doi: 10.15389/agrobiology.2021.4.682rus
УДК 636.52.58:619:616-099:591.1
Исследование выполнено при поддержке гранта Российского научного фонда для реализации научного проекта 20-76-10003 «Изучение действия Т-2 и НТ-2 токсинов на пищеварение у птиц, разработка методов диагностики и создание нового комплексного препарата для профилактики микотоксикозов».
СОДЕРЖАНИЕ Т-2 И НТ-2 ТОКСИНОВ, АКТИВНОСТЬ ФЕРМЕНТОВ В КИШЕЧНИКЕ И ГЕМАТОЛОГИЧЕСКИЙ СТАТУС ЦЫПЛЯТ-БРОЙЛЕРОВ (Gallus gallus L.) ПРИ ЭКСПЕРИМЕНТАЛЬНОМ Т-2 ТОКСИКОЗЕ
В.Г. ВЕРТИПРАХОВ, А.А. ГРОЗИНА, Н.Н. ГОГИНА, И.В. КИСЛОВА, Н.В. ОВЧИННИКОВА, М.В. КОЩЕЕВА
В настоящее время описано более 400 микотоксинов. Все они обладают канцерогенными, мутагенными, тератогенными, эмбриотоксическими, аллергенными и иммуносупрессивными свойствами, подавляют клеточный и гуморальный иммунитет. В кормах птицы часто встречаются Т-2 токсин и НТ-2 токсин, которые относятся к наиболее опасным трихотеценовым микотоксинам и вызывают гастроэнтериты, некроз кожи и слизистой оболочки ротовой полости, нарушение деятельности центральной нервной системы. Содержание Т-2 токсина в комбикормах для птиц регламентировано (ПДК = 100 мкг/кг, СанПиН 2.3.2.1078-01). Поражению органов под действием микотоксинов предшествуют функциональные нарушения, которые влияют на гематологический статус и ферментативные функции в пищеварительном канале. При исследовании этих функций наиболее точные результаты дают эксперименты in vivo на фистульных животных. В настоящей работе мы использовали этот подход в сочетании с анализом микотоксинов в лиофилизированных образцах химуса и помета методом высокоэффективной жидкостной хромато-масс-спектрометрии (ВЭЖХ-МС/МС) и показали, что при скармливании контаминированного Т-2 токсином комбикорма в кишечнике птицы обнаруживается Т-2 токсин в его метаболит — НТ-2 токсин, причем последний интенсивно всасывается в кровь и, видимо, переходит в ткани, поскольку его количество значительно снижается в помете по сравнению с дуоденумом. Целью работы было определение содержания Т-2 и НТ-2 токсинов в химусе и помете, активности пищеварительных ферментов и морфо-биохимических показателей крови цыплят-бройлеров кросса Смена 8 при экспериментальном Т-2 токсикозе. Птице в течение 2 нед скармливали корм, контаминированный Т-2 токсином в количестве от 100 мкг/кг (1ПДК) до 400 мкг/кг (4ПДК). Анализ химуса и помета показал, что в кишечнике цыплят-бройлеров Т-2 токсин трансформируется в метаболит — НТ-2 токсин. При этом последний имел тенденцию к интенсивному всасыванию в кровь. Потребление корма, содержащего Т-2 в количествах 100, 200 и 400 мкг/кг, привело к увеличению протеолитической активности дуоденального химуса на 46,3-96,6 % (p < 0,05), активности липазы — на 16,8-25,5 % (p < 0,05), амилазы — на 99,7 % (при 4ПДК по Т-2 токсину). Активность щелочной фосфатазы снижалась на 23,8-27,9 % (p < 0,05). При этом в помете протеолитическая активность увеличивалась на 76,0-169,1 % (p < 0,05), активность липазы снижалась на 23,2 %(p < 0,05) (4ПДК по Т-2 токсину), амилазы — на 55,1-57,2 % (p < 0,05). Биохимические исследования крови бройлеров показали снижение активности трипсина на 12,2-22,2 % (p < 0,05), увеличение активности щелочной фосфатазы (от 52,4 % увеличения до 5-кратного). В результате фосфатазно-протеазный индекс увеличился в среднем с 31 до 92. В крови цыплят, получавших с кормом Т-2 токсин в дозе 0,4 мг/кг (4ПДК), содержание общего белка снизилось на 13,2 % (p < 0,05), глюкозы — на 13,5 % (p < 0,05), триглицеридов — в 4 раза. Вместе с тем наблюдалось снижение числа лейкоцитов в крови на 15,8-16,6 %. В лейкоцитарной формуле наблюдалась тенденция увеличения лимфоцитов в опытных группах, получавших корм с Т-2 токсином в количестве 100 и 200 мкг/кг, в отличие от псевдоэозинофилов, что связано со специфической защитой организма при развитии микотоксикоза. Установлено, что образование метаболита НТ-2 из токсина Т-2 находится в обратной зависимости от активности трипсина в химусе кишечника и фосфатазно-протеазного индекса в крови бройлеров, что можно использовать на практике в качестве теста для диагностики Т-2 токсикоза.
Ключевые слова: Т-2 токсин, НТ-2 токсин, Т-2 токсикоз, бройлеры, химус, помет, пищеварительные ферменты, протеазы, липазы, амилазы, биохимия крови, трипсин, щелочная фосфатаза, триглицериды, морфология крови, лейкоцитарная формула.
