








УДК 635.646:631.52:575.222.7:581.481:57.086.83
ПОЛУЧЕНИЕ МЕЖВИДОВЫХ ГИБРИДОВ БАКЛАЖАНА МЕТОДОМ ЭМБРИОКУЛЬТУРЫ
В.М. ВЕРБА, М.И. МАМЕДОВ, О.Н. ПЫШНАЯ, Т.П. СУПРУНОВА, Н.А. ШМЫКОВА
Проводили межвидовые реципрокные скрещивания между тремя видам баклажана Solanum melongena, S. aethiopicum и S. integrifolium. С использованием культуры зародышей in vitro получили межвидовые гибриды следующих комбинаций: S. melongena x S. aethiopicum, S. melongena x S. integrifolium, S. aethiopicum x S. melongena, S. aethiopicum x S. integrifolium и S. integrifolium x S. melongena. Определена стадия развития зародыша, на которой он наиболее пригоден для изоляции, оптимизирован состав питательных сред для развития зародыша и укоренения проростков.
Ключевые слова: баклажан, межвидовая гибридизация, эмбриокультура.
Межвидовая гибридизация имеет важное практическое значение как прием расширения генетической изменчивости культурных растений, позволяющий получать большое разнообразие исходного материала для создания новых сортов и гибридов.
Вид Solanum aethiopicum L. может служить источником генов устойчивости к Fusarium oxysporum (1), F. solani (2), Phytophtora parasitica (3), S. aethiopicum sp. Gilo — как источник устойчивости к грибу Phomopsis vexans (4). S. integrifolium использован при передаче устойчивости к стеблевому и цветочному долгоносику, вирусной мелколиственности и плодовой гнили (6). Указывается на возможность переноса генов устойчивости к двупятнистому паутинному клещу от S. macrocarpon (син. S. integrifolium) к S. melongena (7). Отмечено, что S. aethiopicum и S. macrocarpon меньше поражались бактериальным увяданием (5).
При межвидовых скрещиваниях трудности связаны с нескрещиваемостью исходных форм, стерильностью и нарушением формообразования у межвидовых гибридов. Так, S. aethiopicum и S. integrifolium при скрещивании с S. melongena проявляли несовместимость на постзиготическом и постэмбриональном этапах (8).
Целью нашей работы было получение межвидовых гибридов баклажана с использованием метода эмбриокультуры.
Методика. Материалом для исследований служили селекционные и коллекционные образцы баклажана из генофонда лаборатории селекции и семеноводства пасленовых культур Всероссийского НИИ селекции и семеноводства овощных культур (ВНИИССОК) — S. melongena (линии Л-Алмаз и Л-Бриллиант), S. aethiopicum и S. integrifolium. Скрещивания проводили в теплице на растениях, выращиваемых по технологии малообъемной гидропоники, по полной диаллельной схеме.
Опыление с изоляцией опыленных цветков выполняли с 1000 до 1200. Плоды S. aethiopicum и S. integrifolium собирали на 20-е, 25-е, 30-е, 35-е, 40-е и 45-е сут после опыления (в зависимости от комбинации скрещивания и с учетом биологических особенностей развития зародышей исходных видов), S. melongena — на 20-е, 23-е, 27-е, 31-е, 35-е и 40-е сут. Семена, извлеченные из плодов, обеззараживали 50 % коммерческим препаратом «Белизна» (гипохлорит натрия) в течение 10 мин с последующим многократным промыванием стерильной дистиллированной водой.
Для культивирования зародышей использовали среду Мурасиге и Скуга (МС) (9) с добавлением регуляторов роста — тидиазурона (ТДЗ) (0,1 мг/л) и нафтилуксусной кислоты (НУК) (10 мг/л) (10, 11). Экспланты культивировали в стеклянных сосудах объемом 100 мл с 20 мл агаризованной среды. Условия культивирования: освещенность — 1,5-2,0 тыс. лк, фотопериод — 14 ч, температура — 20-22 °С.
Проростки длиной 0,3-1,0 см (10-15-суточные) переносили для укоренения на агаризованную среду Мурасиге и Скуга с половинным содержанием основных компонентов (1/2 МС) без добавления регуляторов роста. После появления 1-й пары настоящих листьев и хорошо развитых корней (15-25-е сут) растения пересаживали из сосудов в торфяные горшочки с почвосмесью под пластиковые стаканы с перфорациями.
