








УДК 634.11:581.5:577.125.33
ОСОБЕННОСТИ ПЕРЕКИСНОГО ОКИСЛЕНИЯ ЛИПИДОВ МЕМБРАН В ЛИСТЬЯХ ЯБЛОНИ В УСЛОВИЯХ ТЕХНОГЕННОГО ЗАГРЯЗНЕНИЯ
М.Н. КУЗНЕЦОВ, П.С. ПРУДНИКОВ
Изучали влияние техногенного загрязнения на интенсивность перекисного окисления липидов мембран (ПОЛ) в листьях яблони сорта Осеннее полосатое. Показано, что баланс фитогормонов и активность пероксидазы изменялись в зависимости от степени техногенного загрязнения, что способствовало интенсификации ПОЛ в тканях.
Ключевые слова: пероксидаза, индолилуксусная кислота, абсцизовая кислота, ПОЛ.
Под действием неблагоприятных факторов среды в кислородпотребляющих биологических системах, как правило, усиливается образование свободных радикалов, что приводит к интенсификации процессов перекисного окисления липидов (ПОЛ) мембран и нарушению целостности структурно-функционального состояния клеток (1-2). Об интенсивности протекания ПОЛ большинство авторов судят по накоплению в тканях конечного продукта липопероксидации — малонового диальдегида (МДА). Так, в листьях берез, произрастающих в неблагополучной по техногенному загрязнению зареченской части Нижнего Новгорода, отмечено повышенное содержание МДА (3). Аналогичные результаты получены на мхах (FunariahygrometricaHedw.) при действии ионов свинца на гаметофит (4). Обнаружена прямая зависимость между количеством МДА на всех стадиях развития растений и концентрацией тяжелого металла в субстрате.
Фитогормоны служат одними из основных регуляторов роста и развития растений (5). Согласно данным литературы, под действием стрессоров видоизменяется баланс гормонов, что приводит к нарушению физиологических процессов. В условиях гипо- и гипертермии в растениях, как правило, увеличивается содержание абсцизовой кислоты (АБК) (6-7) и снижается количество цитокининов и индолилуксусной кислоты (ИУК) (6, 8). В исследованиях, проведенных на огурце, выявлено повышение содержания эндогенной АБК в семядольных листьях под действием ионов кадмия (9). Также известно, что фитогормоны активируют экспрессию генов ферментов антиоксидантной защиты (10), интенсифицируют окислительные процессы, поставляющие активные формы кислорода (11). В свою очередь, компоненты антиоксидантной системы могут регулировать метаболизм фитогормонов (12-13). Вместе с тем в литературе отсутствуют сведения о влиянии техногенного загрязнения на регуляцию перекисного гомеостаза гормональной системой растений.
Цель настоящей работы заключалась в изучении влияния техногенного загрязнения на перекисное окисление липидов мембран листьев яблони в зависимости от функционирования гормональной и антиоксидантной систем растения.
Методика. Исследования проводили на Думчинском шлакоотвале (Мценский р-н, Орловская обл.) на растениях яблони сорта Осеннее полосатое, удаленных от источника техногенного загрязнения на 100 и 3000 м. В качестве контроля использовали листья растений яблони того же сорта, произрастающих на относительно благополучной по показателям техногенного загрязнения территории (Всероссийский НИИ селекции плодовых культур, Орловский р-н).
Фитогормоны экстрагировали из одной навески листьев (250 мг сухой массы), предварительно зафиксированной в парах этанола комплексным методом, разработанным в лаборатории В.И. Кефели (14). Содержание ИУК и АБК в листьях определяли методом биологической пробы. Для этого сухие остатки экстрактов растворяли в этаноле и наносили на хроматографическую бумагу. При фракционировании ИУК и АБК в качестве растворителя применяли смесь изопропанол:аммиак:вода (10:1:1). Биотестом служили отрезки колеоптилей пшеницы сорта Мироновская 808. Количество гормонов рассчитывали по калибровочной кривой, построенной для ИУК и АБК («Serva», Германия). Активность пероксидазы (усл. ед.) определяли методом Бояркина (15). В качестве субстрата использовали бензидин, который при окислении образует соединение синего цвета. О содержании МДА (мкмоль/г сырой массы) судили по цветной реакции с тиобарбитуровой кислотой (ТБК) (16).
Анализы проводили в 5-кратной биологической повторности. Достоверность результатов оценивали по стандартным методикам с использованием программ BIOSTAT и EXEL (17).
Результаты. На расстоянии 100 м от источника интоксикации содержание ИУК в листьях яблони уменьшалось в 7,6 раза, при удалении на 3000 м — в 3,3 раза по сравнению с контролем; количество АБК возрастало соответственно в 3,5 и 2,0 раза (рис., А).