В продуктах питания и кормах часто встречаются микотоксины — вторичные метаболиты, продуцируемые видами несовершенных грибов (формальный класс Fungi imperfecti) родов Fusarium, Aspergillus, Myrothecium, Trichoderma, Trichothecium, Penicillium и др. Микотоксины различаются по химическому строению, токсичности и механизму действия (1, 2). В настоящее время известно более 400 различных микотоксинов, наиболее распространенными и опасными для здоровья человека и животных считаются афлатоксины (афлатоксин B1), фумонизины, зеараленон, трихотецены типа B (дезоксиниваленол), трихотецены типа A (Т-2 токсин) и охратоксин А (3).
Трихотеценовые микотоксины синтезируются плесневыми грибами рода Fusarium, однакоCephalosporium, Myrothecium, Stachybotrys и Trichoderma также способны продуцировать эти соединения (4). Известно более 150 структурно родственных соединений, которые подразделены по химической структуре на 4 типа (от A до D) (5). Наиболее распространены и часто встречаются токсины группы А (НТ-2 и Т-2 токсины) и группы В (дезоксиниваленол — ДОН, 3-ацетилдезоксиниваленол, 15-ацетилдезоксиниваленол, ниваленол). Т-2 и НТ-2 токсины, хотя и представляют собой малочисленную группу, относятся к самыми токсичными представителями трихотеценовых микотоксинов (6). Указанные токсины способны привести к снижению прироста живой массы, геморрагическим поражениям слизистых оболочек, абортам, снижению молочной продуктивности и яйценоскости (7).
Мониторинговым исследованиям содержания микотоксинов в кормах во всем мире уделяется большое внимание. При этом наиболее контаминированным трихотеценовыми токсинами типа А сырьем признаны кукуруза (8), комбикорм (9) и ячмень (10). В климатических условиях Российской Федерации наиболее распространены трихотеценовые микотоксины типа А (Т-2 токсин и др.), трихотеценовые микотоксины типа В (ДОН и др.), зеараленон, охратоксин, фумонизины (11, 12).
Для промышленного птицеводства значительную угрозу представляет контаминация кормов Т-2 токсином, который негативно влияет на ряд физиологических функций птицы, что обусловлено его способностью ингибировать синтез белка, изменять экспрессию генов воспаления (13), что впоследствии приводит к некрозу и десквамации эпителиальных клеток слизистой оболочки железистого желудка (14). Поскольку Т-2 токсин имеет низкую биодоступность, большая его часть трансформируется в метаболиты, в частности в НТ-2 токсин (15).
Поступление больших количеств Т-2 и НТ-2 токсинов в организм животных приводит к снижению продуктивности, изменению биохимических показателей крови (16), структуры гепатоцитов (17), спленоцитов (18), сумки Фабрициуса (19). Существенный вред животноводству наносит аддитивная токсичность Т-2 токсина при наличии в кормах охратоксина А (20), фумонизина B и дезоксиниваленола (21), циклопиазоновой кислоты (22), афлатоксина (23).
Полнорационные комбикорма составляют основу кормления сельскохозяйственных животных. Согласно данным мониторинговых исследований кормов (24), Т-2 токсин распространен во всех регионах Российской Федерации. Наиболее часто (79,1 % от исследованных проб) он встречается в комбикормах для птицы и свиней (25). Т-2 токсин также обнаружен в 32 % проб пшеницы, 70,9 % — ячменя, 94 % — кукурузы (26), что следует учитывать при составлении полнорационных комбикормов. Иммуноферментный анализ контаминации микотоксинами комбикормов для птицы (27) показал, что в них присутствуют все основные группы микотоксинов, при этом Т-2 токсин был обнаружен в 88,2 % исследованных проб.
Длительное время угрозу развития микотоксикозов оценивали на основании контроля безопасности кормов. Прямую и более информативную характеристику состояния животного при отравлении микотоксинами дает исследование биоматериалов — крови, мочи или кала (28). Метод высоко-эффективной жидкостной хромато-масс-спектрометрии (ВЭЖХ-МС/МС), который с высокой точностью позволяет определить микотоксины и их метаболиты в образцах крови, сердца, печени, селезенки, легких, почки, железистого желудка, мышечного желудка, тонкой кишки, мышц, костей, головного мозга (29).
У птицы вопросы патогенеза при потреблении контаминированных Т-2 и НТ-2 токсинами кормов остаются малоизученными, до настоящего времени не создано эффективных методов диагностики и средств профилактики этих микотоксикозов. Поражению органов под действием микотоксинов предшествуют функциональные нарушения, которые влияют на ферментативный статус пищеварительного канала и морфо-биохимические показатели крови. Попадая в организм, микотоксины подвергаются биохимическим превращениям. Наиболее полно проанализировать закономерности таких процессов позволяют опыты in vivo на фистульных животных с применением ВЭЖХ-МС/МС для определения микотоксинов в образцах.
