








УДК 633.8:581.144:577.175.19:58.056
РОСТ И БИОСИНТЕЗ ЭКДИСТЕРОИДОВ У ЛЕВЗЕИ САФЛОРОВИДНОЙ ПОД ВЛИЯНИЕМ ЭДАФИЧЕСКИХ ФАКТОРОВ
Н.П. ТИМОФЕЕВ
Исследовали ростовые процессы у левзеи сафлоровидной Rhaponticum carthamoides (Willd.) Iljin в условиях Архангельской области и их связь с биосинтезом экдистероидов. Установлены факторы внешней среды, лимитирующие рост побегов (диапазон ростовых реакций на освещенность, температуру и влажность), устойчивость к низким и отрицательным температурам, а также влияние стресса на синтез и накопление экдистероидов в листьях.
Ключевые слова: левзея сафлоровидная, факторы внешней среды, рост побегов, стресс, биосинтез экдистероидов.
Фармацевтические средства и биологически активные добавки на основе левзеи сафлоровидной Rhaponticum carthamoides (Willd.) Iljin (Leuzea, рапонтикум сафлоровидный, маралий корень) используются при лечении сердечно-сосудистых и раковых заболеваний, в технологиях иммунокоррекции, жизнеобеспечения и защиты человека в экстремальных условиях, в агропромышленном комплексе (1-3). Выделяемые из растения экдистероиды служат объектами и инструментами исследований по клеточной биологии и молекулярной генетике (экдизон-индуцированные системы экспрессии генов), применяются в профессиональном спорте (препараты Экдистен и их аналоги на основе экдистерона), при разработке экологически безопасных инсектицидов (4-7).
Известно, что после первоначального синтеза и накопления во взрослых листьях фитоэкдистероиды (ФЭС) перемещаются в интенсивно растущие ткани и органы (8-9), причем эти процессы непрерывны (10). Условия внешней среды (температура, длина дня, освещенность, стресс при засухе и заморозках), влияя на рост и направленность метаболических процессов в растениях, способны изменять биосинтез и накопление целевых веществ в лекарственном сырье. До настоящего времени факторы, регулирующие образование ФЭС, не исследованы (за исключением воздействия спектра освещения на содержание экдистероидов в листьях растений родов Rhaponticum и Lychnis в условиях фитотрона) (11).
Нашей целью было изучение роли основных эдафических факторов (освещенность, высокие и низкие температуры, влажность, водный стресс) в регуляции роста побегов и биосинтеза экдистероидов у Rhaponticum car-thamoides.
Методика. Объектом изучения былидве разновозрастные агропопуляции R. carthamoides (площадь 1 и 4 га), расположенные в подзоне средней тайги европейского Северо-Востока (62° с.ш., 47° в.д.; г. Котлас-Коряжма, Архангельская обл.). Подробные характеристики объекта и природно-климатические условия описаны в предыдущих публикациях (10, 12). В целом климат в подзоне умеренно-прохладный, средняя температура января равна -14,3 °С, июля — +17,4 °С, сумма температур свыше +5/+10 °С — 1936/1577 °С, свыше 15 °С — 911 °С. Длина светового дня в мае—июне равна 16-20 ч; коэффициент увлажнения — 1,5 (13). Почвы участков супесчаные дерново-подзолистые на водно-ледниковых двучленных отложениях, средней окультуренности (pH 6,4-6,5; содержание гумуса — 1,5-3,6 %; K2O — 7,1-12,3; P2O5 — 18,0-31,2 мг/100 г; MgO — 1,0-1,4 мг-экв/100 г).
Норма высева семян в опыте составляла 3 кг/га (собственные семена). Минеральные и органические удобрения, химические средства защиты растений не применялись. Портативными цифровыми приборами фиксировали температуру и относительную влажность воздуха (PDT 300, «COMARK Div. of Fluke Electronics», США), а также освещенность на уровне травостоя (люксметр ELVOS LM 1010, «Elvos GmbH», Германия; диапазон измерений 0-200 тыс. лк). Влажность почвы определяли в корнеобитаемом слое 1-25 см (усредненные образцы, отобранные в 6-10 точках по диагонали участка, которые доводили до воздушно-сухого состояния, а затем досушивали при 130 °С в течение 30 мин).