Для определения оптимальных сроков изоляции гибридных зародышей изучали стадии развития зародышей родительских видов и гибридов, полученных с их участием. Учитывали следующие стадии развития: глобулярный, сердечковидный и торпедовидный эмбриоид, почти сформированный и сформированный зародыш. В каждой гибридной комбинации измеряли 15-20 зародышей с помощью окуляр-микрометра стереомикроскопа (STEMI-SV8, «OPTON», Германия).
Наряду с морфологическим описанием выполняли тест на гибридность полученных межвидовых гибридов баклажана с помощью полимеразной цепной реакции (ПЦР) с использованием ISSR (inter simple sequence repeat — полиморфизм фрагментов ДНК, фланкированных микросателлитными последовательностями) и IRAP (inter retrotransposon amplified polymorphism — полиморфизм фрагментов ДНК, фланкированных инвертированными терминальными участками ретротранспозона) праймеров.
Тотальную ДНК родительских линий и гибридных растений баклажана выделяли из молодых листьев, как описано (12), с незначительными модификациями, а именно: растительные пробы растирали непосредственно в экстракционном буфере, после чего пробирки инкубировали в термостате 20 мин при температуре 65 °С. Амплификацию ДНК проводили в реакционной смеси объемом 25 мкл, содержащей 1xTaq-буфер (50 % глицерол, 20 мM Трис-HCl, pH 8,0, 100 мM KCl, 0,1 мM ЭДТА, 1мM 1,4-дитио-DL-треитол, 0,5 % Твин 20) из соответствующего набора реагентов, 1,5 мМ MgCl2, 0,2 мМ каждого dNTP, 20 пмоль праймера, 0,2 ед. Taq-по-лимеразы (UAB «Fermentas», Литва) и 10-50 нг ДНК. Использовали термоциклер MyCycler («Bio-Rad», США) в следующем режиме: предварительная денатурация — 4 мин при 94 °С; денатурация — 40 с при 94 °С; отжиг праймеров — 40 с при 50-62 °С (в зависимости от праймеров); синтез ДНК — 1 мин при 72 °С, 30 циклов; заключительная элонгация — 10 мин при 72 °С. Контролем служила ПЦР-смесь, не содержащая ДНК. Продукты амплификации разделяли электрофорезом в 1,5 % агарозном геле с 0,5xТВЕ-буфером (0,89 М Трис-HCl, 0,89 М борная кислота, 20 мМ ЭДТА, рН 7,5), окрашивали бромистым этидием; результаты фотодокументировали с помощью системы CN-1500 Darkroom («Vilber Lourmat», Франция). Для амплификации использовали восемь ISSR-праймеров и пять IRAP-праймеров, синтезированных в ЗАО «Синтол» (Россия).
Стандартным вариационно-статистическим методом определяли предельную ошибку выборочной средней с помощью программы Microsoft Excel (13).
Результаты. Учитываемые стадии развития зародышей у всех трех видов баклажана проходили в период с 20-х до 45-х сут с даты опыления (рис. 1). Наиболее быстро зародыш формировался у вида S. aethiopicum, медленно — у вида S. integrifolium(табл. 1).
| 1. Стадии развития зародышей у изученных видов рода SolanumL. | |||
Вид |
Время после опыления, сут |
Длина зародыша, мм |
Стадия развития зародыша |
S. melongena |
20-е |
0,032±0,001 |
Глобулярный эмбриоид |
23-и |
0,100±0,010 |
Сердечковидный эмбриоид |
|
27-е |
0,600±0,060 |
Торпедовидный эмбриоид |
|
31-е |
1,200±0,060 |
Торпедовидный эмбриоид |
|
35-е |
3,800±0,090 |
Почти сформированный зародыш |
|
40-е |
4,300±0,090 |
Сформированный зародыш |
|
S. aethipicum |
20-е |
0,080±0,007 |
Глобулярный эмбриоид и сердечковидный эмбриоид |
25-е |
2,200±0,300 |
Торпедовидный эмбриоид/почти сформированный зародыш |
|
30-е |
4,100±0,050 |
Сформированный зародыш |
|
S. integrfolium |
20-е |
0,070±0,005 |
Глобулярный эмбриоид |
25-е |
0,150±0,030 |
Сердечковидный эмбриоид |
|
30-е |
1,100±0,100 |
Торпедовидный эмбриоид |
|
35-е |
1,800±0,200 |
Торпедовидный эмбриоид |
|
40-е |
3,900±0,200 |
Почти сформированный зародыш |
|
45-е |
4,500±0,100 |
Сформированный зародыш |
|
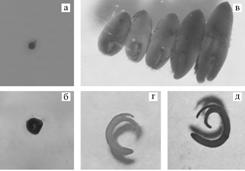 |
Рис. 1. Стадии развития зародышей у представителей рода Solanum L.: а — глобулярный эмбриоид, б — сердечковидный эмбриоид, в — торпедовидные эмбриоиды, г — почти сформированный зародыш, д — сформированный зародыш. |
Наблюдение за развитием зародышей in vitro у гибридов между S. melongena и дикими видами показало, что стадия полностью сформированного зародыша наступала несколько раньше, чем у материнского компонента S. melongena. У гибрида S. melongena x S. aethiopicum зародыш полностью формировался на 35-е сут после опыления и достигал 4,4-4,5 мм в длину. У гибрида S. melongena x S. integrifolium зародыш полностью формировался на 40-е сут (так же, как у S. melongena) при длине 4,2-4,3 мм.