В настоящей работе мы использовали такой подход и показали, что при скармливании контаминированного Т-2 токсином комбикорма в кишечнике птицы обнаруживается Т-2 токсин в его метаболит — НТ-2 токсин, причем последний интенсивно всасывается в кровь и, видимо, переходит в ткани, поскольку его количество значительно снижается в помете по сравнению с дуоденумом. Это сопровождается изменениями активности ферментов в дуоденальном химусе, ферментов крови и лейкоцитарной формулы, что свидетельствует о повышении токсической нагрузки на печень и развитии патологического процесса. В качестве маркера для ранней диагностики микотоксикозов мы предлагаем использовать фосфатазно-протеазный индекс, повышение которого в крови до значений более 20 ед. указывает на нарушение пищеварения (30).
Целью работы было определение содержания Т-2 и НТ-2 токсинов в химусе и помете, активности пищеварительных ферментов и морфо-биохимических показателей крови цыплят-бройлеров при экспериментальном Т-2 токсикозе.
Методика. Эксперименты проводили на цыплятах-бройлерах (Gallus gallus L.) кросса Смена 8 в соответствии с требованиями Европейской конвенции о защите позвоночных животных, используемых для экспериментов или в иных научных целях (ETS № 123, Страсбург, 1986) (31). Вся птица с 1- до 47-суточного возраста содержалась в условиях вивария (ФНЦ ВНИТИП РАН, 2020 год) с соблюдением режимов кормления и содержания в соответствии с требованиями для определенной возрастной группы и кросса птицы (32). Фистулирование 20 особей проводили в 20-25-суточном возрасте, канюлю вживляли напротив места впадения в кишечник панкреатических и желчных протоков по авторскому методу (33). В течение 1 сут после операции птицу не кормили, затем применяли нормированное кормление, следя за прохождением химуса в кишечнике. Швы снимали на 5-е сут. Из клинически здоровой птицы сформировали 4 группы по 5 гол. в каждой: I — контрольная, рацион без добавления Т-2 токсина, II — получающая рацион с добавлением Т-2 токсина на границе ПДК (100 мкг/кг), III — 2ПДК (200 мкг/кг), IV — 4ПДК (400 мкг/кг). Корм контаминировали Т-2 токсином до задаваемого уровня ПДК механическим способом с соблюдением требований безопасности персонала. Применяли стандартный Т-2 токсин (порошок с массовой долей основного вещества 99,7±0,3 %, «Romer Labs», Австрия, cat. N 10000310, LOT #S17052T). Свежий корм задавали ежедневно, доступ к воде не ограничивали.
Учетный подготовительный (фоновый) период длился с 26- до 33-суточного возраста птицы, период опыта — 14 сут (с 34- до 47-суточного возраста). Образцы дуоденального химуса и помет для определения содержания Т-2 и НТ-2 токсинов и ферментативной активности отбирали ежесуточно в течение всего эксперимента, включая подготовительный период, и помещали в морозильную камеру при температуре -20 °С. Из полученных образцов готовили объединенные пробы по каждому бройлеру (за подготовительный период и за каждую неделю эксперимента — к возрасту 40 и 47 сут) для последующей пробоподготовки и анализа (для определения Т-2 и НТ-2 токсинов дополнительно объединяли образцы от цыплят-бройлеров в каждой группе). Пробы химуса (1,0-2,0 мл) и помета (5,0 г) собирали ежесуточно в утренние часы, помещали в холодильную камеру при температуре -20 °С, образцы (по 5 г) высушивали в лиофильной сушилке серии TFD («ilShinBioBase Co., Ltd.», Южная Корея) в течение 34 ч при -77,8 °С и давлении 5 mTorr (удаление 97 % влаги из субстрата с сохранением биологически активных веществ). Кровь (2 мл) для исследования брали перед началом опыта и в конце каждой недели опыта (в возрасте птицы 33, 40 и 47 сут) из подкрыльцовой вены (cutanea ulnaris) на внутренней стороне крыла над локтевым сочленением. Место пункции на несколько минут зажимали стерильным тампоном. Образцы для биохимических исследований отбирали в стерильные вакуумные пробирки с литий-гепарином (объем 4,0 мл, «Shandong Weigao Group Medical Polymer Co., Ltd.», Китай), для морфологических — с антикоагулянтом K3-EDTA (объем 2,0 мл, «SOYAGREENTEC Co., Ltd.», Южная Корея). Для отделения плазмы от форменных элементов пробы центрифугировали при 5000 об/мин в течение 5 мин.
Пробы корма для анализа отбирали согласно ГОСТ 13496.0-2016 «Комбикорма, комбикормовое сырье. Методы отбора проб» (М., 2020) из мешков. Арбитражная проба готовилась из средней пробы массой 1 кг. Каждую пробу корма анализировали по 3 раза. Пробоподготовку осуществляли согласно ГОСТ 34140-2017 «Продукты пищевые, корма, продовольственное сырье. Метод определения микотоксинов с помощью высокоэффективной жидкостной хроматографии с масс-спектрометрическим детектированием» (М., 2020).