Активность суммарной пероксидазы на расстоянии 100 м была на 50,7 % ниже контроля (см. рис., Б). В работах других авторов показано, что АБК и ИУК инициируют экспрессию нескольких групп генов раннего ответа растений на действие стрессоров, в том числе генов антиоксидантных ферментов (10, 18). Так, в условиях гипотермии при возрастании количества АБК и незначительном снижении — ИУК активность пероксидазы и каталазы в побегах возобновления клубней картофеля, обогащенных селеном, повышалась (19). Вероятно, в наших исследованиях резкий фитогормонональный дисбаланс приводил к ингибированию антиокислительных процессов. На фоне меньшей диспропорции между содержанием ИУК и АБК в листьях растений, удаленных от источника загрязнения на 3000 м, активность пероксидазы была ниже контроля на 19,0 %.
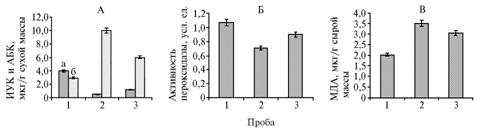 |
Содержание индолилуксусной (ИУК, а) и абсцизовой (АБК, б) кислоты (А), активность пероксидазы (Б) и содержание малонового диальдегида (МДА) (В) в листьях яблони сорта Осеннее полосатое при разном удалении от источника техногенного загрязнения (Думчинский шлакоотвал, Мценский р-н, Орловская обл.): 1 — контроль, 2 и 3 — пробы, отобранные на удалении соответственно 100 и 3000 м. |
В работах F. Zhang с соавт. показано снижение активности аскорбатпероксидазы у пшеницы при высоких дозах кадмия (20). Вместе с тем в растениях PotamogetoncrispusL. под влиянием ионов Pb2+ (20 и 15 мг/л) отмечали увеличение активности пероксидазы и инактивацию других антиоксидантных ферментов — каталазы и супероксиддисмутазы (21). Неодинаковый ответ изучаемого фермента на присутствие тяжелых металлов может быть связан с видовой и сортовой спецификой растений, разным механизмом действия токсикантов и их концентрацией в среде. Так, в растениях металлотолерантного сорта гороха Харьковский янтарный под влиянием ионов кадмия активность глутатионпероксидазы снижалась, тогда как у проростков неустойчивого сорта Резонатор — возрастала (22). Активность супероксиддисмутазы и глутатионпероксидазы в листьях Elodeacanadensisповышалась при низкой концентрации ионов никеля, а затем снижалась при более высоких дозах токсиканта (23).
Изменения в гормональном статусе и активности антиоксидантного фермента пероксидазы, вызванные интоксикационной нагрузкой, оказывали влияние на интенсивность перекисного окисления липидов мембран. На расстоянии 100 м от шлакоотвала содержание МДА в листьях растений было выше на 73 % по сравнению с контролем (см. рис., В). При удалении на 3000 м интенсивность процессов ПОЛ превышала контроль на 48 %. В исследованиях, проведенных рядом авторов, также показано увеличение активности ПОЛ мембран под действием тяжелых металлов на растительный организм (3, 4, 21).
Таким образом, баланс фитогормонов и активность антиоксидантного фермента пероксидазы в листьях яблони сорта Осеннее полосатое изменяются в зависимости от степени техногенной нагрузки, что способствует интенсификации процессов липопероксидации биомембран.
Л и т е р а т у р а
1. Б а р а б о й В.А. Механизмы стресса и перекисное окисление липидов. Усп. совр. биол., 1991, 111(6): 923-931.
2. D a t J., V a n d e n a b e e l e S., V r a n j v a E. e.a. Dual action of the active oxygen species during plant stress responses. Cell Mol. Live Sci., 2000, 57: 779-795.
3. Е р о ф е е в а Е.А., Н а у м о в а М.М., Л и с и ц и н а О.Н. Сравнительный анализ влияния средового стресса на содержание диеновых конъюгатов и малонового диальдегида в листовой пластинке березы повислой, произрастающей в условиях городской среды. Мат. межд. конф. «Современная физиология растений: от молекул до экосистем». Сыктывкар, 2007, ч. 2: 133-135.
4. К и я к Н.Я. Действие свинца на интенсивность процессов ПОЛ на разных этапах развития гаметофита мха FumariahygrometricaHedw. Мат. межд. конф. «Современная физиология растений: от молекул до экосистем». Сыктывкар, 2007, ч. 2: 187-189.
5. П о л е в о й В.В. Фитогормоны. Л., 1982.