В других гибридных комбинациях, где материнским компонентом служили дикие виды, развитие зародыша происходило в сроки, характерные для исходных материнских форм. Размеры зародышей также соответствовали их размерам у исходных видов.
| 2. Характер развития зародышей у межвидовых гибридов (Solanum melongena — материнский компонент) через 14-28 сут после начала культивирования in vitro на среде с разными регуляторами роста | |||
Гибридная |
Время после опыления, сут |
Регулятор роста |
|
ТДЗ (0,1 мг/л) |
НУК (10 мг/л) |
||
S. melongena (Л-Бриллиант) x S. integrifolium |
25-е |
Посадка в зародышевых мешках, развития нет |
Посадка в зародышевых мешках, развития нет |
30-е |
Ненормальные семядоли, толстый главный корень без придаточных |
Слабый белый каллус с зеленой почкой в центре |
|
35-е |
Развитие нормальных побегов, адвентивные почки на гипокотиле |
Слабое развитие, разрастание белой паренхимной ткани на гипокотиле |
|
40-е |
Развитие нормальных побегов, начало укоренения, адвентивные почки на гипокотиле |
Белый и слабый зеленый каллус с почками |
|
S. melongena (Л-Бриллиант) x S. aethiopicum |
20-е |
Посадка в зародышевых мешках, развития нет |
Посадка в зародышевых мешках, развития нет |
25-е |
Посадка в зародышевых мешках, развития нет |
Посадка в зародышевых мешках, развития нет |
|
30-е
|
Развитие нормальных побегов
|
Развитие побегов с корнями, мощный белый каллус |
|
35-е |
Развитие нормальных побегов, на некоторых зеленый каллус с почками |
Белый каллус, ненормально развитые семядоли, побегов и корней нет |
|
S. melongena (Л-Алмаз) x S. integrifolium |
25-е |
Посадка в зародышевых мешках, развития нет |
Посадка в зародышевых мешках, развития нет |
30-е |
Развитие побегов, ненормальные семядоли и мощный центральный корень |
Белый каллус со слабым зеленым побегом в центре |
|
35-е |
Развитие побегов, адвентивные почки на гипокотиле |
Развитие слабого побега, разрастание белой паренхимной ткани, белый каллус |
|
40-е |
Развитие нормальных побегов, начало укоренения, адвентивные почки на гипокотиле |
Белый каллус с почками |
|
S. melongena (Л-Алмаз) x S. aethiopicum |
20-е |
Посадка в зародышевых мешках, развития нет |
Посадка в зародышевых мешках, развития нет |
25-е |
Посадка в зародышевых мешках, развития нет |
Посадка в зародышевых мешках, развития нет |
|
30-е |
Развитие нормальных побегов |
Белый каллус, ненормально развитые листовые розетки без гипокотиля |
|
35-е |
Развитие нормальных побегов |
Белый каллус, слабые ненормально развитые листовые розетки |
|
П р и м е ч а н и е. TДЗ — тидиазурон, НУК — нафтилуксусная кислота. |
|||
У всех трех видов зародыши лучше развивались на среде МС + ТДЗ (0,1 мг/л). В полноценные растения с нормально развитыми семядолями и корневой системой развивались 30-40-суточные зародыши. На среде МС + НУК (10 мг/л) зародыши, как правило, формировали белый рыхлый каллус с ненормально развитыми семядолями без стебля и с сильной корневой системой (либо, наоборот, она отсутствовала). Из таких зародышей не формировались нормальные растения.