Количественное содержание Т-2 и НТ-2 токсинов в исходном комбикорме, дуоденальном химусе и помете измеряли (дважды в каждой анализируемой пробе) методом тандемной высокоэффективной жидкостной хромато-масс-спектрометрии (ВЭЖХ-МС/МС) (хроматографическая система Agilent 1260 Infinity, «Agilent Technologies», Германия, масс-спектрометр AB SCIEX Triple QuadTM 5500, «Applied Biosystems», США; колонка Gemini® с обращенно-фазовым сорбентом на основе силикагеля с органическим полимером С18 5 mm 150×4,6 mm, «Phenomenex», США). Для построения калибровочных графиков и в качестве внутренних стандартов использовали стандартные растворы соответствующих микотоксинов (Т-2 и НТ-2 токсины) серии Biopure («Romer Labs», Австрия). При проведении анализов руководствовались ГОСТ 34140-2017. Использовали метанол, ацетонитрил (чистота ВЖХ-МС, массовая доля основного вещества не менее 99,9 %) («Merk», Германия); ледяную уксусную кислоту, ацетат аммония (массовая доля основного вещества не менее 99,0 %) («Sigma», Германия). Элюент А: 890 см3 деионизированной воды, 100 см3 метанола, 10 см3 уксусной кислоты и 0,384 г ацетата аммония (5 ммоль/л). Элюент Б: 970 см3 метанола, 20 см3 деионизированной воды, 10 см3 уксусной кислоты и 0,384 г ацетата аммония (5 ммоль/л). Раствор для экстракции — ацетонитрил:вода:уксусная кислота 79:20:1. Хроматографическое разделение проводили в режиме бинарного градиентного элюирования: 1,5 мин 100 % элюэнта A, линейное увеличение доля элюента B до 50 % в течение 1,5 мин, линейное увеличение доли элюента B до 100 % в течение 9 мин. Время хроматографического разделения со 100 % элюента В — 5 мин, последующее уравновешивание колонки — 3 мин, 100 % элюента A. Температура колонки — 25 °С; скорость подвижной фазы — 1 см3/мин; температура ввода проб — 10 °С, объем вводимой пробы 5 мм3. Время удерживания для Т-2 токсина и НТ-2 токсина — соответственно 10,30 и 9,79 мин; предел обнаружения для Т-2 токсина и НТ-2 токсина — 3,25 и 2,70 мкг/кг, предел определения — 5,23-129,20 и 3,50-129,20 мкг/кг. Расчетный ион (в режиме положительной ионизации распылением в электрическом поле) для Т-2 токсина — 305,1 m/z, для НТ-2 токсина — 345,1 m/z.
Амилазу определяли по Smith-Roy в модификации для высокой активности фермента (34), активность протеаз — по гидролизу казеина очищенного по Гаммерстену (калориметрический контроль при λ = 450 нм), липазу — на полуавтоматическом биохимическом анализаторе SINNOWA BS-3000P («SINNOWA Medical Science & Technology Co., Ltd», КНР) с набором ветеринарных диагностических реагентов («ДИАКОН-ВЕТ», Россия).
Биохимические исследования крови выполняли на полуавтоматическом биохимическом анализаторе BS-3000P (КНР) с набором для определения общего белка, щелочной фосфатазы, глюкозы, холестерина, триглицеридов, липазы («ДИАКОН-ВЕТ», Россия). Активность трипсина в плазме крови измеряли на полуавтоматическом биохимическом анализаторе BS-3000P (КНР) кинетическим методом (35) с использованием в качестве субстрата Na-бензоил-DL-аргинин-п-нитроанилид (BAPNA, «ACROS ORGA-NICS», Швейцария).
Морфологические исследования крови выполняли на автоматическом гематологическом анализаторе для ветеринарии DF-50 («Dymind Biotech», КНР) с применением фирменных реагентов.
Для статистической обработки результатов применяли программное обеспечение JMP Trial 14.1.0 (https://www.jmp.com/en_us/software/data-ana-lysis-software.html). Результаты представлены в виде М±SD, где М — среднее арифметическое, ±SD — среднеквадратичное отклонение. Достоверность различий устанавливали по t-критерию Стьюдента, различия считали статистически значимыми при p < 0,05.
Результаты. Попадая в пищеварительный канал, микотоксины метаболизируются и в таком виде поступают в кровь. Известно, что в организме метаболизм Т-2 токсина происходит в реакциях деацетилирования, гидроксилирования и конъюгации (2, 16). Ранее мы показали (36), что у птиц Т-2 токсин превращается в НТ-2 токсин и существенно влияет на концентрацию оксида азота и активность панкреатических ферментов. В продолжение в настоящей работе данные химико-токсикологических и физиолого-биохимических исследований дополнены результатами общего и биохимического анализа крови в качестве клинических показателей статуса птицы при экспериментальном Т-2 токсикозе. Кроме того, нами была применена лиофильная сушка образцов для улучшения пробоподготовки.