Среднесуточный прирост фиксировали у 15-20 типичных случайных особей. Динамику роста у каждой особи учитывали по высоте наиболее развитых побегов — как вегетативных (розеточных с укороченными междоузлиями), так и генеративных (удлиненных репродуктивных). Часовой прирост побегов измеряли в периоды отрастания и бутонизации у шести растений на микроделянках в течение 9 сут и сопоставляли с показателями освещенности, температуры и влажности.
Изменчивость содержания ФЭС отслеживали во взрослых розеточных листьях вегетативных побегов. Образцы отбирали через каждые 3-6 сут (по два листа от 20-25 растений), высушивали при комнатной температуре и формировали средний образец методом квартования. Обращенно-фазовую высокоэффективную жидкостную хроматографию (ВЭЖХ) ФЭС выполняли в биохимической лаборатории Ботанического сада Института биологии Коми НЦ УрО РАН (г. Сыктывкар) (14). Содержание суммы ФЭС пересчитывали на абсолютно-сухое вещество.
Математическую обработку данных проводили стандартными методами статистики. При оценке результатов использовали значения среднего арифметического (x), коэффициенты корреляции (r) и уровни их достоверности (r), а также сглаженные (приближенные) кривые аппроксимации экспериментальных данных 4-й степени с определением достоверности аппроксимации (R2на рисунках).
Результаты. Р а з в и т и е R. c a r t h a m o i d e s в р а з л и ч
н ы х
п р и р о д н ы х з о н а х. Данные о культивировании R. carthamoides в различных регионах указывают, что теплообеспеченность и плодородие почвы не влияют на формирование продуктивности растений. Например, на географических широтах 52° (Прибалтика), 55-57° (Томская—Новосибирская области) и 61-62° (Карелия—Коми Республика) сумма активных температур свыше 10 °С различалась в 1,5-1,8 раза (от 2100-2300 до 1400-1500 °С), содержание гумуса — в 2-3 раза (от 5-9 % до 2-3 %) (15), в то время как продуктивность фитомассы левзеи сафлоровидной в этих зонах на 3-4-й годы жизни не различалась и варьировала в пределах от 55-60 до 81-86 г для надземных и от 58-60 до 76-91 г для подземных органов (1, 12).
Наше 4-летнее сравнительное возделывание R. carthamoides в условиях агропопуляций Центральной Польши (53° с.ш.) и на европейском Северо-Востоке России (62° с.ш.), проведенное по единой методике, показало, что процессы роста и развития растений в двух этих климатических зонах идентичны — среднее число побегов, их высота, ширина листовых пластинок были одинаковыми для одних и тех же возрастных состояний (16).
Величина сухой фитомассы надземных органов с 1-го по 4-й год жизни в условиях Польши составила по годам соответственно 0,23; 7,00; 17,50 и 47,80 г. Такая же динамика прироста наблюдалась в агропопуляции на европейском Севере — соответственно 0,26; 6,20; 16,40 и 56,80 г. Фитомасса подземных органов (корни и корневища) на 3-4-й год равнялась соответственно 16,80-29,50 и 11,90-38,20 г. Содержание ФЭС во взрослых листьях в условиях Польши составило 0,036 % у имматурных растений, 0,18 % — у виргинильных, на европейском Севере — соответственно 0,17-0,19 и 0,22 %.
Д и а п а з о н р о с т о в ы х р е а к ц и й н а т е м п е р а т у р у,
о с в е щ е н н о с т ь и в л а ж н о с т ь п о ч в ы. У с т о й ч и в о с т ь
к о т р и ц а т е л ь н ы м т е м п е р а т у р а м.Начало массового отрастания вегетативных побегов в зависимости от погодных условий в годы наблюдений (1990-2008 годы) отмечали в сроки между 17 апреля и 12 мая, видимый рост генеративных побегов — через 5-7 сут. В этот период часты возвраты холодов с выпадением снега и многократные заморозки. Отрицательные температуры (до -5 °С) R. carthamoides выдерживает без видимых последствий. При температурах -8…-10 °С повреждалась апикальная зона роста (1,5×2,0 см) у листовых органов (верхушки листовых пластинок), через 4-5 сут поврежденные участки восстанавливались, заменяясь новообразованной тканью. У генеративных побегов при кратковременных ночных заморозках (-7…-10 °С) апикальные части (соцветия) повреждались необратимо, чернели и отмирали. Осенние заморозки (-2…-3 °С в сентябре—октябре) не причиняли вреда вегетирующим розеточным листьям.
