








УДК 633.12:575.174.015.3:577.2.08
АНАЛИЗ ГЕНОМНОГО РАЗНООБРАЗИЯ ОБРАЗЦОВ И СОРТОВ ГРЕЧИХИ ПОСЕВНОЙ И ТАТАРСКОЙ ISSR-МЕТОДОМ
Г.Д. КАДЫРОВА1, Ф.З. КАДЫРОВА1, Е.В. МАРТИРОСЯН2, Н.Н. РЫЖОВА2
Для оценки генетического разнообразия представителей культивируемых видов Fagopyrum (27 образцов разного эколого-географического происхождения, включая 14 отечественных сортов) осуществили подбор ISSR-праймеров, способных выявлять внутривидовой полиморфизм, и получили специфичные полиморфные ISSR-спектры амплификации ДНК-фрагментов. Впервые определены группы геномного сходства исследованных сортов и образцов. Полученные данные могут быть использованы в селекционных программах для молекулярной характеристики сортов и гибридов гречихи.
Ключевые слова: гречиха, ISSR-метод, межмикросателлитные последовательности, генетическое разнообразие сортов.
Простые тандемно повторяющиеся последовательности ДНК — мини- и микросателлиты представляют собой основной компонент всех эукариотических геномов. В целом число микросателлитных повторов колеблется от 5x103 до 3x105 на геном, по которому они распределены достаточно равномерно (1). Как правило, многие мини- и микросателлиты характеризуются высокой вариабельностью даже у близкородственных организмов (2). Исследование полиморфизма микросателлитных и межмикросателлитных последовательностей методом ISSR (intersimple sequence repeats) (3) основано на амплификации геномной ДНК с использованием 15-20- нуклеотидных праймеров, гомологичных микросателлитному повтору, часто с короткой (2-4 нуклеотида) случайной последовательностью, называемой якорной (anchor) на 3´- или 5´-конце, что повышает селективную способность праймера и воспроизводимость ISSR-спектров, полученных при амплификации участков ДНК между двумя близко расположенными и противоположно ориентированными микросателлитами. ISSR- и RAPD-спектры (random amplified polymorphic DNA) аналогичны, но обычно первые содержат больший набор полиморфных полос (до 97), что позволяет выявлять более высокий уровень геномного полиморфизма (4, 5).
В настоящее время метод ISSR широко используется при обнаружении внутривидового полиморфизма, в первую очередь у близкородственных генотипов культивируемых растений. Это особенно актуально в связи с программами современной селекции сортов принципиально нового качественного уровня (4, 6-8), для которой требуется создание обширного генофонда исходного материала, сочетающего множество хозяйственно полезных признаков, что невозможно без расширения генетического пула и предварительной оценки природного разнообразия вида.
Вид Fagopyrum esculentum Moench (гречиха посевная)семейства Polygonaceae (сем. Гречишные)относится к ценнейшим крупяным культурам и приобретает все большее значение в качестве важного диетического продукта, некоторые формы гречихи посевной используются как сырье для получения рутина — ценного лекарственного препарата из группы флавоноидов. С целью создания обширного генетического пула гречихи посевной в селекцию отечественных сортов весьма широко вовлекались местные популяции F. esculentum ssp. esculentum(9, 10). Уже к концу 1950-х годов на их основе было районировано более 50 местных сортов (11), которые, в свою очередь, продолжают включать в селекционную работу. Еще один культивируемый вид гречихи — F. tataricum ssp. tataricum, его возделывают преимущественно в высокогорных районах Китая, Японии, Индии, Непала, а также в ряде европейских стран.В селекции сортов гречихи посевной он служит компонентом скрещиваний, применяемым для придания растениям большей морозостойкости и повышения общей жизнеспособности (12).