Определение содержания Т-2 токсина и НТ-2 токсина в корме, дуоденальном химусе и помете контрольной фистулированной птицы (табл.1) показало, что в корме до контаминации присутствовали Т-2 и НТ-2 токсины в сопоставимых количествах — соответственно 4,3±0,19 и 5,6±0,47 мкг/кг. У цыплят, получавших неконтаминированный рацион, эти токсины были найдены в химусе и помете в других соотношениях. В среднем за период эксперимента в контрольной группе в помете по сравнению с дуоденумом количество Т-2 токсина повышалось в 5 раз (p < 0,05), что указывает на удаление из организма большей части токсина в неизменном виде. Однако содержание НТ-2 токсина в дуоденуме, превышающее в 6 раз таковое для Т-2 токсина, указывает на то, что в пищеварительном канале Т-2 токсин активно превращался в метаболит, способный всасываться в кровь. В помете мы наблюдаем такое же количество НТ-2 токсина, как и в дуоденальном химусе, за исключением IV группы. При увеличении количества Т-2 токсина в корме до 100 мкг/кг корма (1ПДК) наблюдается резкое (в 40 раз) повышение содержания токсина в химусе по сравнению с контролем, однако в помете его выделение сохраняется на уровне контрольной пробы. Это можно объяснить переходом Т-2 токсина в НТ-2 форму с дальнейшим поступлением в кровь и ткани цыплят, поскольку уровень НТ-2 токсина в помете существенно не изменяется. Увеличение дозы токсина в корме до 200 мкг/кг сопровождалось повышением содержания Т-2 токсина в химусе в 2,4 раза (p < 0,05), а в помете — в 2,1 раза по сравнению с 1ПДК. Содержание НТ-2 токсина увеличивалось соответственно на 20,9 % (p < 0,05) и на 20,8 % (p > 0,05). Следовательно, соотношение Т-2 токсина и его метаболита в кишечнике и помете указывает на интенсивное всасывание НТ-2 токсина в кровь. При дозировке Т-2 токсина 400 мкг/кг его количество в помете увеличивалось по сравнению с контролем в 7,7 раза, что свидетельствует об адаптационных изменениях в кишечнике птицы, которые обеспечивают более интенсивное выведение токсина из кишечника, возможно, за счет более быстрого продвижения химуса в каудальном направлении. Клинически это проявляется расстройствами пищеварения и энтеритами с признаками диареи.
В нашем эксперименте, выполненном ранее, показано, что микотоксины в больших дозах оказывают отрицательное действие на активность пищеварительных ферментов (37). В продолжение исследований в настоящей работе мы определили активность пищеварительных ферментов в лиофилизированном дуоденальном содержимом и помете (табл. 2). Контаминация корма Т-2 токсином повлияла на протеолитическую активность дуоденального химуса, увеличивая ее во II группе на 78,9 % (p < 0,05), в III группе — на 46,3 % (p < 0,05), в IV — на 96,6 % (p < 0,05) по сравнению с подготовительным (фоновым) периодом. Повышение активности липазы во II и III группах были достоверны — соответственно на 25,5 и 16,8 % (p < 0,05). Активность амилазы увеличивалась в IV группе на 99,7 % (p < 0,05). Активность щелочной фосфатазы в отличие от пищеварительных ферментов имела тенденцию снижаться (во II группе — на 23,8 %, в III — на 27,9 %) или не изменялась (в IV группе).
Индивидуальные изменения активности протеаз, амилазы, трипсина и липазы в дуоденальном химусе цыплят-бройлеров, определяющие особенности их реакции на разные дозы Т-2 токсина в корме, представлены на сайте (http://www.agrobiology.ru, рис. 1). У большинства особей характерным ответом на введение в корм токсина было повышение активности протеолитических ферментов.
Биохимический анализ помета (см. табл. 2) выполняли после лиофильной сушки, полностью сохраняющей пищеварительные ферменты в активном состоянии. Результаты показали, что активность амилазы уменьшалась во II и III опытных группах соответственно на 55,1 и 57,2 % по сравнению с фоновым периодом. Активность липазы снижалась при максимальной дозировке токсина на 23,2 % (p < 0,05). Протеолитическая активность в помете значительно повышалась по мере увеличения дозы Т2 токсина в корме — соответственно на 136,4 %, 76,0 % и 169,1 % относительно фонового периода.
То есть активность протеаз в дуоденальном содержимом значительно превышает показатели в помете: во II группе в фоновый период в 2,5 раза, в опытный — в 1,9 раза; в III — соответственно в 3,2 и 2,7 раза, в IV — в 3,4 и 2,5 раза. Значительное снижение активности протеаз по мере продвижения по пищеварительному каналу можно объяснить возвращением ферментов в кровь согласно гипотезе о рекреции панкреатических ферментов (38, 39) либо их разрушением микрофлорой кишечника. Следует отметить стимулирующее действие микотоксинов на активность протеаз содержимого 12-перстной кишки. Кроме того, высокая протеолитическая активность со-храняется в помете птиц, что указывает на проявление признаков расстройства пищеварения, так как при усилении моторики кишечника происходит удаление избытка ферментов с пометом. Следовательно, при поступлении с кормом относительно небольших количеств токсина организм адаптируется и на первом этапе реагирует повышением протеолитической активности кишечных ферментов.
С учетом того, что активность протеаз кишечника коррелирует с активностью ферментов в крови, клинический тест на выявление Т-2 токсикоза может включать не только оценку протеолитической активности в экскрементах, но и изменение активности трипсина в плазме крови бройлеров (табл. 3).
Согласно данным биохимических исследований крови, активность трипсина при экспериментальном Т-2 токсикозе снижалась с повышением дозы Т-2 токсина (наиболее заметно при 4ПДК) (см. табл. 3). Активность щелочной фосфатазы имела обратную динамику: при 4ПДК за 2 нед этот показатель возрастал в 5 раз с увеличением фосфатазно-протеазного индекса в среднем с 31 до 92 ед., что свидетельствует о снижении секреторной функции поджелудочной железы, уменьшении доступности питательных веществ и повышении токсической нагрузки на печень (40). При 4ПДК в крови цыплят содержание общего белка повысилось на 13,2 %, но количество глюкозы снизилось на 13,5 %, а триглицеридов в крови — в 4 раза, что указывает на наличие стрессовой реакции при поступлении Т-2 токсина с кормом.