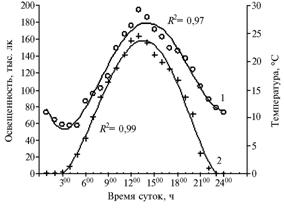 |
Рис. 1. Динамика суточных показателей температуры (1) и освещенности (2) в период отрастания Rhaponticumcarthamoides. R2 — достоверность аппроксимации (Архангельская обл., подзона средней тайги, 2003-2004 годы). |
С у т о ч н ы й ц и к л
п а р а м е т р о в в н е ш-
н е й с р е д ы. Ростовые процессы весьма чувствительны к колебаниям освещенности, температуры и влажности (17). На растениях 3-го и 9-го годов жизни в фазу отрастания (II декада мая, длина дня 17-18 ч) мы исследовали динамику часового прироста побегов в зависимости от суточного цикла факторов внешней среды. Установили, что в безоблачную погоду в ночное время полного отсутствия освещенности не наблюдается, минимальная величина рассеянного света приходится на 100-200 и составляет 10-20 лк, с 300-400 до 1200-1400 освещенность возрастает до максимума (156-164 тыс. лк), после чего снижается и в 2100-2200 составляет 24-7 тыс. лк (рис. 1). При легкой и средней облачности освещенность в полдень снижалась в 1,5-2,0 раза (до 110-70 тыс. лк), утром и вечером — в 3,0-4,0 раза (до 30-40 тыс. лк в 900 и 1800).
Минимум температурной кривой (6-10 °С) приходился на раннее утро (300-500), максимум (28-30 °С в травостое) — на 1300-1400; в послеполуденное время температура постепенно снижалась (до 20 °С в 1900, до 13-11 °С — в 2200-2400), то есть следовала за освещенностью (см. рис. 1) (кривая несколько сдвигалась вправо, проявляя инерционность). Корреляционная зависимость между температурой и освещенностью в ночное время отрицательная, с 500-600 — положительная, в дневное время (с 900-1000 утра) — прямолинейная (r = 0,94-0,97; r = 0,999).
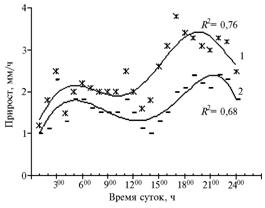 |
Рис. 2. Часовой прирост вегетативных побегов у Rhaponticumcarthamoides в суточном цикле на 3-й (1) и 9-й (2) годы жизни. R2— достоверность аппроксимации (Архангельская обл., подзона средней тайги, 2003-2004 годы). |
Динамика изменения относительной влажности воздуха противоположна динамике освещения и температуры; корреляция между ними отрицательная и высокая (r = 0,999) — r= -0,74… -0,82 для влажности и величины солнечной радиации, r= -0,85… -0,90 для влажности и температуры.
Наибольшие показатели влажности (88-90 %) приходятся на раннее утро (200-800), наименьшие (31-33 %) — на полуденное время — 1400-1800. В период с 900 до 1200 влажность постепенно уменьшается с 80-83 до 40-42 %, с 1900 до 2300 — увеличивается с 35 до 80 %.
Ч а с о в о й п р и р о с т п о б е г о в в с у т о ч н о м ц и к л е.
Характер кривых, описывающих прирост вегетативных побегов (рис. 2), оказался идентичен у растений разного возраста, но величина абсолютных показателей у молодых 3-летних (виргинильных) была в 1,5-1,7 раза меньше, чем у 9-летних взрослогенеративных. Ростовые процессы в условиях высокой влажности не прекращались даже в ночное время: прирост составлял около 1 мм/ч у 3-летних особей, 1,2-1,8 мм/ч — у 9-летних. В условиях сумеречного рассеянного освещения в 300 (2 тыс. лк) он возрастал в 2 раза, к 900-1000 на фоне яркой освещенности наблюдалась тенденция к его замедлению (см. рис. 2). Экстремальные значения солнечной радиации (150-160 тыс. лк) и температуры (28-30 °С) в 1300-1400 ингибировали прирост побегов до значений, сопоставимых с ночными (1,0-1,1 мм/ч у 3-летних и 1,6-1,8 мм/ч у 9-летних растений).
