируемые. Стало понятно, что почвенная микробиота (в особенности ризосферная) играет большую роль в питании растений и их защите от биотических и абиотических стрессов, вследствие чего в настоящее время особое внимание уделяется анализу ризосферных сообществ микроорганизмов (30).
Очевидно, что феномен высокого плодородия почв черневой тайги не может ограничиваться только агрохимическими и агрофизическими параметрами и должен быть связан с особенностями почвенной микробиоты. Можно также ожидать, что исследования этого потенциального источника новых экономически значимых штаммов (например, целлюлозолитиков, продуцентов антибиотиков и разнообразных биологически активных молекул), характерных для изучаемой экосистемы, выйдут за пределы почвенной и сельскохозяйственной микробиологии. Однако какие-либо данные о микробиоте почв черневой тайги до настоящего времени отсутствовали.
В этом сообщении впервые охарактеризована таксономическая структура прокариотической микробиоты почвы черневой тайги. Показано, что различия между фоновой зональной почвой и почвой черневой тайги скорее всего определяются на уровнях ниже филумов. Выявлены таксоны, численность которых достоверно увеличивается в почве черневой тайги. В основном это неклассифицируемые прокариоты. Среди идентифицируемых микроорганизмов особый интерес вызывает порядок Chthoniobacterales, первый представитель которого выделен лишь недавно.
Нашей целью было определение основных морфологических, агрохимических особенностей и таксономического состава микробиоты почв на участках черневой тайги в Западной Сибири в сравнении с почвами, приуроченными к олиготрофным экосистемам сосновых лесов на песчаных почвообразующих субстратах.
Методика. Во второй половине июля 2019 года проводились экспедиционные обследования почв в Томской, Кемеровской и Новосибирской областях, а также в Алтайском крае. В качестве объектов исследования выбрали четыре почвенных профиля (разные варианты черневой тайги в Салаирском и Кузнецко-Алатаусском ареалах), а также две почвы из относительно олиготрофных местообитаний, приуроченных к древнедюнным эоловым песчаным массивам, покрытым сосновыми лесами. Южные варианты исследованных почв были представлены следующими объектами: N1 (темно-серая почва черневой тайги, Алтайский край; 54.14070° с.ш., 84.9495° в.д.), N2 (стратозем темногумусовый, травяной сосновый лес, эрозионная долина, Новосибирская обл.; 54.37083° с.ш., 82.4393° в.д.), N3 (серогумусовая почва на эоловых супесях под олиготрофной боровой экосистемой, Новосибирская обл.; 54.40810° с.ш., 82.18420° в.д.), северные варианты — T1 (темно-серая почва в высокотравном пихтово-осиновом лесу с усыхающим пихтовым древостоем, Томская обл.; 56.30693° с.ш., 85.47063° в.д.), T2 (дерново-подзолистая почва, высокотравно-широкотравный березово-пихтовый послерубочный черневой лес, Кемеровская обл.; 55.88619° с.ш., 86.00433° в.д.), T3 (дерново-элювозем супесчаный под олиготрофной экосистемой сосняка разнотравного с примесью лиственницы, Томская обл., координаты: 56.48106° с.ш., 84.79860° в.д.).
Гранулометрический состав анализировали по Качинскому с пирофосфатной пептизацией микроагрегатов (седиментационный метод). Содержание органического углерода и азота определяли на элементном анализаторе (EURO EA-3028-HT, «EuroVector S.p.A.», Италия; ресурсный центр «Методы изучения состава вещества» Научного парка СПбГУ). рН водной вытяжки измеряли при соотношении 1:2,5, когда рН водной суспензии был ниже 7,0, при том же соотношении измеряли рН солевой суспензии. Подвижные соединения фосфора и калия определяли по Кирсанову в модификации ЦИНАО (ГОСТ Р 54650-2011. Национальный стандарт Российской Федерации. Почвы), обменный аммоний — по методу ЦИНАО (ГОСТ 26489-85. Государственный стандарт СССР. Почвы), нитраты — ионометрически (ГОСТ 26951-86. Государственный стандарт СССР. Почвы).
Базальное дыхание (БД) как показатель биологической метаболической активности почв измеряли согласно описанию (37) в закрытых камерах, учитывая выделившееся за 7 сут количество СО2 методом титрования.
Температурный режим почвы регистрировали автоматическим прибором мониторинга климатических параметров САМ-СМ (Институт мониторинга климатических и экологических систем СО РАН, Россия).
ДНК для метагеномного анализа выделяли из почвенных образцов с помощью набора NucleoSpin Soil («Macherey-Nagel GmbH & Co. KG», Германия) в соответствии с инструкцией производителя.
Подготовка библиотек для высокопроизводительного секвенирования включала амплификацию целевого фрагмента вариабельного участка V4 гена 16S rRNA с использованием универсальных праймеров (515F — 5´-GTGCCAGCMGCCGCGGTAA-3´/806R — 5´-GGACTACVSGGGTAT-CTAAT-3´) (31) совместно с линкерами и уникальными баркодами. ПЦР проводилась на приборе T100 Thermal Cycler («Bio-Rad Laboratories», США) в 15 мкл реакционной смеси, содержащей 0,5 ед. ДНК-полимеразы Q5® High-Fidelity DNA Polymerase («New England BioLabs», США), 1X Q5 Reaction Buffer, 5 пмоль каждого из праймеров, 3,5 мM dNTP («Evrogen», Россия) и 1-10 нг ДНК-матрицы. Программа ПЦР включала стадию денатурации при 94 °С — 1 мин, амплификацию продукта в течении 35 циклов (94 °С — 30 с, 50 °С — 30 с, 72 °С — 30 с) и финальную элонгацию при 72 °С — 3 мин. Дальнейшую пробоподготовку и секвенирование проводили в соответствии с протоколом Illumina (16S Metagenomic Sequencing Library Preparation) на приборе Illumina MiSeq («Illumina, Inc.», США) с использованием набора реагентов MiSeq Reagent Kit v3 (600 cycle) с двусторонним чтением (2×300 n) («Illumina, Inc.», США).
Предобработка полученных данных включала удаление служебных последовательностей с использованием программы cutadapt (32), а также денойзинг, объединение парных прочтений и удаление химер при помощи пакета dada2 (33), реализованного в программной среде R. В нем же проводили таксономическую классификацию полученных последовательностей ASV (amplicon sequence variant) с использованием 132 релиза базы данных SILVA (https://www.arb-silva.de), содержащей данные для гена SSU rRNA (34). Дальнейшую обработку, включающую построение филогенетического дерева с помощью алгоритма SEPP (35), расчет α- и β-разнообразия выполняли в рамках пакета QIIME2 (36) и реализованных в нем плагинов. Для оценки α-разнообразия учитывали индексы разнообразия, отражающие фактическое прогнозируемое видовое богатство (Chao1), степень выравненности (Shannon, H) и доминантности (Simpson, D).
Для выявления дифференциально представленных таксонов использовали пакеты phyloseq (37) и DESeq2 (38). На основе результатов выполненного анализа была сформирована подвыборка филотипов, изменение обилия которых значимо при a ≤ 0,1 (с учетом поправки Benjamini-Hochberg).
Содержание общего органического углерода и азота, базальное дыхание почв, агрохимические показатели измеряли в 3-кратной аналитической повторности и представляли как средние (M) и стандартные ошибки средних (±SEM). Гранулометрический состава оценивали в одной повторности. Для оценки тесноты связи между параметрами обеспеченности почв элементами питания рассчитывали ранговый коэффициент корреляции r Спирмена. Достоверность различий между показателями для почв черневой тайги и олиготрофных местообитаний оценивали методом однофакторного дисперсионного анализа (one-way ANOVA). Таксономическую структуру проб почвенной микробиоты анализировали в 4 повторностях.
Результаты. Морфология и аналитические характеристики. Морфологическое описание и гранулометрический составе почв выбранных участков (рис. 1) представлены в таблице 1. Изученные разрезы охватывали основные компоненты педоразнообразия черневой тайги, представленного на покровных глинах такими контрастными вариантами, как темно-серые и дерново-подзолистые почвы (39).
Морфология изученных почв довольно сильно различается даже в том случае, если мы сравниваем почвы черневых экосистем друг с другом. Разрезы N1 и T1 представлены темно-серой суглинисто-глинистой почвой на лессовидных суглинках и темно-серой, остаточно-гумусовой, с темными кутанами, ветровально-турбированной почвой на лессовидной глине (рис. 2). В обеих почвах имеется хорошо развитый глубокий темногумусовый горизонт, имеющий более сложное строение в Т1, что связано с признаками остаточной гумусированности, а именно с уменьшением светлоты по шкале Манселла на 1 (см. табл. 1). Этот горизонт в обеих почвах характеризуется хорошо развитой трехмерной комковатой структурой, сменяющейся постепенно призматической и ореховато-призматической отдельностями (рис. 2, А, Б). Темногумусовый горизонт в первой случае переходит в субэлювиальную толщу с признаками элювиального горизонта EL в виде отдельных мелких морфонов. Во втором случае темногумусовый горизонт сменяется потечно-гумусовым горизонтом Ahh, который характеризуется повышенным влагосодержанием, наличием на гранях структурных отдельностей тонких темных глинисто-гумусовых кутан, подчеркнутых более темной окраской поверхностей раздела в сравнении с внутрипедной массой.