Результаты выполненного нами исследования морфологических показателей крови цыплят отражают характер наблюдаемых гематологических отклонений при экспериментальном микотоксикозе (табл. 4). Число лейкоцитов значительно снизилось при 2ПДК Т-2 токсина в корме — на 15,8 % у 40-суточных и на 16,6 % у 47-суточных цыплят-бройлеров. Однако этот показатель оставался в пределах физиологической нормы, поэтому говорить о снижении иммунитета преждевременно. В лейкоцитарной формуле при 2ПДК и 4ПДК наблюдалась тенденция к увеличению числа лимфоцитов по сравнению с псевдоэозинофилами, что связано со специфической защитой организма при инфекционных и неинфекционных патологиях. Доля эозинофилов при 2ПДК снижалась, в остальных группах — повышалась по сравнению с контролем. Доля моноцитов при 1ПДК Т-2 токсина за первую неделю снижалась в 3 раза, затем стабилизировалась на одном уровне. При повышении дозы Т-2 токсина до 2ПДК снижение количества моноцитов было более выраженным (соответственно в 6 и 2 раза) по сравнению с контролем. Максимальная доза токсина (4ПДК) вызывала увеличение количества моноцитов в 1,5 раза в 40-суточном возрасте по сравнению с контрольной группой. Доля базофилов в лейкоцитарной формуле имела тенденцию к снижению в 47-суточном возрасте цыплят-бройлеров во II и III группах (в 2 раза по сравнению с контролем).
Отмеченное нами увеличение количества НТ-2 токсина в химусе и помете цыплят-бройлеров при повышении дозы Т-2 токсина до 2ПДК (см. табл.1) указывает на активный переход этого токсина из одной формы в другую. При этом количество Т-2 токсина в химусе 12-перстной кишки увеличилось в 2,5 раза, а НТ-2 токсина — в 1,2 раза по сравнению со II группой (1ПДК Т-2 токсина). Одновременно активность трипсина, который участвует не только в переваривании белков, но и в регуляции метаболизма (35), в 12-перстной кишке уменьшалась, а в помете — увеличивалась при незначительном изменении активности фермента в крови (рис. 2). Это указывает на защитную реакцию пищеварительных желез на действие токсина, которая проявляется в повышении секреции панкреатического и кишечного соков для сохранения энтерального гомеостаза при больших потерях жидкости организмом. В связи с увеличением скорости движения кишечного химуса при адаптации к воздействию токсина нарушается процесс возврата ферментов в кровь, и часть их выделяется вместе с пометом (36). При повышении дозы Т-2 токсина в корме бройлеров до 4ПДК отмечалось снижение количества Т-2 токсина в химусе 12-перстной кишки в 2,6 раза при 8,4-кратном увеличении в помете. Адаптация кишечника к Т-2 токсину в дозе 4ПДК имела существенные отличия от наблюдаемой при меньших дозах токсина: активность трипсина в дуоденальном химусе увеличивалась при практически постоянном содержании фермента в помете (см. рис. 2).
При сопоставлении метаболизации Т-2 токсина в НТ-2 токсин (соотношение количества НТ-2 токсина и Т-2 токсина) с изменением активности трипсина в химусе проявилась обратная зависимость между этими процессами, особенно при дозе Т-2 токсина 4ПДК (рис. 3). Также нами установлена обратная зависимость между фосфатазно-протеазным индексом крови и количественным соотношением НТ-2 токсин/Т-2 токсин в дуоденальном химусе при 4ПДК Т-2 токсина в корме, что может быть использовано при диагностике Т-2 микотоксикозов (см. рис. 3).
Таким образом, показано, что при экспериментальном Т-2 токсикозе в пищеварительном канале цыплят-бройлеров Т-2 токсин преобразуется в свой метаболит — НТ-2 токсин, причем последний интенсивно всасывается в кровь и переходит, видимо, в ткани, поскольку его количество значительно (на 28,7 %) снижается в помете по сравнению с дуоденумом. При добавлении Т-2 токсина в корм (1ПДК, 2ПДК и 4ПДК — 100, 200 и 400 мкг/кг) протеолитическая активность в дуоденальном химусе возрастает соответственно на 78,9 % (p < 0,05), 46,3 % (p < 0,05) и 96,6 % (p < 0,05), то есть в наибольшей степени при 4ПДК. Здесь же отмечена максимальная активность амилазы (2-кратное увеличение). Повышение активность липазы (меньшее, но достоверное — на 25,5 и 16,8 % при p < 0,05) происходило при скармливании 1ПДК и 2ПДК Т-2 токсина. Активность щелочной фосфатазы, наоборот, имела тенденцию снижаться (1ПДК и 2ПДК) либо не изменялась. В крови активность трипсина при экспериментальном Т-2 токсикозе снижалась на 12,2-22,2 % (II, IV группы) в 40-суточном возрасте цыплят и на 22,1 % (III группа) в 47-суточном, а активность щелочной фосфатазы, наоборот, возрастала (5-кратно за 2 нед при скармливании 4ПДК Т-2 токсина). Увеличение фосфатазно-протеазного индекса свидетельствовало о повышении токсической нагрузки на печень. Число лейкоцитов крови снижалось при 1ПДК в среднем на 16 %. В лейкоцитарной формуле отмечали тенденцию к увеличению доли лимфоцитов при нагрузке 1ПДК и 2ПДК в отличие от псевдоэозинофилов, что связано со специфической защитой организма от токсинов. Установлена обратная зависимость фосфатазно-протеазного индекса крови (ФПИ) от количественного соотношения НТ-2 и Т-2 токсинов в химусе при 4ПДК Т-2 токсина в корме. Выявлено повышение ФПИ при высоких дозах Т-2 токсина в корме. Определение ФПИ может служить тестом при диагностике Т-2 токсикозов птицы.