Наибольший прирост (2,0-2,6 и 3,0-3,8 мм/ч соответственно для 3- и 9-летних растений) в суточном цикле отмечали в вечернее время — с 1600-1700 до 2200-2300, когда освещенность снижалась со 120 тыс. лк до 7 тыс. лк, температура — с 25 до 10-12 °С, а влажность воздуха возрастала с 30 до 70-80 %.
В целом полученные данные согласуются с результатами модельных опытов Т.К. Головко с соавт. (18), показавших, что у R. carthamoides величина светового компенсационного пункта (баланс между фотосинтезом и дыханием по CO2) составила 4,4-6,7 Вт/м2, а интенсивность радиации приспособления равна 30-50 Вт/м2, или около 5-10 % от максимальной инсоляции.
Анализ кривой аппроксимированных значений корреляции между приростом побегов и условиями внешней среды свидетельствует, что 3-летние растения сильнее нуждались в свете, чем 9-летние (R2 = 0,84 против R2 = 0,54 в суточном цикле). Реализация потенциала 9-летних особей зависела прежде всего от оптимальной влажности и температуры (соответственно R2 = 0,96 и R2 = 0,92), что может быть связано с возрастанием роли корневой системы в обеспечении жизнедеятельности (12).
С р е д н е с у т о ч н ы й п р и р о с т. Исследование развития розеточных побегов R. carthamoides в зависимости от температуры и влажности проведено у взрослогенеративных 6-летних растений (табл. 1). В начале отрастания наблюдались значительные суточные перепады температуры — от 15-18 °С в дневное время до 2-7 °С в вечерние и ночные часы. Влажность была в пределах 56-87 %. Среднесуточный прирост вегетативных побегов в этот период составляет 2,1 см. На фоне пониженных дневных температур (7-10 °С) при близких параметрах влажности (62-73 %) прирост снижается, но незначительно — до 1,7 см/сут.
| Среднесуточный прироствегетативных побегов у Rhaponticumcarthamoides по датам сезона вегетации в связи с температурой и влажностью воздуха (Архангельская обл., 6-й год жизни растений, 1995 год) | ||||||||
Показатель |
Дата |
|||||||
24 апреля |
14 мая |
18 мая |
28 мая |
06 июня |
12 июня |
18 июня |
23 июня |
|
Температура дневная, °С |
15-18 |
0-5а |
7-10 |
20-25 |
7-10 |
15-18 |
20-25 |
23-30 |
Влажность воздуха, % |
56-87 |
78-93 |
62-73 |
50-65 |
46-58 |
42-48 |
30-35 |
23-26 |
Срок вегетации, сут |
7-е |
27-е |
31-е |
41-е |
50-е |
56-е |
62-е |
67-е |
Интервал наблюдений, сут |
7 |
20 |
4 |
10 |
9 |
6 |
6 |
5 |
Прирост побегов, см/сут |
2,1 |
0,5 |
1,7 |
5,1 |
1,5 |
0,5 |
0,4 |
0,1 |
П р и м е ч а н и е. а —выпадение снега и заморозки на уровне почвы (-2…-6 °С). |
||||||||
Наибольший прирост (5,1 см/сут в течение 10-суточного интервала) зафиксировали при температуре 20-25 °С и влажности 50-65 %. В экстремальных условиях (выпадение снега и многократные заморозки на почве до -2…-6 °С) рост полностью не прекращался (прирост — около 0,5 см/сут за счет использования растениями краткосрочного подъема дневной температуры до 3-5 °С). Во II-III декадах июня, когда температура воздуха находится в пределах оптимума (20-25 °С), также происходило замедление роста: если в первом случае он был ограничен минимальной температурой, то во втором — снижением влажности воздуха с 78-93 % до 30-35 % и, как следствие, истощением влаги в корнеобитаемом слое почвы.
Для аппроксимированных кривых зависимости прироста от влажности получено значение R2 = 0,99, между влажностью и температурой — R2 = 1,00. Однако положительная корреляция прироста с этими показателями была достоверна только для условий, когда температура не превышает 25 °С, а влажность одновременно не опускается ниже 40 % (r = 0,93-0,73; r = 0,95).
Таким образом, в течение вегетации с уменьшением запасов влаги в почве и снижением относительной влажности воздуха до минимальных значений средняя скорость роста розеточных побегов у R. carthamoides снижалась более чем в 10 раз (с 5,1 до 0,4-0,5 см/сут). Аналогичные данные получены и для условий Белоруссии — на 4-м году жизни линейный прирост вегетативных побегов во II декаде мая был равен 4,4 см/сут, в начале июня — 0,8 см/сут (19).