Темно-серые почвы в черневой тайге не относятся к широко распространенным. Для низкогорной части они ранее не были описаны, хотя проведенные нами исследования показали, что имеется высокая вероятность встретить их в нижних частях склонов, в ложбинах, то есть в местах с дополнительным увлажнением. Видимо, то, что такие почвы не упоминаются в работах других авторов по черневой тайге, связано с малой географической изученностью этого региона. Два обследованных нами профиля N1 и T1 различаются особенностями нижней части глубокого темногумусового горизонта. В профиле N1 появляются элювиальные морфоны, что связано с активным внутрипочвенным стоком в условиях низкогорного рельефа, когда в этой части происходит не аккумуляция, а концентрирование потоков и вынос тонкодисперсной фракции, которая и обогащена гумусом. На месте происходит относительное накопление скелетан, пылеватой фракции, имеющий светлые тона. Такие морфоны свидетельствуют об эволюции этой почвы в сторону темногумусовых подбелов, которые широко распространены в предгорной подтайге Западной Сибири (Приалтайская почвенная провинция), где занимают сниженные позиции в микро-, реже в мезорельефе. В профиле T1 в нижней части глубокого AU обособляется подгоризонт Ahh. Это второй гумусовый горизонт, несущий признаки иллювиальной трансформации, что диагностируется по темным гумусово-глинистым кутанам на поверхностях раздела (см. рис. 2, В). Глинистые кутаны оседают благодаря замедленному течению верховодок (что подтверждается серией полевых наблюдений в весеннее и позднелетнее время) в условиях ложбин, которые дренируют пологие склоны относительно равнинного рельефа северной части изученного ареала черневой тайги. Ниже в текстурном горизонте состав кутан сохраняется, но увеличивается их толщина и обилие (см. рис. 2, Г).
Почвы точек N2 и T2 представлены следующей стадией трофности, то есть не темно-серыми почвами, а стратоземом и дерново-подзолистой, обе они также характерны для черневой тайги. Стратозем темногумусовый среднесуглинистый на стратифицированных суглинках (делювии) расположен в эрозионной долине р. Каракан под травяным сосновым лесом с присутствием аконита. Дерново-подзолистая среднесуглинистая на лессовидном суглинке расположена в послерубочном осиново-березово-пихтовом высокотравном черневом лесе. Почвы точек N3 и T3 (референтные почвы олиготрофных сосняков) представлены относительно олиготрофными вариантами, поскольку в своем происхождении они связаны с легкими почвообразующими породами и сосновыми лесами: N3 — серогумусовая супесчаная на эоловых супесях в травяно-сосновом орляково-осоковом лесу, Т3 — дерново-элювозем супесчаный под олиготрофной экосистемой сосняка разнотравного с примесью лиственницы. При этом в пределах точки Т3 имеются виды растений, характерные для черневого леса: Cacalia hastata L., Aconitum septentrionale (Koelle) Korsh.,Milium effusum L. Однако их обилие и жизнеспособность существенно ниже, чем в изученных черневых лесах. Профили почв черневой тайги, как правило, более дифференцированы по содержанию ила и физической глины по сравнению с почвами олиготрофных вариантов (рис. 3), что свидетельствует об интенсивном развитии элювиальных процессов.
Таким образом, видно, что тип почвы и ее профильная организация во многом соответствуют типу леса. Так, черневая тайга не формируется на почвах, приуроченных к легким по гранулометрическому составу песчаным породам. Черневые леса приурочены либо к темно-серым, либо к дерново-подзолистым почвам с выраженным двучленным строением почвенного профиля, суглинисто-глинистым гранулометрическим составом.
Химический анализ показал постепенное убывание в профильном распределении углерода и азота (табл. 2), что в принципе типично для текстурно-дифференцированных и серогумусовых почв. Наибольшее содержание углерода органических соединений и азота характерно для верхних органоминеральных горизонтов почв черневой тайги, причем именно для темно-серых почв, что свидетельствует об их максимальной трофности. Почвы олиготрофных местообитаний имеют в среднем наименьшее содержание гумуса. Приблизительно те же закономерности касаются содержания общего азота. Соотношение углерода и азота (40) свидетельствует о средней или высокой обогащенности почв азотом. Изученные почвы близки к нейтральным или слабокислыми, что в принципе характерно для текстурно-дифференцированных почв. При этом почвы олиготрофных местообитаний более кислые, что связано с меньшей буферностью супесчаного мелкозема по сравнению с суглинистым (41). Интенсивность базального дыхания в целом была выше в олиготрофных почвах (точки N3 и T3), а по профилям — в подстилочных горизонтах.
Распределение доступных форм фосфора в почвенных профилях носило негомогенный характер. Так, имеется первый максимум в гумусовом горизонте, а также второй максимум в иллювиальной толще (табл. 3). Подобное распределение характерно для калия, хотя иногда второй максимум оказывается чуть ниже иллювиальной толщи. Что касается аммонийных и нитратных форм азота, то они концентрируются в основном в верхних горизонтах. При этом характерно преобладание нитратных форм азота в почвах.
Величины рангового коэффициента корреляции Спирмена, рассчитанные для почв черневой тайги и олиготрофных местообитаний (табл. 4), выявили в изученных почвах отрицательную связь между содержанием фосфора и калия, а также фосфора и аммонийных и нитратных форм азота. Эта корреляция была сильнее выражена в почвах черневой тайги, что указывает на более высокую степень их трофности по сравнению с почвами олиготрофных местообитаний. Коэффициенты корреляции для калия и остальных сравниваемых элементов имели положительные значения, а их величины были выше для почв черневой тайги. Накопление общего азота хорошо коррелировало с накоплением калия и нитратного азота. Для общего азота и аммонийного азота тесных связей мы не выявили.
Термический мониторинг, проведенный нами для темно-серой почвы (Т1), показал, что в зимний период (2019-2020 год) почва практически не промерзала. Отрицательные температуры (от -1 до 0 °С) начали проникать в почву в начале ноября, достигнув глубины 30 см к концу декабря, после чего нулевая завеса (температуры от +0,1 до -0,1 °С) медленно опустилась до глубины 40 см к середине февраля. В пределах этих глубин зимой наблюдалось большое постоянство температур. Лишь в первых сантиметрах от поверхности температура могла опускаться ниже -0,1 °С, не достигая -2 °С. В летний период почвы прогревались до +12 °С до глубины 50 см, а максимальная температура составляла +16 °С до глубины 10 см. Это, вероятно, связано, в том числе с высокой влажностью почвы.
Следует отметить, что для черневой тайги характерна необыкновенно высокая биологическая продуктивность всех слагающих биоценозы компонентов. Никакой другой автономный ландшафт в Сибири не может конкурировать с черневой тайгой ни по объему «живого вещества», ни по интенсивности его воздействия на геогенные и микроклиматические факторы. По имеющимся данным, масса ежегодного наземного опада растений составляет порядка 55-63 ц/га сухого вещества в год, а биомасса выражается в следующих цифрах: фитомасса — до 4000 ц/га, зоомасса (герпеторий и педобионты) — 4-8 ц/га, биомасса почвообитающих микроорганизмов — 80-90 ц/га (42). В черневых лесах из суммарного годового количества наземного опада 55-63 ц/га сухих веществ высокотравье поставляет 28-30 ц/га. Содержание азота и зольных элементов в опаде травянистого яруса значительно выше, нежели в материале древесного опада (зольность травянистого — 11,3, а древесного — 3,4 %, количество азота — соответственно 2,4 и 1,6 %) (26). Древесный ярус, имеющий огромную фитомассу, возвращает ежегодно в биологический круговорот непропорционально малое количество зольных элементов и азота — почти в 3-4 раза меньше их ежегодного поступления на поверхность почвы при отмирании травостоя. Запасы фитомассы в черневой тайге в 1,5-2 раза выше, чем в равнинной южной тайге Западной Сибири, запасы азота — в 2-2,5 раза, кальция — в 1,4-1,8 раза, ежегодное поступление Са с опадом выше в 4 раза (41). Известно об исследовании, согласно которому содержание фосфора в почвах черневой тайги — 879-1042 мг/кг (43). Отметим, что оно соответствуют верхнему диапазону, характерному для других экосистем биосферы Земли (включая дождевые леса тропического и умеренного пояса). Сообщалось, что по количеству актиномицетов и спорообразующих форм бактерий эти почвы приближаются к некоторым степным почвам, черноземам, при этом для них характерно небольшое абсолютное и относительное число грибов (33).