Широкое использование местных природных популяцийF. esculentum и образцов культурного вида F. tataricum в значительной мере способствовало увеличению урожайности отечественных сортов гречихи за счет усиления ростовых процессов и устойчивости к условиям окружающей среды. Однако возникает вопрос, насколько генетический пул современных сортов гречихи поддерживает естественное разнообразие вида F. esculentum и насколько полно используется потенциал природных популяций в современной селекции. Немногочисленные исследования внутривидового разнообразия местных дикорастущих форм и сортовF. tataricum и F. esculentum (преимущественно японской селекции) с применением молекулярных методов (AFLP — amplified fragment length polymorphism, RAPD — random amplified polymorphic DNA, SSR — simple sequence repeats) были предприняты японскими учеными (13-15). В отношении сортов российской селекции имеются лишь данные, полученные нами ранее с помощью RAPD-анализа (16).
Целью этой работы стал ISSR-анализ внутривидового генетического полиморфизма у Fagopyrum esculentum, а также оценка генетического пула отечественных сортов и образцов гречихи, в том числе F. tataricum, наиболее часто используемых в селекции.
Методика. Для молекулярного анализа геномов культивируемых видов было подобрано 27 образцов гречихи коллекций генбанка Всероссийского НИИ растениеводства (г. Санкт-Петербург) и Татарского НИИ сельского хозяйства (г. Казань): F. esculentum—14 сортов и 8 образцов, F. ta-taricum—5 образцовприродных популяций разного эколого-географиче-ского происхождения.
Растительную ДНК выделяли из 8-10-суточных проростков по стандартной методике (17) с дополнительной депротеинизацией смесью фенола с хлороформом. В работе использовали 13 олигонуклеотидных последовательностей, гомологичных микросателлитным повторам.
Полимеразную цепную реакцию (ПЦР), разделение и визуализацию продуктов ПЦР проводили по стандартным методикам (18). Реакционная смесь (объем — 15 мкл) содержала 100 нг тотальной растительной ДНК, 0,5 мкМ праймера, 0,2 мМ каждого dNTP, 2,5 мМ MgCl2, 0,3 ед. Taq-полимеразы («Диалат ЛТД», г. Москва) в соответствующем 1x буфере. Амплификацию с предварительной денатурацией (94 °С, 5 мин) проводили в термоциклере GeneAmp PCR System2700 («Applied Biosystems», США) в режиме: денатурация — 94 °С, 30 с, отжиг праймера — 30 с (температуру подбирали для каждого праймера индивидуально), элонгация — 72 °С, 1 мин (40 циклов). Температуру плавления для каждого праймера рассчитывали по формуле Т = 69,3 + 0,41(GC, %) - 650/L, где L — число нуклеотидов в праймере; GC, % — доля GC-нуклеотидов в праймере. Продукты амплификации разделяли электрофорезом в 1,7 % агарозном геле (High resolution, «Sigma», MetaPhor, «Cambrex», США) в 1½ТВЕ-буфере с последующим окрашиванием бромистым этидием.
Для статистической обработки данных по каждому из праймеров составляли бинарные матрицы (1/0) и рассчитывали коэффициенты генетических различий в программе STATISTICA 6.0 с использовании методов объединения (древовидной кластеризации) и невзвешенного попарного среднего UPGMA (http://www.statsoft.com).
Результаты. Характеристика изученных образцов приведена в таблице 1; образцы гречихи татарской отбирали, основываясь на данных RAPD-анализа (16).