| А |
|
Б |
|
Рис. 2. Развитие зародышей (возраст 35 сут) гибрида Solanummelongena (Л-Бриллиант) Í S. aethiopicumчерез 7-14 сут после пересадки на индукционную среду: А — среда МС + тидиазурон (0,1 мг/л), Б — среда МС + нафтилуксусная кислота (10 мг/л). |


Характер развития зародышей у всех гибридных комбинаций и исходных видов был схож (табл. 2, рис. 2, А). На среде с НУК (10 мг/л) 30- и 35-суточные зародыши образовывали белый рыхлый каллус с ненормальными семядолями, без корней (рис. 2, Б).
В комбинации S. melongena x S. integrifolium зародыши развивались в нормальные растения только с 35-суточного возраста на среде МС с ТДЗ (0,1 мг/л). Учет выживаемости на этой среде показал, что у 30-40-суточных зародышей (торпедовидный эмбриоид—полностью сформировавшийся зародыш) показатель составил в среднем 86,4 %, у 20-25-суточных (глобулярный эмбриоид—торпе-довидный зародыш) — только 7,9 %. Исключение составила комбинация S. melongena (Л-Бриллиант) x S. aethiopicum (35 сут), в которой из зародышей развивались сильные побеги и корни, а также белый каллус.
В среднем доля укоренившихся и пригодных к пересадке в почву растений для зародышей, перенесенных со среды с добавлением ТДЗ (0,1 мг/л) и НУК (10 мг/л), составила соответственно 69,0 и 9,6 %. Частота укоренения у зародышей в возрасте 35 сут и более в большинстве комбинаций превышала 80,0 %.
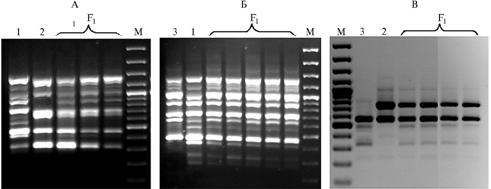
Наиболее информативными из протестированных оказались ISSR-праймер (CAG)5 и IRAP-праймер AACGAGGGGTTCGAGGCC, с которыми амплифицировалось наибольшее число полиморфных фрагментов ДНК. Эти праймеры были использованы для изучения чистоты гибридности растений, полученных при межвидовых скрещиваниях. В спектрах амплифицированных ISSR- и IRAP-фрагментов индивидуальных растений F1 присутствовали как отцовские, так и материнские компоненты (рис. 3). Результаты молекулярного анализа трех гибридных комбинаций подтвердили гибридную природу всех индивидуальных растений F1, полученных с использованием метода эмбриокультуры in vitrо.
|
Рис. 3. Электрофореграмма продуктов ПЦР-амплификации с помощью ISSR (А и Б) и IRAP (В) праймеров у трех родительских форм баклажана — Solanum integrifolium (1), S. melongena (Л-Алмаз) (2), S. aethiopicum (3) и их межвидовых гибридов F1, полученных с использованием метода эмбриокультуры in vitrо. М — маркер молекулярных масс (GeneRuler, 100 bp DNA Ladder, UAB «Fermentas», Литва). |
Таким образом, зародыши видов Solanum melongena, S. aethiopicum и S. integrifolium проходят основные стадии развития (от глобулярного эмбриоида до полностью сформировавшегося зародыша) за период от 20-х до 45-х сут после опыления цветка материнского растения. Быстрее, чем у других изученных видов, развивается зародыш у S. aethiopicum, медленнее — у S. integrifolium. Для изоляции наиболее пригодны хорошо развитые торпедовидные зародыши (> 2 мм) на более поздних стадиях развития (30-45-е сут). Оптимальная индукционная среда для развития зародышей на первом этапе — среда Мурасиге и Скуга с добавлением тидиазурона (0,1 мг/л), на которой период развития зародыша не должен превышать 7-14 сут. Для укоренения развившихся побегов подходит среда Мурасиге и Скуга с половинным количеством основных компонентов без добавления регуляторов роста. В этих условиях в короткие сроки были получены гибридные формы в следующих комбинациях скрещивания: S. melongena x S. aethiopicum, S. melongena x S. integrifolium, S. aethiopicum x S. melongena, S. aethiopicum x S. integrifolium и S. integrifolium x S. melongena.