Скорость оборота подстилки в черневой тайге составляет 1-1,5 года (44). Подстилка состоит из нескольких фракций, различающихся скоростью разложения (наименьшая — у хвои пихты и кедра, листьев березы, опада веток и коры кустарников и деревьев). Лабильным, разлагающимся за год, является опад высокотравья. Отсутствие подстилки, удержание минеральных веществ от выщелачивания в условиях гумидного климата, мощное развитие высокотравья придают биологическому круговороту «тропические» черты, а черневую тайгу называют еще «сибирскими тропиками».
Еще одна особенность черневой тайги состоит в том, что валовое содержание кальция в почвообразующих породах (лессовидные глины) в слое 1 м составляет 3300 ц/га (44). Такое количество кальция проходит через биологический круговорот менее чем за 6-7 тыс. лет. Следовательно, при отсутствии надежного механизма рециклинга, за который отвечают высокотравные парцеллы, черневая тайга не смогла бы существовать. Именно это можно наблюдать в южной части лесной зоны Западной Сибири, где механизмы рециклинга были нарушены лесными пожарами.
Наиболее распространенный компонент почвенного покрова черневой тайги — текстурно-дифференцированные почвы (подзолистые, дерново-подзолистые и светло-серые). Тренд голоценовой эволюции этих почв состоял в обеднении корнеобитаемой зоны илом и периодическом перемешивании почвенной массы ветровалами. В настоящее время в пределах средней глубины корневых систем основных лесообразователей — пихты и осины сформировался относительно гомогенный горизонт EL или AEL, мощностью от 45 до 80 см, под которым залегает горизонт BT. Наши промеры глубин ветровальных западин пихты и осины в черневой тайге показали, что их средняя глубина в условиях нормального увлажнения составляет 51 см, при переувлажнении — 40 см. При этом достоверных различий в глубине западин осины и пихты мы не обнаружили. Подобное строение почв отвечает определению климаксного профиля лесной почвы и свидетельствует об отсутствии существенных экзогенных нарушений, прерывавших устойчивый поток поколений древесных видов.
Почвенный покров черневой тайги — феномен высотной дифференциации ландшафтов на горных макросклонах юга Сибири, открытых западным влажным воздушным течениям. В горах и предгорьях у черневой тайги имеется верхний предел распространения, выше которого в области среднегорного рельефа в почвенных профилях доминируют горные подзолистые почвы, элювоземы — псевдоподзолистые почвы без иллювиального горизонта (42) и буроземы (45). Географически общее для всех почв черневой тайги — наличие хорошо оструктуренной, с качественными водно-физическими свойствами, облегченной по гранулометрическому составу толщи, представленной в изученных нами почвах горизонтами AY, AEL, EL, AU, Ahh. Ниже этих горизонтов всегда залегает водоупорный горизонт, будь то покровные суглинки и глины, преобразованный в горизонт BT, или обломочная кора выветривания и залегающий глубже плотный горизонт R. Наличие подобного двучленного строения благоприятствует формированию верховодок и более длительному сохранению влаги, что в условиях суббореального пояса и высокого расчленения рельефа поддерживает возможность существования высокотравных сообществ с достаточной влагообеспеченностью в сухой сезон. Вне подобного двучленного строения почв функционирование высокотравных экосистем невозможно, высокотравье будет сменяться разнотравьем или осочковыми фитоценозами. Весной и в сезон дождей наличие хорошо оструктуренной полуметровой толщи способствует инфильтрации влаги, отчего в верхнем корнеобитаемом горизонте всегда сохраняется благоприятный воздушный режим, способствующий благоприятному функционированию корневых систем. Это доказывается и тем, что в олиготрофной точке N3, в которой отсутствовал водоупорный горизонт, в сообществе не было высокотравных травянистых видов, а в точке T3 эти виды были, так как наличие горизонта D в указанной точке способствует благоприятному водному режиму, хотя недостаточная трофность местообитания не позволяет высокотравным видам удерживать свои фитоценотические позиции.
В изученных почвах также сильно различались кутанные комплексы. Темно-серые почвы, занимающие вогнутые склоны, имеют лучше сформированный кутанный комплекс, чем дерново-подзолистые почвы.
В черневой тайге два основных древесных эдификатора — пихта сибирская Abiessibirica Ledeb. и осина Populus tremula L. При движении к северу, подъеме в горы, в нижних частях склонов осина может уступать первенство березе (Betula pendula L.). Черневые экосистемы тесно связаны с дерново-подзолистыми сверхглубокоосветленными почвами, которые претерпевают изменения на протяжении ареала. Как уже отмечалось, наиболее значимы не сами почвы, а их элювиальные горизонты при наличии нижележащего водоупорного горизонта, о чем свидетельствует широкое распространение элювоземов в горах.
Почвы с мощным элювиальным профилем доминируют не только в почвенных профилях черневой тайги, они также развиты на хорошо дренированных равнинах Западной Сибири (46-49). Для сверхглубокоподзолистых почв равнины, как и для гор, характерно распространение на них пихтовых лесов либо лесов с доминированием пихты. Как отмечается (50), коэффициент связи пихтовых фитоценозов с дерново-подзолистыми почвами равен 1,0. При экспедиционном обследовании мы подтвердили, что пихта обладает глубокопроникающей корневой системой (большое число якорных корней сосредоточено в толще 1 м), причем она реагирует на характер увлажнения и на переувлажненных территориях распространение корней сменяется поверхностным. С пихтой распространен и кедр (Cedrus Trew, 1757, nom. cons.), также обладающий глубокой корневой системой. За период голоцена, когда пихтовая формация занимала территории с глубокоподзолистыми почвами, происходило неоднократное оборачивание почвенного пласта при ветровалах, о чем свидетельствует комплекс признаков ветровальных нарушений. При сукцессиях ветровально-турбированных почв усиливаются процессы, вызывающие элювиальное оглеение и лессиваж (51, 52), что приводит к текстурной дифференциации. Можно предположить, что там, где условия для глубокой ветровальности (с вовлечением толщи до 1 м) складывались благоприятнее, происходило формирование более глубоко элювиированных почв. Усилению элювиирования, несомненно, благоприятствовала энергичная внутрипочвенная эрозия, усиливающая морфогенетический эффект от ветровалов.
Высокое содержание гумуса вообще характерно для темно-серых почв (53). При этом изученные нами темно-серые почвы характеризовались развитым в глубину гумусовым профилем, что свидетельствует об их высоком плодородии. Содержание гумуса было несколько ниже в дерново-подзолистых почвах и еще ниже в серогумусовых и дерново-подзолах олиготрофных сосновых лесов, что соответствует данным литературы (54). В изученных нами почвах не наблюдалась элювиально-иллювиальной дифференциация гумусовых профилей, что в большей степени характерно для текстурно-дифференцированных почв, чем для альфегумусовых (55). Кислотность почв, по нашим данным, вполне соответствовала их природе и была повышенной в случае серогумусовых и дерново-подзолистых почв, что связано с типом растительного опада (сосновый) и легким гранулометрическим составом почвообразующих пород.
Разобщенность иллювиальных профилей различных элементов питания обусловлена неодинаковой подвижностью их форм в почвенной среде (55). Само же наличие второго максимума связано с ролью элювиального типа почвообразования в формировании биологически активных запасов элементов питания в различных частях почвенного профиля. Отмеченное нами преобладание нитратных форм азота, легкодоступных для растений (особенно в верхних горизонтах), свидетельствует о важной роли поступления нутриентов из опада.
Таким образом, темно-серые почвы черневой тайги представляют собой своеобразный вариант текстурно-дифференцированных почв, отличающихся от европейских аналогов из нечерневых лесов развитостью гумусового профиля и повышенным содержанием элементов питания, что способствует формированию особого трофического статуса с выраженным феноменом гигантизма. Одновременно с этим наблюдается снижение степени морфохроматической дифференциации профилей почв в связи с пропитанностью гумусовыми веществами.
Характеристика таксономической структуры почвенной микробио т ы. Анализ таксономической структуры почвенной прокариотической микробиоты фоновой зональной почвы (ZT) и почвы черневой тайги (ChT), выполненный в четырех повторностях, показал, что на уровне прокариотических филумов обе почвы имеют близкую структуру и содержат представителей 15 филумов, обычных для почвенных микробных сообществ. Среди них доминировали Proteobacteria, Verrucomicrobia, Actinobacteria, Acidobacteria, Planctomycetes и Firmicutes, а также весьма высокой была доля неклассифицируемых микроорганизмов (рис. 4, А).