В л и я н и е с т р е с с о в ы х ф а к т о р о в н а н а к о п л е-
н и е Ф Э С. В целом для лекарственных растений справедливо, что стрессовые ситуации приводят к неоднозначным изменениям содержания биологически активных веществ в фитомассе (20). В фазу отрастания снижение дневных температур воздуха с 12-15 до 5 °С, чередующихся с ночными и утренними заморозками до -6 °С, привело к торможению роста R. carthamoides, и сухая масса листа в возрасте 9-12 сут практически не изменилась (соответственно 0,12 и 0,14 мг) (табл. 2), в то время как валовое содержание ФЭС увеличилось в 1,33 раза (с 0,36 до 0,48 мг).
После прекращения заморозков как фактора, оказывающего рост- ингибирующий эффект, пул ФЭС возрос в 2,06 раза (с 48 до 99 мг), а масса листа — в 2,28 раза. Повторное резкое понижение температуры на 30-37-е сут вегетации (с кратковременными ночными заморозками до -7…-10 °С) вновь привело к существенному изменению пула ФЭС. Содержание ФЭС в этом случае возросло с 0,31 до 0,43 %, а валовый сбор — в 1,61 раза (сухая масса листа увеличилась только в 1,16 раза).
| 2. Динамика показателей накопления фитоэкдистероидов (ФЭС) во взрослых розеточных листьях Rhaponticumcarthamoidesв связи с условиями внешней среды (Архангельская обл., 7-й год жизни, 1996 год) | |||||||||
Показатель |
Срок вегетации по фазам развития растений, сут |
||||||||
отрастание |
бутонизация |
цветение |
плодоношение |
||||||
2-е |
9-е |
12-е |
22-е |
30-е |
37-е |
41-е |
57-е |
77-е |
|
Воздушно-сухая масса листа, г |
0,02 |
0,12 |
0,14 |
0,32 |
0,37 |
0,49 |
0,60 |
1,43 |
2,11 |
Содержание ФЭС, % |
0,25 |
0,30 |
0,34 |
0,31 |
0,43 |
0,33 |
0,28 |
0,11 |
0,07 |
Валовое содержание ФЭС, мг/лист |
0,05 |
0,36 |
0,48 |
0,99 |
1,59 |
1,62 |
1,68 |
1,57 |
1,48 |
Коэффициент прироста: |
|
|
|
|
|
|
|
|
|
сухой массы листа |
– |
6,00 |
1,17 |
2,28 |
1,16 |
1,32 |
1,22 |
2,38 |
1,47 |
пула ФЭС |
– |
7,20 |
1,33 |
2,06 |
1,61 |
1,02 |
1,04 |
0,93 |
0,94 |
Температура воздуха, °С |
8-10 |
12-15 |
0-4а |
3-10 |
0-5б |
15-25 |
20-30 |
25-35в |
20-25 |
Влажность воздуха, % |
65-90 |
56-87 |
78-93 |
56-78 |
62-73 |
46-58 |
38-48 |
27-32 |
22-28 |
Влажность почвы, % |
23,4 |
17,3 |
16,4 |
14,7 |
12,8 |
12,7 |
9,4 |
2,5 |
6,3 |
П р и м е ч а н и е.а — многократные заморозки до -6…-8 °С; б — ночные заморозки до -7…-10 °С; в — засуха с температурой до 48-52 °С на поверхности оголенной почвы. Прочерки означают, что процесс не наблюдали. |
|||||||||
После повышения дневной температуры воздуха (на 37-е и 41-е сут вегетации) накопление ФЭС практически прекратилось (см. табл. 2). В период с 57-х по 77-е сут, когда наблюдалась засуха (повышение температуры до 35 °С и снижение относительной влажности воздуха до 23-27 % сопровождалось водным стрессом) (см. табл. 2), молодые розеточные листья теряли тургор, апикальные части генеративных побегов желтели и засыхали. Влагообеспеченность почвы в этот период снизилась до 2,5-3,0 % от ее сухой массы, что близко к показателям наименьшей влагоемкости с разрывом капилляров пахотного слоя на супеси (21).