1. Концентрация микотоксинов (мкг/кг) в дуоденальном химусе и помете цыплят-бройлеров (Gallus gallus L.) кросса Смена 8 при экспериментальном Т-2 токсикозе (M±SD, виварий ФНЦ ВНИТИП РАН, 2020 год) |
||||
Микотоксин |
Группа (n = 5) |
|||
I (контроль без Т-2 токсина) |
II (Т-2 токсин 100 мкг/кг корма) |
III (Т-2 токсин 200 мкг/кг корма) |
IV (Т-2 токсин 400 мкг/кг корма) |
|
|
Дуоденальный химус |
|||
Т-2 токсин |
0,5±0,13 |
20,2±2,75* |
49,6±6,00* |
41,9±6,36* |
НТ-2 токсин |
30,0±4,90 |
47,9±1,60* |
57,9±3,20* |
53,9±1,45* |
|
Помет |
|||
Т-2 токсин |
2,5±0,26 |
2,3±0,40 |
4,8±0,68* |
19,2±6,20* |
НТ-2 токсин |
40,6±8,40 |
44,6±3,26 |
53,9±3,60 |
38,5±5,67 |
Примечание. В исходном корме (без контаминации) измеренное содержание Т-2 токсина составило 4,3±0,19 мкг/кг, НТ-2 токсина — 5,6±0,47 мкг/кг. Образцы химуса и помета от каждого бройлера отбирали ежедневно в течение 14 сут опыта, объединяли в недельные пробы (возраст 40 и 47 сут; начало опыта с 34-суточного возраста). По каждой группе из недельных проб формировали объединенные лиофилизированные пробы. Концентрации аналитов определяли методом тандемной высокоэффективной жидкостной хромато-масс-спектрометрии (ВЭЖХ-МС/МС) (каждое измерение выполнялось трижды, хроматографическая система Agilent 1260 Infinity, «Agilent Technologies», Германия, масс-спектрометр AB SCIEX Triple QuadTM 5500, «Applied Biosystems», США). Представлены средние (M) по недельным пробам по группам. * Различия с контрольной группой статистически значимы при p < 0,05. |
||||
2. Активность пищеварительных ферментов в дуоденальном химусе и помете цыплят-бройлеров (Gallus gallus L.) кросса Смена 8 при экспериментальном Т-2 токсикозе (M±SD, виварий ФНЦ ВНИТИП РАН, 2020 год) |
||||||
Показатель |
Группа (n = 5) |
|||||
II (Т-2 токсин, 100 мкг/кг корма) |
III (Т-2 токсин, 200 мкг/кг корма) |
IV (Т-2 токсин, 400 мкг/кг корма) |
||||
фоновый период |
опыт |
фоновый период |
опыт |
фоновый период |
опыт |
|
Дуоденальный химус |
||||||
Амилаза, мг/(г·мин) |
6433±446,0 |
8100±861,2 |
8550±1670,0 |
5450±437,5 |
5033±314,1 |
10050±875,2* |
Липаза, мкмоль/(г·мин) |
29,4±2,59 |
36,9±1,66* |
25,6±0,49 |
29,9±1,29* |
22,0±3,43 |
26,1±0,35 |
Протеазы, мг/(г·мин) |
190±18,7 |
340±21,2* |
229±22,8 |
335±27,6* |
233±13,5 |
458±56,3* |
Трипсин, мкмоль/(г·мин) |
4,92±0,364 |
6,86±0,354 |
5,27±0,581 |
5,84±0,196 |
6,47±0,589 |
9,33±2,045 |
Щелочная фосфатаза, мкмоль/(г·мин) |
147,5±13,64 |
112,4±5,19* |
134,6±1,88 |
97,1±7,18* |
140,0±2,11 |
188,8±37,62 |
Фосфатазно-протеазный индекс |
29,9 |
16,4 |
25,5 |
16,6 |
21,6 |
20,2 |
Помет |
||||||
Амилаза, мг/(г·мин) |
1413±111,0 |
635±67,1* |
1890±275,1 |
810±82,0* |
1145±153,1 |
1140±94,2 |
Липаза, мкмоль/(г·мин) |
1802±54,0 |
1946±316,0 |
913±6,6 |
880±43,0 |
1225±43,0 |
941±33,0* |
Протеазы, мг/(г·мин) |
77±10,3 |
182±22,1* |
71±10,0 |
125±6,5* |
68±6,3 |
183±4,6* |
Трипсин, мкмоль/(г·мин) |
4,38±0,349 |
2,72±0,175 |
3,79±0,233 |
3,32±0,242 |
3,39±0,313 |
3,00±0,382 |
Щелочная фосфатаза, мкмоль/(г·мин) |
211,6±10,08 |
140,8±30,98* |
104,9±8,55 |
77,5±4,92* |
82,8±4,19 |
143,1±14,08* |