На уровне родов (см. рис. 4, Б) может быть выявлена гораздо более тонкая структура, из которой очевидно, что различия между почвами скорее всего обусловлены различиями между таксонами более низкого ранга, чем филумы. Выявленные доминанты почвенной микробиоты характерны для большинства природных почв умеренного пояса (56), к которому относится бореальный тип почв и лесов. Присутствие ацидобактерий типично для почв со слабокислой реакцией среды (38), что характерно для изученных таежных почв. Представители филы Firmicutes могут участвовать в разложении органических алькильных фрагментов лесной подстилки, что также имеет место в изученных почвах, где происходит остаточное накопление хорошо гумифицированного органического вещества. Также в деградации растительных полимеров способны принимать участие Actinobacteria, что подтверждает высокую интенсивность биологического круговорота в лесных экосистемах черневой тайги и быструю минерализацию низкомолекулярных компонентов органического вещества почв. Представители Verrucomicrobia индицируют наличие интенсивного зоогенного (возможно, копрогенного) фактора в почвообразовании, что также подтвердили результаты выполненного нами морфологического исследования почв.
Анализ индексов разнообразия (см. рис. 4, В) показал, что по числу выявляемых таксонов между почвами нет существенной разницы, однако по индексам Симпсона и Шеннона (57) можно видеть, что есть, хотя и незначительные, но тем не менее достоверные различия в параметрах, обусловленных как ровностью, так и богатством разнообразий. Суммарный эффект можно видеть при анализе β-разнообразия (см. рис. 4, Г), где видна четкая дифференциация между микробиотой фоновых зональных почв и почв черневой тайги.
Понятно, что наиболее интересны возможные различия в таксономической структуре микрбиомов фоновой зональной почвы и почвы черневой тайги с целью выявления прокариотических таксонов, предположительно связанных с высоким плодородием. В результате проведенного анализа были выявлены таксоны, численность которых достоверно увеличивается при переходе от фоновой зональной почвы к почве черневой тайги. Суммарная доля таких таксонов составила 5,6 % в фоновой почве и 32,2 % в почве черневой тайги. В таблице 5 приведен список выявленных таксонов ранга рода, сгруппированных на уровне прокариотических порядков, демонстрирующих достоверное увеличение численности при переходе от фоновой почвы к почве черневой тайги.
Более половины дифференциально представленных таксонов относятся к неклассифицируемым прокариотам, как, впрочем, и во всей ис-ходной микробиоте. Однако среди таксонов, которые удалось идентифицировать, можно выделить несколько весьма интересных. В таблице 5 таксоны отсортированы по численности в почве черневой тайги. В этом списке доминируют роды порядков Rhizobiales, Chthoniobacterales, Bacillales, Myxococcales, каждый из которых потенциально имеет некоторую связь с почвенным плодородием, хотя утверждать это определенно затруднительно. Тем не менее порядок Rhizobiales, несомненно, может быть связан с почвенными, преимущественно симбиотическими, азотфиксаторами. Bacillales и Myxococcales, а также представленные в списке Chitinophagales могут иметь отношение к разложению органического вещества.
Довольно интересен отмеченный в этом списке порядок Chthoniobacterales: лишь недавно первый представитель этого порядка был выделен в культуру (58) и его геном просеквенирован (59). Этот таксон интересен не только тем, что принадлежит к филуму Verrucomicrobia, экологическая значимость которого (особенно в почвенных сообществах) становится очевидной в последнее время, но и своим «говорящим» названием, имеющим некоторую связь с темой нашего исследования, так как происходит от греческого слова χθών (земля, почва) и используется в древнем мифологическом и современном философском дискурсе для обозначения «хтонических» существ и сущностей, олицетворяющих собой первобытную природную мощь земли. Конечно, не стоит слишком однозначно воспринимать это обстоятельство, но и не обратить на него внимание нельзя, так как этот таксон — один из доминантов, предположительно имеющих отношение к плодородию почв черневой тайги. В любом случае очевидно, что таксономический и функциональный состав специфического компонента микробиоты почв черневой тайги может стать источником новых знаний о механизмах формирования и поддержания почвенного плодородия.
Обращает внимание фактическое отсутствие архейных филумов в фоновой почве, в то время как филум Thaumarchaeota (0,1 %) представлен в почве черневой тайги.
Итак, в результате выполненного исследования мы уточнили морфологическую организацию, таксономическое положение, термический режим и гранулометрический состав почв черневой тайги Западной Сибири. Установлено, что почвы черневой тайги Западной Сибири в основном относятся к отделу текстурно-дифференцированных почв, типам дерново-подзолистых, серых и темно-серых почв с тяжелосуглинистым и легкоглинистым гранулометрическим составом почвообразующих пород. Эти почвы формируются в уникальных комбинациях геогенных и биоклиматических условий, не промерзают зимой, обеспечены влагой, что способствует быстрой минерализации подстилочного материала и закреплению минеральных элементов питания в верхней гумусированной толще почвенного профиля. Аккумуляция биофильных элементов — важнейшее свойство почв черневой тайги, с которым связан феномен гигантизма и крайне высокой продуктивности растений. Расположенные в смежных биотопах на легких почвообразующих породах почвы олиготрофных лесов бедны по показателям агрохимического плодородия, не имеют выраженного гумусированного профиля и относятся либо к серогумусовым, либо к альфегумусовым и дерново-элювиальным вариантам. На уровне прокариотических филумов обе почвы имеют близкую структуру и содержат представителей 15 филумов, обычных для почвенных микробных сообществ. В целом таксономический состав микробных филумов соответствует таковому в