6. В е с е л о в А.П. Гормональная и антиоксидантная системы при ответе растений на тепловой шок. Автореф. докт. дис. М., 2001.
7. Т а л а н о в а В.В., Т и т о в А.Ф. Влияние неблагоприятных факторов среды на содержание АБК и устойчивость растений. Тез. докл. Межд. науч. конф. «Проблемы физиологии растений Севера». Петрозаводск, 2004: 175.
8. Ф а р х у т д и н о в Р.Г. Температурный фактор в гормональной регуляции водного обмена растений. Автореф. докт. дис. Уфа, 2005.
9. Т а л а н о в а В.В., Т и т о в А.Ф., Б о е в а Н.П. Влияние ионов кадмия и свинца на рост и содержание пролина и АБК в проростках огурца. Физиол. раст., 1999, 46(1): 164-167.
10. X i a n g C., M i a o Z.H., L a m E. Coordinate activation of as-1-type elements and a tobacco glutathione-S-transferase gene by auxing, salicylic acid, methyljasmonate and hydrogen peroxide. Plant Mol. Biol., 1996, 32: 415-426.
11. Я к у ш к и н а Н.И. Энергетический обмен и рост растений. В сб.: Особенности гормональной регуляции процессов обмена и темпов роста растений /Под ред. Н.И. Якушкиной. М., 1983: 3-11.
12. Г у с ь к о в А.В. Метаболизм ауксинов в растениях и его регуляция. Итоги науки и техники ВИНИТИ. Серия: Физиология растений, 1991, 8: 151.
13. K i m K.Y., K w o n H.K., K w o n S.Y. e.a. Differential expression of four sweet potato peroxidase genes in response to abscisic acid and ethephon. Phytochemistry, 2000, 54: 19-22.
14. В л а с о в П.В., М а з и н В.В., Т у р е ц к а я Р.Х. и др. Комплексный метод определения природных регуляторов роста. Первичный анализ незрелых семян кукурузы на активность свободных ауксинов, гиббереллинов и цитокининов. Физиол. раст., 1979, 26(3): 648-655.
15. Е р м а к о в А.И. Методы биохимического исследования растений. 3-е изд., перераб. и доп. Л., 1987.
16. С т а л ь н а я И.Д., Г а р и ш в и л и Т.Г. Метод определения малонового диальдегида с помощью тиобарбитуровой кислоты. В кн.: Современные методы в биохимии /Под ред. В.Н. Ореховича. М., 1977: 66-68.
17. Л а к и н Г.Ф. Биометрия. М., 1980.
18. G u a n L., S c a n d a l i o s J.G. Effect of the plant growth regulation abscisic acid and high osmoticum on the developmental expression of the maize catalase genes. Physiologia Plantarum, 1998, 104: 413-422.
19. П р у д н и к о в П.С. Влияние селена на физиолого-биохимические процессы при адаптации растений картофеля к гипотермии. Автореф. канд. дис. М., 2007.
20. Z h a n g F., S h i W., J i n Z. e.a. Response of antioxidative enzymes in cloroplasts to cadmium toxicity. J. Plant Nutr., 2003, 26: 1779-1788.
21. Х у Ц.Ц., Ш и Г.С., С у Ц.С. и др. Воздействие Pb2+ на активность антиоксидантных ферментов и ультраструктуру клеток листьев Potamogetoncrispus. Физиол. раст., 2007, 54(3): 469-474.
22. С ы щ и к о в Д.В. Специфика функционирования глутатионпероксидазы различных по устойчивости к кадмию сортов гороха. Мат. межд. конф. «Современная физиология растений: от молекул до экосистем». Сыктывкар, 2007, ч. 2: 394-396.
23. М а л е е в а М.Г., Н е к р а с о в а Г.Ф. Влияние возрастающих концентраций никеля на физиологические показатели ElodeaсanadensisMichx.Мат. межд. конф. «Современная физиология растений: от молекул до экосистем». Сыктывкар, 2007, ч. 2: 257-259.
PECULIARITIES OF MEMBRANE LIPID PEROXIDATION IN APPLE LEAVES IN THE CONDITIONS OF TECHNOGENIC POLLUTION
M.N. Kuznetsov, P.S. Prudnikov
The authors investigated the influence of technogenic pollution upon the intensity of membrane lipid peroxidation in apple leaves of the Autumn striped variety. In connection with degree of technogenic load the phytohormones are imbalanced and the activity of peroxidase is changed pro-moting to intensification of membrane lipid peroxidation in tissues.
Key words: peroxidase, indoleacetic acid, abscisic acid, POL.
ГНУ Всероссийский НИИ селекции плодовых |
Поступила в редакцию
|
Оформление электронного оттиска
![]()
![]()