В отличие от низкой температуры дефицит влаги не приводил к повышению количества экдистероидов в листьях: содержание ФЭС резко падало до 0,11 % (57-е сут вегетации), а затем до 0,07 % (77-е сут). Накопление экдистероидов становилось отрицательным (коэффициент — 0,93-0,94), валовое содержание в листьях убывало. Как оказалось, существует достоверная (r = 0,99) отрицательная корреляция между накоплением ФЭС и влажностью почвы на 57-77-е сут (r=-0,88…-0,92).
Аналогичные свидетельства разнонаправленного действия температурного и водного стресса получены с алкалоидсинтезирующими растениями другими авторами (20). В полевых опытах в листьях маклеи сердцевидной (Macleayacordata) суммарное содержание действующих веществ (сангвинарин и хелеритрин) после многократных осенних заморозков в ноябре не уменьшалось по сравнению с показателями при уборке в сентябре, тогда как кратковременное снижение влажности почвы в 2,5 раза (до 40 % от полной влагоемкости) за 7 сут до уборки у мачка желтого (Glaucium flavum) приводило к уменьшению сбора алкалоида глауцина на 16 %.
По данным С.С. Шаина, у кассии остролистной (Cassia acutifolia) длительное воздействие водного стресса (засуха в течение 20 сут на экспериментальной базе Казахстанской опытной станции ВИЛАР, Чимкентская обл.) уменьшало содержание производных антрацена на 55 % (20).
Итак, для формирования продуктивности и биосинтеза фитоэкдистероидов (ФЭС) теплообеспеченность района возделывания и плодородие почвы не имеют решающего значения (в условиях прохладного климата европейского Севера на обедненных подзолистых почвах по фитомассе растения не уступают произрастающим в более южных регионах на черноземах). В ранневесенний период на фоне оптимальной влажности и низкой температуры скорость роста побегов выше, чем летом, и ростовые процессы в суточном цикле продолжаются даже в ночное время. Наибольший прирост наблюдается в вечернее время (с 1700 до 2200-2300). Экстремальные значения солнечной радиации (150-160 тыс. лк) и температуры (25-30 °С и выше) ингибируют рост побегов. Оптимальным сочетанием температуры, относительной влажности воздуха, освещенности и влажности почвы логично считать соответственно 12-25 °С, 40-80 %, 7-120 тыс. лк и 9-16 %. Скорость роста у растений 9-го года жизни в 1,5-1,7 раза выше, чем у 3-летних. Молодые растения сильнее нуждаются в свете, взрослогенеративные — зависят от оптимальной влажности почвы, что указывает на необходимость исследования роли корневой системы в биосинтезе ФЭС для реализации потенциала вида. Однофакторный эффект низких и отрицательных температур проявляется в торможении роста побегов, повышении содержания и общего количества экдистероидов в листьях (в 1,3-2,1 раза). Многофакторное действие водного стресса (который сопровождается истощением почвенной влаги на фоне высокой температуры и низкой влажности воздуха) приводит к противоположному результату — прекращению биосинтеза и оттоку ФЭС из надземных органов.
Автор выражает признательность старшему научному сотруднику биохимической лаборатории Ботанического сада Института биологии Коми НЦ УрО РАН, к.х.н. В.В. Пунегову за помощь в проведении ВЭЖХ-анализа ФЭС.
Л и т е р а т у р а
1. Т и м о ф е е в Н.П. Достижения и проблемы в изучении биологии лекарственных растений Rhaponticumcarthamoides (Willd.) Iljin и Serratulacoronata L. С.-х. биол., 2007, 3: 3-17.
2. М а с л о в Л.Н., Г у з а р о в а Н.В. Кардиотропные и антиаритмические свойства препаратов Leuzea carthamoides, Aralia mandshurica, Eleutherococcus senticosus. Эксп. и клин. фармакол., 2007, 70(6): 48-54.
3. G a u b e F., W o l f l S., P u s c h L., W e r n e r U., K r o l l T.C., S c h r e n k D., H a r t m a n n R.W., H a m b u r g e r M. Effects of Leuzea carthamoides on human breast adenocarcinoma MCF-7 cells determined by gene expression profiling and functional assays. Planta Med., 2008, 74(14): 1701-1708.
4. B a t h o r i M., P o n g r a c z Z. Phytoecdysteroids — from isolation to their effects on humans. Cur. Med. Chem., 2005, 12: 153-172.