умеренно-влажных почвах умеренного пояса. Различия между почвами скорее всего обусловлены различиями между таксонами более низкого ранга, чем филумы. Разнообразие микроорганизмов в изученных почвах различается в зависимости от режима трофности экосистемы. Количество филотипов в пробах почв черневой тайги повышено по сравнению с олиготрофными местообитаниями. Почвы черневой тайги характеризуются бóльшим разнообразием микробного сообщества по индексу Шеннона, а также присутствием филумов Nitrospirae и Thaumarchaeota,которых нет в почвах олиготрофных местообитаний. Фила Actinobacteria, являющаяся одной из прокариотических доминант, обеспечивает высокую интенсивность биологического круговорота в лесных экосистемах черневой тайги и быструю минерализацию низкомолекулярных компонентов органического вещества почв. Таким образом, почвы черневой тайги обладают специфической микробиотой и соответствующими микробными драйверами почвенных процессов, ответственными за продуктивность этих почв. Они представляют собой уникальный компонент бореальных экосистем Западной Сибири, позволяющий получить новые знания о механизмах повышенной продуктивности почв при уникальном сочетании биоклиматических и геогенных факторов.
1. Морфологическая характеристика, окраска почвенных горизонтов, содержание скелета и мелкозема в исследованных почвах черновой тайги (Западная Сибирь, 2019 год) |
||||
Глубина, см |
Горизонт |
Код цвета по Манселлу |
Содержание скелета % |
Содержание мелкозема, % |
N1, темно-серая почва (Алтайский край) |
||||
0-10 |
O |
10 YR 3/1 |
Не определяли |
Не определяли |
10-20 |
AU |
10 YR 4/1 |
11 |
89 |
20-30 |
AU |
10 YR 4/1 |
12 |
88 |
30-40 |
BEL |
10 YR 6/2 |
18 |
82 |
40-50 |
BEL |
10 YR 6/2 |
21 |
79 |
60-80 |
BI |
5 YR 6/3 |
19 |
81 |
80-100 |
BC |
5 YR 6/3 |
21 |
79 |
N2, с тратозем темногумусовый (Новосибирская обл.) |
||||
0-2 |
O |
10 YR 3/1 |
Не определяли |
Не определяли |
2-10 |
AU |
10 YR 4/1 |
22 |
78 |
10-20 |
RU |
5 YR 2.5/1 |
11 |
89 |
30-40 |
RU |
5 YR 2.5/1 |
0 |
100 |
60-70 |
RU |
5 YR 2.5/1 |
0 |
100 |
80-90 |
C |
7.5 YR 8/1 |
0 |
100 |
N3, серогумусовая почва на эоловых супесях (Новосибирская обл.) |
||||
0-3 |
O |
10 YR 4/1 |
Не определяли |
Не определяли |
3-15 |
AY |
7.5 YR 8/1 |
6 |
94 |
20-30 |
AC |
7.5 YR 8/1 |
4 |
96 |
40-50 |
C |
7.5 YR 8/1 |
2 |
98 |
T1, темно-серая почва (Томская обл.) |
||||
0-1 |
O |
10 YR 4/1 |
Не определяли |
Не определяли |
1-15 |
AU |
10 YR 4/1 |
19 |
81 |
15-30 |
AU |
10 YR 4/1 |
18 |
82 |
35-55 |
Ahh |
10 YR 3/3 |
19 |
81 |
70-110 |
BTtur |
5 YR 4/6 |
21 |
79 |
Т2, дерново-подзолистая почва (Кемеровская обл.) |
||||
0-3 |
AY |
7.5 YR 6/1 |
93 |
7 |
3-17 |
EL |
5 YR 7/1 |
25 |
75 |
20-30 |
BEL |
5 YR 7/1 |
18 |
82 |
30-40 |
BT |
5 YR 4/6 |
21 |
79 |
40-50 |
BT |
5 YR 4/5 |
22 |
78 |
60-70 |
BT |
5 YR 4/5 |
23 |
77 |
80-90 |
BCi |
5 YR 6/3 |
5 |
95 |
105-120 |
BCi |
5 YR 6/3 |
10 |
90 |
T3, дерново-элювозем (Томская обл.) |
||||
0-3 |
O |
10 YR 4/1 |
Не определяли |
Не определяли |
10-20 |
AY |
7.5 YR 8/1 |
5 |
95 |
40-50 |
EL |
5 YR 7/1 |
2 |
98 |
50-60 |
BT |
5 YR 4/6 |
1 |
99 |
60-80 |
BT |
5 YR 4/6 |
1 |
99 |
Примечание. N1, N2, T1, T2 — почвы черневой тайги, N3, T3 — референтные почвы олиготрофных сосняков. |
||||
2. Содержание углерода, азота, кислотность и базальное дыхание изученных почв (n = 3, M±SEM, Западная Сибирь, 2019 год) |
||||||
Код, |
C, % |
N, % |
C/N |
рН вытяжки |
Базальное дыхание, мг СО2/(100 г почвы/сут) |
|
водной |
солевой |
|||||
N1 0-10 |
4,76±0,23 |
0,37±0,05 |
12,81 |
6,56 |
5,82 |
0,022±0,003 |
N1 10-20 |
3,22±0,17 |
0,27±0,04 |
11,57 |
6,13 |
5,20 |
0,019±0,002 |
N1 20-30 |
2,40±0,17 |
0,18±0,03 |
13,00 |
6,02 |
4,69 |
0,013±0,002 |
N1 30-40 |
2,25±0,10 |
0,17±0,03 |
12,88 |
5,85 |
4,64 |
0,021±0,002 |
N1 40-50 |
1,41±0,07 |
0,11±0,02 |
12,71 |
5,44 |
4,89 |
0,021±0,002 |
N1 60-80 |
0,45±0,05 |
0,06±0,02 |
8,14 |
6,30 |
4,53 |
0,020±0,004 |
N1 80-100 |
0,24±0,04 |
0,05±0,01 |
5,42 |
6,39 |
4,57 |
0,028±0,003 |
N2 0-2 |
2,14±0,21 |
0,16±0,01 |
13,51 |
6,95 |
5,96 |
0,062±0,001 |
N2 2-10 |
2,02±0,19 |
0,15±0,01 |
13,59 |
7,35 |
Не определяли |
0,047±0,009 |
N2 10-20 |
0,23±0,04 |
0,03±0,01 |
6,45 |
7,22 |
Не определяли |
0,037±0,008 |
N2 30-40 |
0,16±0,04 |
0,03±0,01 |
5,68 |
7,09 |
Не определяли |
0,040±0,002 |
N2 60-70 |
0,15±0,02 |
0,03±0,01 |
5,18 |
6,81 |
5,70 |
0,041±0,004 |
N2 80-90 |
0,46±0,09 |
0,05±0,01 |
8,44 |
6,65 |
5,65 |
0,041±0,004 |
N3 0-3 |
2,70±0,21 |
0,17±0,03 |
15,46 |
6,70 |
6,20 |
0,051±0,004 |
N3 3-15 |
0,21±0,04 |
0,03±0,01 |
7,49 |
5,93 |
5,52 |
0,029±0,007 |
N3 20-30 |
0,05±0,02 |
0,02±0,01 |
2,85 |
6,06 |
4,91 |
0,045±0,008 |
N3 40-50 |
0,03±0,05 |
0,01±0,01 |
6,00 |
6,39 |
5,38 |
0,052±0,007 |
T1 0-1 |
9,85±0,28 |
0,62±0,04 |
15,66 |
6,59 |
6,32 |
0,206±0,006 |
T1 1-15 |
3,46±0,25 |
0,28±0,05 |
12,31 |
5,96 |
5,14 |
0,048±0,007 |
T1 15-30 |
2,37±0,20 |
0,31±0,02 |
7,67 |
5,93 |
4,71 |
0,041±0,004 |
T1 35-55 |
1,84±0,12 |
0,14±0,02 |
12,73 |
5,28 |
4,69 |
0,043±0,001 |
T1 70-110 |
0,76±0,09 |
0,08±0,02 |
9,00 |
5,97 |
4,86 |
0,048±0,001 |
T2 0-3 |
3,97±0,42 |
0,37±0,02 |
10,75 |
6,72 |
6,16 |
0,071±0,007 |
T2 3-17 |
1,38±0,12 |
0,14±0,03 |
9,44 |
5,33 |
4,82 |
0,053±0,002 |
T2 20-30 |
0,56±0,07 |
0,06±0,02 |
8,08 |
5,70 |
4,38 |
0,028±0,003 |
T2 30-40 |
0,38±0,07 |
0,04±0,01 |
7,89 |
5,17 |
4,26 |
0,053±0,008 |
T2 40-50 |
0,47±0,09 |
0,07±0,01 |
6,98 |
6,03 |
4,25 |
0,059±0,008 |
T2 60-70 |
0,29±0,04 |
0,05±0,01 |
6,30 |
5,61 |
4,17 |
0,055±0,007 |
T2 80-90 |
0,20±0,04 |
0,04±0,01 |
4,98 |
5,86 |
4,30 |
0,052±0,008 |
T3 0-3 |
2,41±0,31 |
0,20±0,03 |
12,24 |
6,96 |
6,17 |
0,078±0,001 |
T3 10-20 |
0,36±0,08 |
0,04±0,02 |
8,62 |
5,16 |
4,40 |
0,053±0,003 |
T3 40-50 |
0,07±0,02 |
0,02±0,01 |
3,57 |
5,18 |
4,54 |
0,042±0,003 |
T3 50-60 |
0,09±0,02 |
0,03±0,01 |
3,26 |
5,69 |
4,66 |
0,057±0,004 |
T3 70-110 |
0,07±0,01 |
0,03±0,01 |
2,25 |
5,73 |
4,69 |
0,063±0,007 |
One-way ANOVA: |
|
|
|
|
|
|
|
р < 0,04 |
р < 0,03 |