Л И Т Е Р А Т У Р А
1. Y a m a k a w a K., M o c h i z u k i H. Nature and inheritance of Fusarium wilt resistance in eggplant cultivars and related wild Solanum species. Bull. Veg. Orn. Crops Res. Stn, 1979, 6: 19-27.
2. D a u n a y M.C., L e s t e r R.N., L a t e r r o t H. The use of wild species for the genetic improvement of Brinjal eggplant (Solanum melongena) and tomato (Lycopersicon esculentum). In: Solanaceae III: taxonomy, chemistry, evolution /J.C. Hawkes, R.N. Lester, M. Nee, N. Estrada (eds.) /Royal Botanic Gardens Kew and Linnean Soc., London, 1991, 27: 389-413.
3. B e y r i e s A., L e f o r t L., B o u d o n J. Contribution à la recherche du épérissement de l’aubergine (Solanum melongena L.) aux Antilles françaises. I. Etude de la virulence d’isolats de Phytophtora nicotianae var. parasitica (Dastur.) artificielle. Stn. Pathologie végétale, INRA-ENSA. Montpellier, France, 1984.
4. K a l d a T.S., S w a r u p V., C h o u d h u r y B. Resistance to Phomosis blight in eggplant. Veg. Sci., 1977, 4: 90-101.
5. M o n m a S., S a t o T., M a t s u n a g a T. Evaluation of resistance to bacterial fusarium and verticillium wilt in eggplant and eggplant-related species collected in Ghana. Caps. and Egg. Newslett., 1996, 15: 71-72.
6. R a o G., B a k s h S. Relationship between Solanum melongena L. and Solanum integrifolium Poir. Indian J. Genet. Plant Breed, 1981, 41(1): 46-53.
7. S c h a f f D.A., J e l e n k o v i c G., B o y e r C.D., P o l l a c k B.L. Hybridization and fertility of hybrid derivatives of Solarium melongena L. and Solanum macrocarpon L. Theor. Appl. Genet., 1982, 62(2): 149-153.
7. Б у х а р о в А.Ф., Б у х а р о в а А.Р Генетический потенциал диких сородичей баклажана и перспективы вовлечения их в селекционный процесс. В сб.: Селекция и семеноводство овощных культур в XXI веке. Т. I. М., 2000: 146-148.
8. M u r a s h i g e T., S k o o g F.A. A revised medium for rapid growth and bioassays with tobacco tissue culture. Phys. Plant, 1962, 15: 473-497.
9. R o t i n o G.L., F a l a v i g n a A., F i u m e F., N e r v o G., R e s ta i n o F. Possibility of eggplant (Solanum melongena L.) improvement through in vitro techniques. Genet. Agraria, 1987, 41: 314-315.
10. M a r i a n i P. Eggplant somatic embryogenesis combined with synthetic seed technology. Capsicum Newslett., 1992, 289-294.
11. E d w a r d s S.K., J o h n s t o n e C., T h o m p s o n C. A simple and rapid method for the preparation of plant genomic DNA for PCR analyses. Nuc. Acids Res., 1991, 19(6): 1349.
12. Д о с п е х о в Б.А. Методика полевого опыта. М., 1985.
ISOLATION OF EGGPLANT INTERSPECIFIC HYBRIDS BY THE METHOD OF EMBRYO CULTURE
V.M. Verba, M.I. Mamedov, O.N. Pyshnaya, T.N. Suprunova, N.A. Shmykova
The authors made interspecific reciprocal crossings between three species of eggplant: Solanum melongena, S. aethiopicum and S. integrifolium. By the using of embryo culture in vitro the authors obtained the interspecific hybrids of following combinations: S. melongena x S. aethiopicum, S. melongena x S. integrifolium, S. aethiopicum x S. melongena, S. aethiopicum x S. integrifolium и S. integrifolium x S. melongena. The stage of embryonic development, optimal for isolation, was determined, the composition of nutrient media for embryo development and rootage of seedlings was optimized.
Key words: eggplant, interspecific hybridization, embryo culture.
ГНУ Всероссийский НИИ селекции и семеноводства
овощных культур Россельхозакадемии, |
Поступила в редакцию |