6. D i n a n L., L a f o n t R. Effects and applications of arthropod steroid hormones (ecdysteroids) in mammals. J. Endocrinol., 2006, 191: 1-8.
7. Т и м о ф е е в Н.П. Достижения и проблемы в области изучения, использования и прогнозирования биологической активности экдистероидов. Бутлеровские сообщения, 2006, 8(2): 7-34.
8. G o r e l i c k - F e l d m a n J., M a c l e a n D., I l i c N., P o u l e v A., L i l a M.A., C h e n g D., R a s k i n I. Phytoecdysteroids increase protein synthesis in skeletal muscle cells. J. Agric. Food Chem., 2008, 56(10): 3532-3537.
9. Т и м о ф е е в Н.П., В о л о д и н В.В., Ф р о л о в Ю.М. Распределение 20-гидроксиэкдизона в структуре биомассы надземной части Rhaponticum carthamoides (Willd.) Iljin. Растительные ресурсы, 1998, 34(3): 63-69.
10. B a k r i m A., M a r i a A., S a y a h F., L a f o n t R., T a k v o r i a n N. Ecdysteroids in spinach (Spinacia oleracea L.): biosynthesis, transport and regulation of levels. PlantPhysiol. Biochem., 2008, 46(10): 844-854.
11. Т и м о ф е е в Н.П. Накопление и изменчивость содержания действующих веществ (экдистероидов) в лекарственном сырье левзеи сафлоровидной. С.-х. биол., 2009, 1: 106-117.
12. К а р н а ч у к Р.А., П р о т а с о в а Н.Н., Д о б р о в о л ь с к и й М.В., Р е в и н а Т.А.,
Н и ч и п о р о в и ч А.А. Физиологическая адаптация листа левзеи к спектральному составу света. Физиол. раст., 1987, 34(1): 51-59.
13. Т и м о ф е е в Н.П. Продуктивность и динамика содержания фитоэкдистероидов в агропопуляциях Rhaponticum carthamoides и Serratulacoronata (Asteraceae) на европейском Севере. Растительные ресурсы, 2006, 42(2): 17-36.
14. Природно-климатический очерк Котласского района Архангельской области. М., 1994.
15. П у н е г о в В.В., С а в и н о в с к а я Н.С. Метод внутреннего стандарта для определения экдистероидов в растительном сырье и лекарственных формах с помощью ВЭЖХ. Растительные ресурсы, 2001, 37(1): 97-102.
Справочник агронома Нечерноземной зоны /Под ред. Г.В. Гуляева. М., 1990.
16. Т и м о ф е е в Н.П., K o w a l s k i P., K r y w u c h J. Сравнительные результаты 4-летнего культивирования Rhaponticumcarthamoides(Willd.) Iljin в условиях Польши и европейского Севера России. В сб.: Лекарственные растения и биологически активные вещества: фитотерапия, фармация, фармакология. Белгород, 2008: 264-267.
17. Ш е в е л у х а В.С. Рост растений и его регуляция в онтогенезе. М., 1992.
18. Г о л о в к о Т.К., Г а р м а ш Е.В., К у р е н к о в а С.В., Т а б а л е н к о в а Г.Н.,
Ф р о л о в Ю.М. Рапонтик сафлоровидный в культуре на европейском Севере-Востоке (эколого-физиологические исследования). Сыктывкар, 1996.
19. Б о р е й ш а М.С., С е м е н о в Б.Я., Ч е к а л и н с к а я И.И. Маралий корень (рапонтикум сафлоровидный). Минск, 1985.
20. Ш а и н С.С. Биорегуляция продуктивности растений. М., 2005.
21. К у л а к о в с к а я Т.Н. Оптимизация агрохимической системы почвенного питания растений. М., 1990.
N.P. Timofeev
The growth processes in Rhaponticum carthamoides (Willd.) Iljin in the conditions of Arkhangel’skaya oblast’ and their link with a biosynthesis ecdysteroids are investigated. The environmental factors limiting growth of propagules (a range of growth responses to light exposure, temperature and humidity), resistance to the low and negative temperatures, and influence of stressful factors on synthesis and accumulation of ecdysteroids in leaves organs are established.
Key words: Rhaponticum carthamoides, Leuzea, environmental factors, growth of propagules, stress, biosynthesis ecdysteroids.
Научно-производственное предприятие «КХ БИО», |
Поступила в редакцию |