|
|
|
р < 0,04 |
Примечание. N1, N2, T1, T2 — почвы черневой тайги, N3, T3 — референтные почвы олиготрофных сосняков. |
||||||
3. Агрохимические показатели изученных почв (n = 3, M±SEM, Западная Сибирь, 2019 год) |
||||
Код, глубина, см |
P, мг/кг |
K, мг/кг |
N-NH4, мг/кг |
N-NO3, мг/кг |
N1 10-20 |
343±21 |
319±25 |
11,14±2,15 |
15,57±1,53 |
N1 20-30 |
357±24 |
266±17 |
4,33±0,23 |
10,07±0,58 |
N1 30-40 |
702±34 |
217±14 |
2,76±0,22 |
8,59±0,41 |
N1 40-50 |
460±22 |
71±6 |
0,73±0,06 |
5,37±0,32 |
N1 60-80 |
605±23 |
133±8 |
0,41±0,07 |
5,37±0,42 |
N1 80-100 |
682±45 |
120±8 |
0,41±0,04 |
4,97±0,41 |
N2 0-2 |
255±20 |
198±11 |
10,23±0,97 |
14,53±1,01 |
N2 2-10 |
210±13 |
174±12 |
5,42±0,23 |
11,24±0,85 |
N2 10-20 |
178±11 |
125±8 |
0,41±0,05 |
9,54±0,41 |
N2 30-40 |
223±11 |
114±8 |
0,32±0,04 |
4,51±0,31 |
N2 60-70 |
200±15 |
95±8 |
0,25±0,03 |
7,54±0,52 |
N2 80-90 |
210±13 |
152±11 |
0,95±0,08 |
8,56±0,45 |
N3 3-15 |
113±8 |
195±16 |
0,57±0,05 |
7,11±0,40 |
N3 20-30 |
131±9 |
71±5 |
0,49±0,04 |
4,56±0,25 |
N3 40-50 |
243±12 |
58±4 |
0,73±0,05 |
4,03±0,14 |
Т1 1-15 |
234±15 |
470±45 |
7,32±0,54 |
8,59±0,61 |
Т1 15-30 |
166±10 |
355±32 |
2,20±0,04 |
7,52±0,45 |
Т1 35-55 |
231±16 |
262±22 |
1,06±0,08 |
8,19±0,35 |
Т1 70-110 |
373±16 |
186±14 |
0,49±0,05 |
6,71±0,50 |
Т2 3-17 |
79±7 |
200±29 |
12,69±0,89 |
18,93±0,33 |
Т2 20-30 |
76±7 |
106±15 |
1,79±0,09 |
9,66±0,12 |
Т2 30-40 |
107±8 |
62±6 |
0,65±0,05 |
6,98±0,15 |
Т2 40-50 |
104±7 |
98±8 |
Н.п.о. |
7,25±0,40 |
Т2 60-70 |
82±5 |
142±12 |
0,24±0,03 |
12,89±0,60 |
Т2 80-90 |
87±5 |
151±14 |
Н.п.о. |
9,40±0,10 |
Т2 105-120 |
184±6 |
160±12 |
Н.п.о. |
8,46±0,20 |
Т3 10-20 |
44±5 |
106±11 |
0,09±0,01 |
12,21±0,98 |
Т3 40-50 |
97±9 |
18±2 |
0,16±0,02 |
8,05±0,14 |
Т3 50-60 |
319±22 |
22±2 |
0,08±0,02 |
6,31±0,50 |
Т3 70-110 |
234±24 |
71±3 |
0,16±0,03 |
8,32±0,32 |
One-way ANOVA |
р < 0,05 |
р < 0,04 |
р < 0,04 |
р < 0,03 |
Примечание. N1, N2, T1, T2 — почвы черневой тайги, N3, T3 — референтные почвы олиготрофных сосняков. Н.п.о. — ниже пределов обнаружения. |
||||
4. Ранговые коэффициенты корреляции Спирмена (r, р = 0,05) для агрохимических показателей изученных почв (Западная Сибирь, 2019 год) |
||||||
Показатель |
P |
K |
N-NH4 |
N-NO3 |
C |
N |
Почвы черневой тайги (N1, N2, T1, T2) |
||||||
P |
1 |
-0,75 |
-0,37 |
-0,27 |
-0,48 |
-0,55 |
K |
-0,75 |
1 |
0,78 |
0,69 |
0,17 |
0,20 |
N-NH4 |
-0,37 |
0,78 |
1 |
0,93 |
-0,06 |
-0,003 |
N-NO3 |
-0,27 |
0,69 |
0,93 |
1 |
0,08 |
0,11 |
C |
-0,48 |
0,17 |
-0,06 |
0,08 |
1 |
0,98 |
N |
-0,55 |
0,20 |
-0,03 |
0,11 |
0,98 |
1 |
Почвы олиготрофных местообитаний (N3, T3) |
||||||
P |
1 |
-0,36 |
0,24 |
-0,67 |
-0,17 |
-0,10 |
K |
-0,36 |
1 |
0,19 |
0,45 |
0,40 |
0,46 |
N-NH4 |
-0,24 |
-0,19 |
1 |
-0,71 |
-0,03 |
-0,04 |
N-NO3 |
-0,67 |
0,45 |
-0,71 |
1 |
0,35 |
0,37 |
C |
-0,17 |
0,40 |
-0,03 |
0,35 |
1 |
0,98 |
N |
-0,10 |
0,46 |
-0,04 |
0,37 |
0,98 |
1 |
5. Таксоны, увеличивающие численность при переходе от фоновой почвы (ZT) к почве черневой тайги (ChT), сгруппированные на уровне прокариотических порядков (Западная Сибирь, 2019 год) |
|||||
Филум |
Порядок |
Численность таксона, % |
Кратность увеличения |
Число родов |
|
ZT средняя |
ChT средняя |
||||
unclassified |
|
3,085 |
18,616 |
6,0 |
138 |
Proteobacteria |
Rhizobiales |
0,648 |
3,193 |
4,9 |
11 |
Verrucomicrobia |
Chthoniobacterales |
0,581 |
2,744 |
4,7 |
7 |
Firmicutes |
Bacillales |
0,120 |
1,456 |
12,1 |
7 |
Proteobacteria |
Myxococcales |
0,289 |
1,266 |
4,4 |
10 |
Actinobacteria |
Propionibacteriales |
0,199 |
0,772 |
3,9 |
5 |
Bacteroidetes |
Chitinophagales |
0,044 |
0,686 |
15,5 |
5 |
Actinobacteria |
Gaiellales |
0,106 |
0,451 |
4,3 |
2 |
Acidobacteria |
Pyrinomonadales |
0,131 |
0,382 |
2,9 |
1 |
Actinobacteria |
Micromonosporales |
0,106 |
0,304 |
2,9 |
2 |
Planctomycetes |
Pirellulales |
0,034 |
0,255 |
7,6 |
2 |
Actinobacteria |
Frankiales |
0,012 |
0,200 |
16,9 |
1 |
Verrucomicrobia |
unclassified_Verrucomicrobiae |
0,053 |
0,197 |
3,7 |
2 |
Actinobacteria |
Microtrichales |
0,014 |
0,171 |
12,5 |
2 |
Actinobacteria |
Corynebacteriales |
0,012 |
0,160 |
13,5 |
1 |
Actinobacteria |
Solirubrobacterales |
0,044 |
0,153 |
3,5 |
2 |
Proteobacteria |
Steroidobacterales |
0,005 |
0,147 |
29,3 |
1 |
Actinobacteria |
unclassified_Actinobacteria |
0,005 |
0,146 |
26,8 |
1 |
Verrucomicrobia |
Pedosphaerales |
0,026 |
0,139 |
5,4 |
1 |
Proteobacteria |
Xanthomonadales |
0,018 |
0,106 |
5,8 |
2 |
Actinobacteria |
Micrococcales |
0,018 |
0,094 |
5,3 |
1 |
Gemmatimonadetes |
Gemmatimonadales |
0,024 |
0,090 |
3,7 |
1 |
Bacteroidetes |
Flavobacteriales |
0,004 |
0,073 |
19,7 |
1 |
Actinobacteria |
unclassified_Actinobacteria |
0,010 |
0,066 |
6,9 |
1 |
Planctomycetes |
Isosphaerales |
0,019 |
0,063 |
3,3 |
1 |
Planctomycetes |
Tepidisphaerales |
0,008 |
0,063 |
7,6 |
1 |
Actinobacteria |
Streptosporangiales |
0,004 |
0,059 |
14,9 |
1 |
Actinobacteria |
Streptomycetales |
0,014 |
0,046 |
3,3 |
1 |

Рис. 1. Профили исследованных почв черневой тайги (N1, N2, T1, T2) и референтные почвы олиготрофных сосняков (N3, T3): а — темно-серая (N1); б — стратозем темногумусовый (N2); в — серогумусовая почва на эоловых супесях (N3); г — темно-серая почва на покровных глинах (T1); д — дерново-подзолистая типичная почва на покровных глинах (T2); е — дерново-элювозем (T3) (Западная Сибирь, июль 2019 года, подробнее см. раздел «Методика»).

Рис. 2. Мезоморфология темно-серой почвы черневой тайги: А — лесной вариант горизонта AU, верхняя часть; Б — ореховатая структура нижней части горизонта AU (лесной вариант); В — кутаны в подгоризонте Ahh нижней части глубокого AU (лесной вариант); Г — кутаны в горизонте BT (Западная Сибирь, 2019 год).

Рис. 3. Профильные диаграммы гранулометрического состава исследованных почв: N1, N2, T1, T2 — почвы черневой тайги, N3, T3 — референтные почвы олиготрофных сосняков (Западная Сибирь, 2019 год). Описание почвенных проб см. в разделе «Методика» и таблице 1.