| 1. Происхождение изученных образцов рода Fagopyrum | ||
Вид |
№ по каталогу ВИР, сорт |
Происхождение |
F. tataricum |
к-73 |
Монголия |
F. tataricum |
к-65 |
Татарстан |
F. tataricum |
к-38 |
Брестская обл. |
F. tataricum |
к-25 |
Тернопольская обл. |
F. tataricum |
к-57 |
Польша |
F. esculentum |
к-4131 |
Канада |
F. esculentum |
к-4233 |
Япония |
F. esculentum |
к-549 |
Япония |
F. esculentum |
к-4170 |
Индия |
F. esculentum |
к-4117 |
Китай |
F. esculentum |
к-140 |
Китай |
F. esculentum |
к-143 |
Китай |
F. esculentum |
к-4350 |
Непал |
F. esculentum |
Кама |
ТатНИИСХ (г. Казань) |
F. esculentum |
Чатыр Тау |
ТатНИИСХ (г. Казань) |
F. esculentum |
Скороспелая 86 |
ВНИИЗБК (г. Орел) |
F.esculentum |
Краснострелецкая |
ТатНИИСХ (г. Казань), ВНИИЗБК (г. Орел) |
F. esculentum |
Казанская крупнозерная |
ТатНИИСХ (г. Казань) |
F. esculentum |
Каракитянка |
ТатНИИСХ (г. Казань) |
F. esculentum |
Саулык |
ТатНИИСХ (г. Казань) |
F.esculentum |
Черемшанка |
ТатНИИСХ (г. Казань) |
F.esculentum |
Богатырь |
Шатиловская опытная станция (Орловская обл.) |
F.esculentum |
Шатиловская 5 |
ВНИИЗБК (г. Орел) |
F.esculentum |
Баллада |
ВНИИЗБК (г. Орел) |
F.esculentum |
Молва |
ВНИИЗБК (г. Орел) |
F.esculentum |
Батыр |
ТатНИИСХ (г. Казань) |
F.esculentum |
Деметра |
ВНИИЗБК (г. Орел) |
П р и м е ч а н и е. ТатНИИСХ — Татарский НИИ сельского хозяйства, ВНИИЗБК — Всероссийский НИИ зернобобовых культур, ВИР — Всероссийский НИИ растениеводства. |
||
Использованные праймеры (табл. 2) представляли собой в основном динуклеотидные повторы с 1-3 дополнительными селективными якорными нуклеотидами на 3´-конце, а также три-, тетра- и пентануклеотидные повторы. Праймеры отбирали, исходя из данных об их способности детектировать геномный полиморфизм (6, 19, 20).
| 2. Характеристика использованных ISSR-праймеров и результат амплификации | ||
ISSR-праймер |
Последовательность |
Результат амплификации (число детектированных ISSR-фрагментов ДНК) |
M1 |
(AC)8CG |
Дифференцированный спектр (35 полиморфных фрагментов) |
M2 |
(AC)8(С/T)G |
Дифференцированный спектр (29 полиморфных фрагментов) |
М3 |
(GA)8(C/T)C |
Отсутствие ПЦР-продукта |
М4 |
(AG)8(C/T)C |
Малополиморфный спектр |
М5 |
(AT)8 |
Шмер |
М6 |
(CAC)5 |
Малополиморфный спектр |
M7 |
(CAG)5 |
Дифференцированный спектр (25 полиморфных фрагментов) |
M8 |
(GTG)5 |
Малополиморфный спектр |
M9 |
(GACAC)4 |
Отсутствие ПЦР-продукта |
M10 |
(CA)6(A/G)G |
Отсутствие ПЦР-продукта |
M11 |
(CA)6(A/G) |
Дифференцированный спектр (24 полиморфных фрагмента) |
M12 |
(CA)6(A/G)(C/T) |
Малополиморфный спектр |
M13 |
(AGC)4(C/T) |
Мономорфный спектр |
Тестирование ISSR-праймеров на ограниченном наборе образцов (по три для каждого вида гречихи) показало, что не все из них обеспечивали амплификацию четкого мультилокусного полиморфного спектра фрагментов генома. Использование праймеров М4, М12 и М13 (см. табл. 2) выявляло межвидовой полиморфизм, однако спектры у образцов одного вида были либо малополиморфными (М4, М12), либо мономорфными (М13). С четырьмя праймерами получили отрицательный результат: либо амплификация отсутствовала (М3, М9, М10), либо отмечали образование шмера (М5). Детекция продуктов амплификации в виде шмера, как это наблюдалось при использовании праймера М5, встречается довольно часто и, по всей видимости, связана либо со структурой самой последовательности праймера, либо с частотой встречаемости соответствующего микросателлита в геноме. Отсутствие продуктов амплификации при использовании праймеров М3, М9 и М10 могло быть обусловлено тем, что в геноме гречихи микросателлитные повторы (GA)8(C/T)C, (CA)6(A/G)G и (GACAC)n редки. В ряде работ высказано также предположение о частичной самокомплементарности таких праймеров и образовании двуцепочечных структур праймер—праймер как возможной причине отсутствия амплификации (8, 21).