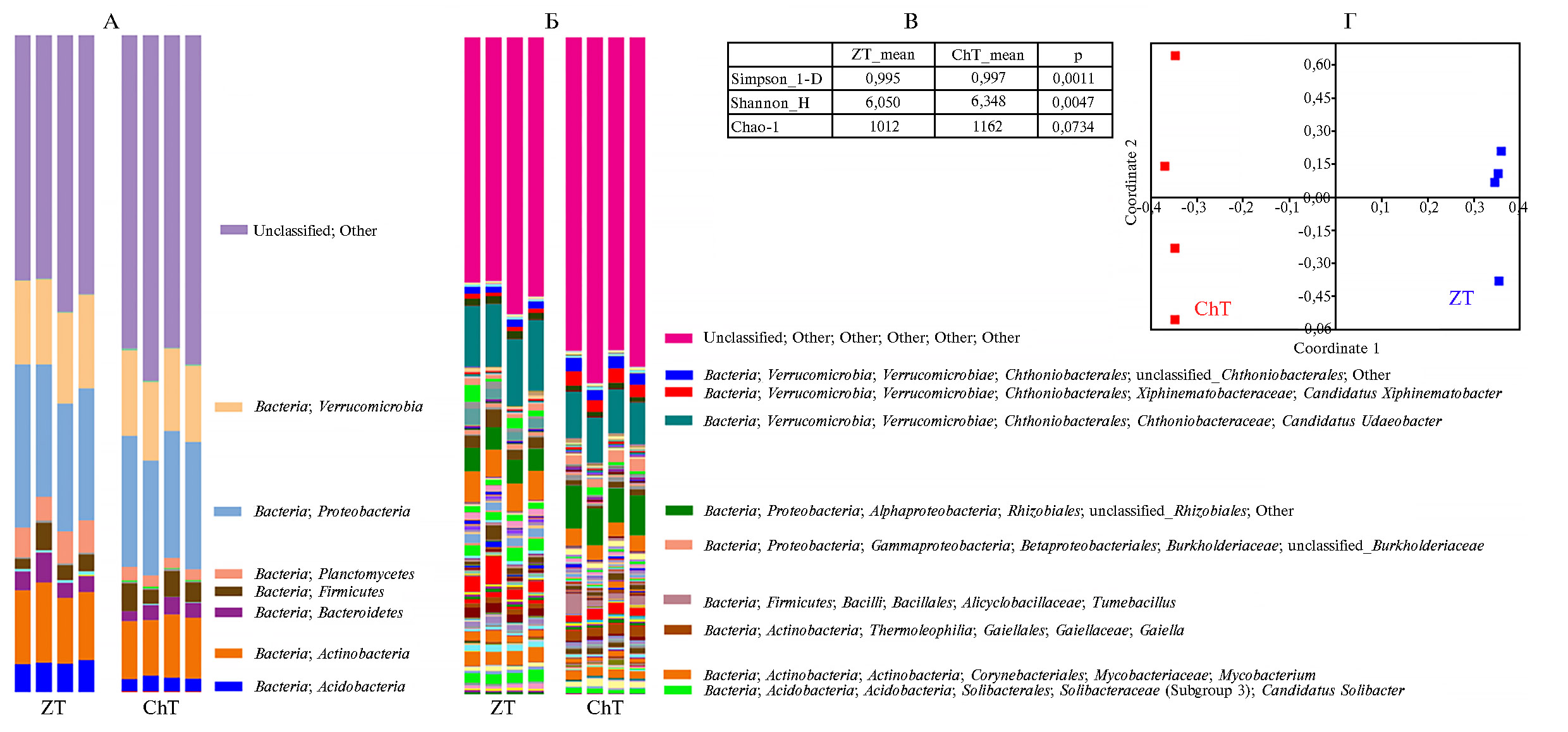
Рис. 4. Таксономическая структура микробиоты фоновой таежной почвы (ZT) и почвы черневой тайги (ChT) на уровне филумов (А) и родов (Б); представлены индексы разнообразия (В) и результаты анализа b-разнообразия (метод главных координат, расстояния по Брэю-Кертису) (Г) (Западная Сибирь, 2019 год).
ЛИТЕРАТУРА
- Vitousek P.M., Mooney H.A., Lubchenco J., Melillo J.M. Human domination of Earth’s ecosystems. Science, 1997, 277(5325): 494-499 (doi: 10.1126/science.277.5325.494).
- Clark C. M., Tilman D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature, 2008, 451(7179): 712 (doi: 10.1038/nature06503).
- Diaz R.J., Rosenberg R. Spreading dead zones and consequences for marine ecosystems. Science, 2008, 321(5891): 926-929 (doi: 10.1126/science.1156401).
- Marques A., Martins I.S., Kastner T., Plutzar C., Theurl M.C., Eisenmenger N., Huijbregts M.A.J., Wood R., Stadler K., Bruckner M., Canelas J., Hilbers J.P., Tukker A., Erb K., Pereira H.M. Increasing impacts of land use on biodiversity and carbon sequestration driven by population and economic growth. Nature Ecology and Evolution, 2019, 3(4): 628-637 (doi: 10.1038/s41559-019-0824-3).
- Tilman D. Biodiversity and environmental sustainability amid human domination of global ecosystems. Daedalus, 2012, 141(3): 108-120 (doi: 10.1162/DAED_a_00166).
- Song X.-P., Hansen M.C., Stehman S.V., Potapov P.V., Tyukavina A., Vermote E.F., Townshend J.R. Global land change from 1982 to 2016. Nature, 2018, 560(7720): 639-643 (doi: 10.1038/s41586-018-0411-9).
- Foley J.A., DeFries R., Asner G.P., Barford C., Bonan G., Carpenter S.R., Chapin F.S., Coe M.T., Daily G.C., Gibbs H.K., Helkowski J.H., Holloway T., Howard E.A., Kucharik C.J., Monfreda C., Patz J.A., Prentice I.C., Ramankutty N., Snyder P.K. Global consequences of land use. Science, 2005, 309(5734): 570-574 (doi: 10.1126/science.1111772).
- Davidson E.A. The contribution of manure and fertilizer nitrogen to atmospheric nitrous oxide since 1860. Nature Geoscience, 2009, 2(9): 659-662 (doi: 10.1038/ngeo608).
- Guo J.H., Liu X.J., Zhang Y., Shen J.L., Han W.X., Zhang W.F., Christie P., Goulding K.W.T., Vitousek P.M., Zhang F.S. Significant acidification in major chinese croplands. Science, 2010, 327(5968): 1008-1010 (doi: 10.1126/science.1182570).
- Gomiero T. Soil degradation, land scarcity and food security: Reviewing a complex challenge. Sustainability (Switzerland), 2016, 8(3): article № 281 (doi: 10.3390/su8030281).
- Kopittke P.M., Menzies N.W., Wang P., McKenna B.A., Lombi E. Soil and the intensification of agriculture for global food security. Environment International, 2019, 132: article № 105078 (doi: 10.1016/j.envint.2019.105078).
- Di H.J., Cameron K.C. Nitrate leaching in temperate agroecosystems: sources, factors and mitigating strategies. Nutrient Cycling in Agroecosystems, 2002, 64(3): 237-256 (doi: 10.1023/A:1021471531188).
- Sebilo M., Mayer B., Nicolardot B., Pinay G., Mariotti A. Long-term fate of nitrate fertilizer in agricultural soils. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(45): 18185-18189. (doi: 10.1073/pnas.1305372110).
- Wang Y., Ying H., Yin Y., Zheng H., Cui Z. Estimating soil nitrate leaching of nitrogen fertilizer from global meta-analysis. Science of the Total Environment, 2019, 657: 96-102 (doi: 10.1016/j.scitotenv.2018.12.029).
- Tilman D., Cassman K.G., Matson P.A., Naylor R., Polasky S. Agricultural sustainability and intensive production practices. Nature, 2002, 418(6898): 671-677 (doi: 10.1038/nature01014).
- Castellano M.J., David M.B. Long-term fate of nitrate fertilizer in agricultural soils is not necessarily related to nitrate leaching from agricultural soils. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(8): E766 (doi: 10.1073/pnas.1321350111).
- Калиничева Е.Ю., Польшакова Н.В., Коломейченко А.С. Мелиорация сельскохозяйственных земель в России: состояние и перспективы развития. Вестник Орловского государственного аграрного университета. 2017, 3(66): 121-128.
- Ramankutty N., Foley J.A. Estimating historical changes in global land cover: Croplands from 1700 to 1992. Global Biogeochemical Cycles, 1999, 13(4): 997-1027 (doi: 10.1029/1999GB900046).
- Крышняя С.В. Феномен сахалинского крупнотравья: условия произрастания растений крупнотравного комплекса. Вестник Сахалинского музея, 2011, 17: 338-356.
- Бобровский М.В. Лесные почвы Европейской России. М., 2010.
- Смирнова О.В., Луговая Д.Л., Проказина Т.С. Модельная реконструкция восстановленного лесного покрова таежных лесов. Успехи современной биологии, 2013, 2: 164-177.
- Смирнова О.В., Шашков М.П., Коротков В.Н., Широков А.И. Лесные «острова» Южного Ямала. Природа, 2008, 12: 20-24.
- Смирнова О.В., Алейников А.А., Смирнов Н.С., Луговая Д.Л. Пионовая тайга. Природа, 2014, 2: 54-63.
- Тишков А.А. Актуальная биогеография как методологическая основа сохранения биоразнообразия. Вопросы географии, 2012, 134: 15-57.
- Мониторинг биологического разнообразия лесов России: методология и методы /Отв. ред. А.С. Исаев. М., 2008.
- Таранов С.А. Экологические и генетические особенности почв лесного пояса Горной Шории. В кн.: Лесные почвы горного окаймления юго-востока Западной Сибири (Восточный Алтай, Горная Шория, Салаир) /Отв. ред. Р.В. Ковалев. Новосибирск, 1974: 75-132.
- Бабенко А.С., Нефедьев П.С., Нефедьева Ю.С. Фауна и динамика населения двупарноногих многоножек (Diplopoda) черневой тайги Салаира. Вестник Томского государственного университета, 2009, 319:182-185.
- Fierer N., Strickland M.S., Liptzin D., Bradford M.A., Cleveland C.C. Global patterns in belowground communities. Ecology Letters, 2009, 12(11): 1238-1249 (doi: 10.1111/j.1461-0248.2009.01360.x).
- Delgado-Baquerizo M., Oliverio A.M., Brewer T.E., Benavent-González A., Eldridge D.J., Bardgett R.D., Maestre F.T., Singh B.K., Fierer N. A global atlas of the dominant bacteria found in soil. Science,2018, 359(6373): 320-325 (doi: 10.1126/science.aap9516).