Для последующего использования отобрали три наиболее информативных праймера (М1, М2 и М7), приводящих к амплификации воспроизводимых, четких, полиморфных спектров, состоящих не менее чем из 20 фрагментов. Всего для 27 генотипов гречихи получили 89 полиморфных ISSR-фрагментов, длина которых варьировала от 2700 п.н. до 320 п.н. Следует отметить, что все детектированные фрагменты генома гречихи были полиморфными, то есть степень полиморфизма, выявляемая с помощью трех отобранных ISSR-праймеров, составила 100 %, в то время как при RAPD-анализе — 88 % (16), что согласуется с данными о более высокой дискриминационной способности первого метода по сравнению со вторым (4).
Всего было маркировано пять ISSR-фрагментов, специфичных для всех исследованных представителей вида F. esculentum, и три — характерных для представителей вида F. tataricum, а также специфичные ISSR-фрагменты у ряда образцов и сортов. В частности, у образца F. esculentum из Китая (к-143) присутствовали три уникальных фрагмента размером 780 п.н., 990 п.н. (праймер М1) и 1500 п.н. (праймер М7), идентифицировались фрагменты, специфичные для сортов Богатырь (1000М2, 1600М2), Скороспелая 86 (780М7, 970М7), Деметра (850М1, 1700М1), Каракитянка (1000М1, 650М2) и Батыр (910М2). Эти фрагменты могут быть использованы при разработке геном-специфичных ДНК-маркеров для сортов, образцов и видов гречихи.
| 3. Коэффициенты внутри- и межвидовых генетических различий у изученных образцов гречихи Fagopyrum | |||||
Анализируемая группа |
Среднее значение |
Стандартная ошибка |
Стандартное отклонение |
Min |
Max |
F. esculentum (сорта) |
0,169 |
0,005 |
0,051 |
0,056 |
0,303 |
F.esculentum (образцы) |
0,191 |
0,009 |
0,048 |
0,101 |
0,281 |
F.esculentum |
0,182 |
0,003 |
0,051 |
0,045 |
0,315 |
F. esculentum (сорта)–F. esculentum |
0,190 |
0,005 |
0,050 |
0,045 |
0,315 |
F.tataricum |
0,099 |
0,014 |
0,044 |
0,011 |
0,169 |
F. esculentum (образцы)–F. tataricum |
0,304 |
0,012 |
0,073 |
0,112 |
0,438 |
F. esculentum (сорта)–F. tataricum |
0,292 |
0,008 |
0,063 |
0,135 |
0,427 |
F.esculentum–F.tataricum |
0,300 |
0,010 |
0,070 |
0,110 |
0,440 |
П р и м е ч а н и е. Коэффициенты генетических различий рассчитывали в программе STATISTICA 6.0 с использованием методов объединения (древовидной кластеризации) и невзвешенного попарного среднего UPGMA (http://www.statsoft.com). |
|||||
Рассчитанные на основе ISSR-спектров и данных о полиморфизме ДНК-фрагментов коэффициенты попарных генетических различий у представителей рода Fagopyrum варьировали от 0,002 до 0,440. Пределы, характеризующие межвидовое разнообразие гречихи посевной и гречихи татарской, оказались значительно уже; наименьший разброс (0,135-0,427) отмечали в паре F. esculentum (сорта)—F. tataricum(табл. 3).
Внутривидовая геномная вариабельность у F. esculentum (0,045-0,315), как и при проведенном ранее RAPD-анализе (16), почти в 2 раза превышала показатель у F. tataricum (0,010-0,169) (см. табл. 3).
Интересно отметить, что межсортовые различия F. esculentum были весьма высоки (коэффициенты попарного сходства — 0,056-0,303) и соответствовали геномной вариабельности у дикорастущих образцов F. esculentum (0,101-0,281) из Японии, Китая, Непала, что свидетельствует о широкой генетической основе отечественных сортов гречихи посевной. Сходный вывод был сделан ранее по результатам RAPD-анализа (16).