- Lundberg D.S., Lebeis S.L., Paredes S.H., Yourstone S., Gehring J., Malfatti S., Tremblay J., Engelbrektson A., Kunin V., del Rio T.G., Edgar R.C., Eickhorst T., Ley R.E., Hugenholtz P., Tringe S.G., Dangl J.L. Defining the core Arabidopsis thaliana root microbiome. Nature, 2012, 488: 86-90 (doi: 10.1038/nature11237).
- Bates S.T., Berg-Lyons D., Caporaso J.G., Walters W.A., Knight R., Fierer N. Examining the global distribution of dominant archaeal populations in soil. The ISME Journal, 2011, 5: 908-917 (doi: 10.1038/ismej.2010.171).
- Martin V. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal, 2011, 17(1): 10-12 (doi: 10.14806/ej.17.1.200).
- Callahan B.J., McMurdie P.J., Rosen M.J., Han A.W., Johnson A.J.A., Holmes S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nature Methods, 2016, 13: 581-583 (doi: 10.1038/nmeth.3869).
- Quast C., Pruesse E., Yilmaz P., Gerken J., Schweer T., Yarza P., Peplies J., Glöckner F.O. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Research, 2013, 41(D1): D590-D596 (doi: 10.1093/nar/gks1219).
- Janssen S., McDonald D., Gonzalez A., Navas-Molina J.A., Jiang L., Xu Z.Z., Winker K., Kado D.M., Orwoll E., Manary M., Mirarab S., Knight R. Phylogenetic placement of exact amplicon sequences improves associations with clinical information. mSystems, 2018, 3: e00021-18 (doi: 10.1128/mSystems.00021-18).
- Bolyen E., Rideout J.R., Dillon M.R., Bokulich N.A., Abnet C.C., Al-Ghalith G.A., Alexander H., Alm E.J., Arumugam M., Asnicar F., Bai Y., Bisanz J.E., Bittinger K., Brejnrod A., Brislawn C.J., Brown C.T., Callahan B.J., Caraballo-Rodríguez A.M., Chase J., Cope E.K., Da Silva R., Diener C., Dorrestein P.C., Douglas G.M., Durall D.M., Duvallet C., Edwardson C.F., Ernst M., Estaki M., Fouquier J., Gauglitz J.M., Gibbons S.M., Gibson D.L., Gonzalez A., Gorlick K., Guo J., Hillmann B., Holmes S., Holste H., Huttenhower C., Huttley G.A., Janssen S., Jarmusch A.K., Jiang L., Kaehler B.D., Kang K.B., Keefe C.R., Keim P., Kelley S.T., Knights D., Koeste.r I, Kosciolek T., Kreps J., Langille M.G.I., Lee J., Ley R., Liu Y.X., Loftfield E., Lozupone C., Maher M., Marotz C., Martin B.D., McDonald D., McIver L.J., Melnik A.V., Metcalf J.L., Morgan S.C., Morton J.T., Naimey A.T., Navas-Molina J.A., Nothias L.F., Orchanian S.B., Pearson T., Peoples S.L., Petras D., Preuss M.L., Pruesse E., Rasmussen L.B., Rivers A., Robeson M.S., Rosenthal P., Segata N., Shaffer M., Shiffer A., Sinha R., Song S.J., Spear J.R., Swafford A.D., Thompson L.R., Torres P.J., Trinh P., Tripathi A., Turnbaugh P.J., Ul-Hasan S., van der Hooft J.J.J., Vargas F., Vázquez-Baeza Y., Vogtmann E., von Hippel M., Walters W., Wan Y., Wang M., Warren J., Weber K.C., Williamson C.H.D., Willis A.D., Xu Z.Z., Zaneveld J.R., Zhang Y., Zhu Q., Knight R., Caporaso J.G. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nature Biotechnology, 2019, 37: 852-857 (doi: 10.1038/s41587-019-0209-9).
- McMurdie P.J., Holmes S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE, 2013, 8(4): e61217 (doi: 10.1371/journal.pone.0061217).
- Love M.I., Huber W., Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biology, 2014, 15(12): 550 (doi: 10.1186/s13059-014-0550-8).
- Лойко С.В., Герасько Л.И., Кулижский С.П., Амелин И.И., Истигечев Г.И. Строение почвенного покрова северной части ареала черневой тайги юго-востока Западной Сибири. Почвоведение, 2015, 4: 410-423 (doi: 10.7868/S0032180X15040061).
- Орлов Д.С., Садовникова Л.К., Суханова Н.И. Химия почв. М., 2005.
- Базилевич Н.И., Титлянова А.А. Биологический круговорот на пяти континентах: азот и зольные элементы в природных надземных экосистемах. Новосибирск, 2008.
- Трофимов С.С. Экология почв и почвенные ресурсы Кемеровской области. Новосибирск, 1975.
- Achat D.L., Bakker M.R., Augusto L., Derrien D., Gallegos N., Lashchinskiy N., Milin S., Nikitich P., Raudina T., Rusalimova O., Zeller B., Barsukov P. Phosphorus status of soils from contrasting forested ecosystems in southwestern Siberia: effects of microbiological and physicochemical properties. Biogeosciences, 2013, 10: 733-752 (doi: 10.5194/bg-10-733-2013).
- Лойко С.В., Бобровский М.В., Амелин И.И. Высокотравная черневая тайга Западной Сибири: почвенные и палеоэкологические причины высокой продуктивности. Мат. докладов Всероссийской научно-практической конференции «Человек и природа — взаимодействие на особо охраняемых природных территориях», посвященной Году особо охраняемых природных территорий и Году экологии (Новокузнецк, 27-30 сентября 2017 года). Новокузнецк, 2017: 81-96.
- Смоленцев Б.А., Смоленцева Е.Н. Бурозёмы Кузнецкого Алатау, их свойства и разнообразие. Вестник Томского государственного университета. Биология, 2020, 50: 6-27 (doi: 10.17223/19988591/50/1).
- Корсунов В.М. Подзолистые почвы автономных ландшафтов средней тайги Приенисейской части Западной Сибири. В сб.: О почвах Сибири. Новосибирск, 1978: 122-131.
- Корсунова Т.М., Корсунов В.М. Экологический условия формирования специфического гумусового профиля подзолистых почв южной тайги Приенисейской равнины. В сб.: Генезис и география лесных почв. М., 1980: 85-104.
- Корсунов В.М., Ведрова Э.Ф., Игнатьева Л.Н. Генетико-лесоводственная характеристика почв Причулымья и перспективы их сельскохозяйственного использования в связи с развитием КАТЭКа. В сб.: Почвы зоны КАТЭКа. Красноярск, 1981: 99-113.
- Корсунов В.М., Ведрова Э.Ф. Оподзоленные почвы южной части Енисейской равнины. В сб.: География и картография лесных почв. Новосибирск, 1982: 66-88.
- Горожанкина С.М., Константинов В.Д. География тайги Западной Сибири. Новосибирск, 1978.
- Васенев И.И., Таргульян В.О. Ветровал и таежное почвообразование (режимы, процессы, морфогенез почвенных сукцессий). М., 1995.
- Васенев И.И. Почвенные сукцессии. М., 2008.
- Урусевская И.С., Хохлова О.С., Соколова Т.А. Влияние почвообразующих пород на дифференциацию почв и почвенного покрова северной части Приволжской возвышенности. Почвоведение, 1992, 8: 22-37.
- Пономарева В.В. Теория подзолообразовательного процесса. Биохимические аспекты /Под ред. М.М. Кононова. Казань, 1964.
- Пономарева В.В., Плотникова Т.А. Гумус и почвообразование (методы и результаты изучения). Л., 1980: 222.
- Pershina E.V., Ivanova E.A., Korvigo I.O., Chirak E.L., Sergaliev N.H., Abakumov E.V., Provorov N.A., Andronov E.E. Investigation of the core microbiome in main soil types from the East European plain. Science of the Total Environment, 2018, 631-632: 1421-1430 (doi: 10.1016/j.scitotenv.2018.03.136)
- Розенберг Г.С. Несколько слов об индексе разнообразия Симпсона. Бюллетень Самарская Лука, 2007, 16(3-21): 581-584.
- Sangwan P., Chen X., Hugenholtz P., Janssen P.H. Chthoniobacter flavus gen. nov., sp. nov., the first pure-culture representative of subdivision two, Spartobacteria classis nov., of the phylum Verrucomicrobia. Applied and Environmental Microbiology, 2004, 70(10): 5875-5881 (doi: 10.1128/AEM.70.10.5875-5881.2004).
- Kant R., van Passel M.W., Palva A., Lucas S., Lapidus A., Glavina del Rio T., Dalin E., Tice H., Bruce D., Goodwin L., Pitluck S., Larimer F.W., Land M.L., Hauser L., Sangwan P., de Vos W.M., Janssen P.H., Smidt H. Genome sequence of Chthoniobacter flavus Ellin428, an aerobic heterotrophic soil bacterium. Journal of Bacteriology, 2011, 193(11): 2902-2903 (doi: 10.1128/JB.00295-11).