|
Дендрограмма, отражающая степень генетического сходства исследованных видов, сортов и образцов гречихи Fagopyrum (построена с использованием метода иерархического кластерного анализа UPGMA по результатам ISSR-маркирования). |
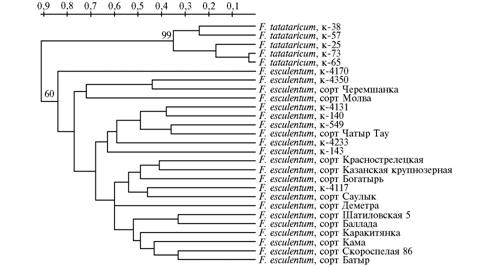
Дендрограмма, построенная по данным кластерного анализа, выявила четкую видовую дифференциацию F. tataricum и F. esculentum, разделив исследованные образцы на две основные группы с достоверными значениями бутстрепа (рис.). Представители F. tataricum формировали сравнительно малополиморфную группу. В свою очередь, в кластере, объединяющем представителей F. esculentum,выделялись три основных группы образцов с различной степенью сходства. В первую (базальную и наиболее полиморфную) кладу объединились дикорастущие местные популяции F. esculentumиз Непала и Индии, а также два сорта — Черемшанка и Молва. Во вторую кладу вошли образцы из Китая и Японии, а также сорт Чатыр Тау. Третья клада была полностью представлена изученными сортами. Несмотря на то, что межсортовые группировки этой клады поддерживались невысокими значениями бутстрепа, наблюдалась некоторая корреляция между степенью сходства и морфобиологическими особенностями сортов этих групп. Так, в группу Каракитянка—Кама—Скороспелая 86—Батыр вошли скороспелые и засухоустойчивые сорта с ограниченным ветвлением растений. В этой группе сорт Батыр выделялся большей продолжительностью вегетации, но известно, что в его родословной присутствует ультраскороспелая форма — сорт Казанская 309. Сорта группы Шатиловская 5—Баллада были близки по происхождению и хозяйственно-биологи-ческим свойствам. В родословной сорта Казанская крупнозерная проявилось присутствие материала отборов из сорта Краснострелецкая. Сорт Деметра формировал отдельную ветвь, что может быть связано с его значительными морфобиологическими особенностями.
Таким образом, для анализа генома гречихи Fagopyrum нами впервые применен ISSR-метод. Определены ISSR-праймеры, способные выявлять внутривидовой полиморфизм. Для каждого образца F. esculentum и F. tataricum получены уникальные ISSR-спектры и выявлены образец- и сортоспецифичные фрагменты, которые в дальнейшем могут быть использованы для разработки геном-специфичных ДНК-маркеров гречихи. На основе вариабельности микросателлитных и межмикросателлитных последовательностей определена степень генетических различий у исследованных образцов и показано, что межсортовой полиморфизм отечественных сортов F. esculentum поддерживается на высоком уровне и сравним с генетическим разнообразием дикорастущих форм. Полученные результаты могут быть использованы в селекционных программах при идентификации сортов и гибридов.
Л и т е р а т у р а
1. K a t t i M.V., R a n j e k a r P.K., G u p t a V.S. Differential distribution of simple sequence repeats in eukaryotic genome sequences. Mol. Biol. Evol., 2001, 18(7): 1161-1167. 2. T o t h G., G a s p a r i Z., J u r k a J. Microsatellites in different eukaryotic genomes: survey and analysis. Genome Res., 2000, 10: 967-981. 3. Z i e t k i e w i c z E., R a f a l s k i A., L a b u d a D. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics, 1994, 20: 176-183. 4. S i c a M., G a m b a G., M o n t i e r i S., G a u d i o L., A c e t o S. SSR markers show differentiation among Italian populations Asparagus acutifolius L. BMC Genetics, 2005, 6. 5. M u t h u s a m y S., K a n a g a r a j a n S., P o n n u s a m y S. Efficiency of RAPD and ISSR markers system in accessing genetic variation of rice bean (Vigna umbellata) landraces. Electronic J. Biotechnol., 2008, 11(3): 1-10. 6. P r e v o s t A., W i l k i n s o n M.J. A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet., 1999, 98: 107-112. 7. G i l b e r t J.E., L e w i s R.V., W i l k i n n s o n M.J., C a l i g a r i P.D.S. Developing an appropriate strategy to assess genetic variability in plant germplasm collections. Theor. Appl. Genet., 1999, 98: 1125-1131. 8. J o s h i S.P., G u p t a V.S., A g g a r w a l R.K., R a n j e k a r P.K., B r a r D.S. Genetic diversity and phylogenetic relationship as revealed by inter simple sequence repeat (ISSR) polymorphism in the genus Oryza.Theor. Appl. Genet., 2000, 100: 1311-1320. 9. Г о р и н а Е.Д. Исходный материал, его биологические особенности и эффективность методов селекции гречихи. Автореф. докт. дис. Жодино, 1981. 10. Ф е с е н к о Н.В. Биологические основы и методы селекции гречихи. Автореф. докт. дис. Л., 1974. 11. К р о т о в А.С. Гречиха СССР и улучшение ее хозяйственных признаков. Автореф. докт. дис. Л., 1955. 12. C a m p b e l l C.G. Buckwheat. Fagopyrum esculentum Moench. Rome, Italy, 1997. 13. T s u j i K., O h n i s h i O. Phylogenetic relationships among wild and cultivated Tartary buckwheat (Fagopyrum tataricum Gaert.) populations revealed by AFLP analyses. Genes Genet. Syst., 2001, 76: 47-52. 14. I w a t a H., I m o n K., T s u m u r a Y., O h s a w a R. Genetic diversity among Japanese indigenous common buckwheat (Fagopyrum esculentum) cultivars as determined from amplified fragment length polymorphism and simple sequence repeat markers and quantitative agronomic traits. Genome, 2005, 48: 367-377. 15. K o n i s h i T., Y a s u i Y., O h n i s h i O. Original birthplace of cultivated common buckwheat inferred from genetic relationships among cultivated populations and neutral populations of wild common buckwheat revealed by AFLP analysis. Genes Genet. Syst., 2005, 80: 113-119. 16. К а д ы р о в а Г.Д., К а д ы р о в а Ф.З., Р ы ж о в а Н.Н., К о ч и е в а Е.З. RAPD-анализ геномного полиморфизма видов и сортов рода Fagopyrum.Экологическая генетика, 2008, VI(3): 3-10.17. Р ы ж о в а Н.Н., П ы ш н а я О.В., К о ч и е в а Е.З. Молекулярный RAPD анализ гибридов перца. С.-х. биол., 2000, 5: 104-106.
18. W i l l i a m s J.G.K., K u b e l i k A.R., L i v a k K.J., R a f a l s k i J.A., T i n g e y S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucl. Acids Res., 1990, 18: 6531-6535. 19. F a n g D.Q., R o o s e M.L. Identification of closely related citrus cultivars with inter-simple sequence repeats markers. Theor. Appl. Genet., 1997, 95: 408-417. 20. W a n g G., M a h a l i n g a m R., K n a p H.T. (C-A) and (G-A) anchored simple sequence repeats (ASSRs) generated polymorphism in soybean, Glycine max (L.) Merr. Theor. Appl. Genet., 1998, 96: 1086-1096. 21. C e k i c C., B a t t e y N.H., W i l k i n s o n M.J. The potential of ISSR-PCR primer pair combinations for genetic linkage analysis using the seasonal flowering locus in Fragaria as a model. Theor. Appl. Genet., 2001, 103: 540-546.
G.D. Kadyrova1, F.Z. Kadyrova1, E.V. Martirosyan2, N.N. Ryzhova2
For an assessment of a genetic polymorphism of common buckwheat (F. esculentum, 27 samples of different ecologo-geographic origin, including 14 domestic cultivars) the authors determined the ISSR-primers of revealing intraspecific polymorphism and obtained the genomspecific polymorphic ISSR-spectra of DNA fragments. Genomic variability and genomic linkage groups cultivars of common buckwheat were determined for the first time. Obtained data can be used in buckwheat breeding program and further molecular characterization of investigated cultivars.
Key words: buckwheat, ISSR-method, inter-microsatellite sequences, genetic variability of cultivars.
1ГНУ Татарский НИИ сельского хозяйства Россельхозакадемии,
420059 г. Казань, Оренбургский тракт, 48, |
Поступила в редакцию |