Фосфатазно-протеазный индекс |
48,3 |
51,8 |
31,6 |
23,3 |
24,2 |
47,7 |
Примечание. В исходном корме (без контаминации) измеренное содержание Т-2 токсина составило 4,3±0,19 мкг/кг, НТ-2 токсина — 5,6±0,47 мкг/кг. В физиологическом опыте (метод групп-периодов) образцы химуса и помета от каждого бройлера отбирали ежедневно до начала опыта (фоновый период с 26- до 33-суточного возраста) и в течение 14 сут опыта (с 34- до 47-суточного возраста), образцы, полученные в опытный период, объединяли в недельные пробы (возраст 40 и 47 сут). * Различия с подготовительным (фоновым) периодом статистически значимы при p < 0,05. |
||||||
3. Биохимические показатели крови у цыплят-бройлеров (Gallus gallus L.) кросса Смена 8 при экспериментальном Т-2 токсикозе (M±SD, виварий ФНЦ ВНИТИП РАН, 2020 год) |
||||
Показатель |
Группа (n = 5) |
|||
I (контроль без Т-2 токсина) |
II (Т-2 токсин, 100 мкг/кг корма) |
III (Т-2 токсин, 200 мкг/кг корма) |
IV (Т-2 токсин, 400 мкг/кг корма) |
|
Активность трипсина, ед/л |
93,3±4,36 77,5±6,43 |
81,9±1,58* 68,6±3,02 |
82,5±1,12 60,4±3,89* |
72,6±0,31* 66,3±7,65 |
Активность щелочной фосфатазы, ед/л |
4179±456,1 1298±192,5 |
4310±609,7 754±37,7 |
4370±476,2 1304±118,8 |
6369±757,8* 6487±570,5* |
Фосфатазно-протеазный индекс |
45 17 |
53 11 |
53 21 |
87 98 |
Общий белок, г/л |
40,1±0,71 36,3±1,20 |
40,3±1,23 36,4±0,13 |
40,4±0,19 35,5±0,21 |
41,4±0,16 40,1±0,85* |
Глюкоза, ммоль/л |
12,4±0,24 10,4±0,32** |
11,3±0,33 9,8±0,16** |
12,9±0,07 10,0±0,16** |
13,0±0,21 9,0±0,31*, ** |
Триглицериды, ммоль/л |
0,3±0,01 0,8±0,04** |
0,3±0,02 0,4±0,02 |
0,4±0,02 0,4±0,15 |
0,4±0,28 0,2±0,01 |
Холестерин, ммоль/л |
3,4±0,04 2,8±0,14 |
3,1±0,05* 3,1±0,60 |
3,6±0,25 3,1±0,26 |
3,6±0,21 2,8±0,04** |
Примечание. Над чертой — возраст бройлеров 40 сут, под чертой — 47 сут. * Различия с I (контрольной) группой статистически значимы при p < 0,05. ** Различия между 47- и 40-суточными цыплятами статистически значимы при p < 0,05. |
||||
4. Лейкоциты крови и лейкоцитарная формула у цыплят-бройлеров (Gallus gallus L.) кросса Смена 8 при экспериментальном Т-2 токсикозе (M±SD, виварий ФНЦ ВНИТИП РАН, 2020 год) |
||||
Показатель |
Группа (n = 5) |
|||
I (контроль без Т-2 токсина) |
II (Т-2 токсин, 100 мкг/кг корма) |
III (Т-2 токсин, 200 мкг/кг корма) |
IV (Т-2 токсин, 400 мкг/кг корма) |
|
Лейкоциты, 109/л |
32,2±0,90 30,8±1,25 |
27,1±0,61* 25,7±0,23* |
29,6±0,70 28,1±0,90 |
32,7±1,65 28,6±2,88 |
Псевдоэозинофилы, % |
44,8±1,49 41,1±2,46 |
33,9±1,95* 34,2±0,98 |
36,0±1,23* 45,2±6,24 |
47,8±3,05 36,6±1,70 |
Лимфоциты, % |
48,8±1,70 52,4±2,22 |
62,3±2,24* 59,9±0,41* |
58,9±1,73* 47,7±7,56 |
41,0±4,02 55,2±0,29 |
Моноциты, % |
0,6±0,08 0,4±0,07 |
0,2±0,03* 0,3±0,10 |
0,1±0,05* 0,2±0,06 |
0,9±0,05* 0,5±0,09 |
Эозинофилы, % |
5,5±0,42 5,7±0,75 |
3,1±0,28* 5,3±0,50 |
4,7±0,48 6,6±1,24 |
9,7±1,05* 7,2±1,36 |
Базофилы, % |
0,4±0,04 0,4±0,06 |
0,4±0,05 0,2±0,03* |
0,2±0,01* 0,2±0,03* |
0,4±0,03 0,3±0,03 |
Примечание. Над чертой — возраст бройлеров 40 сут, под чертой — 47 сут. * Различия с I (контрольной) группой статистически значимы при p < 0,05. |
||||
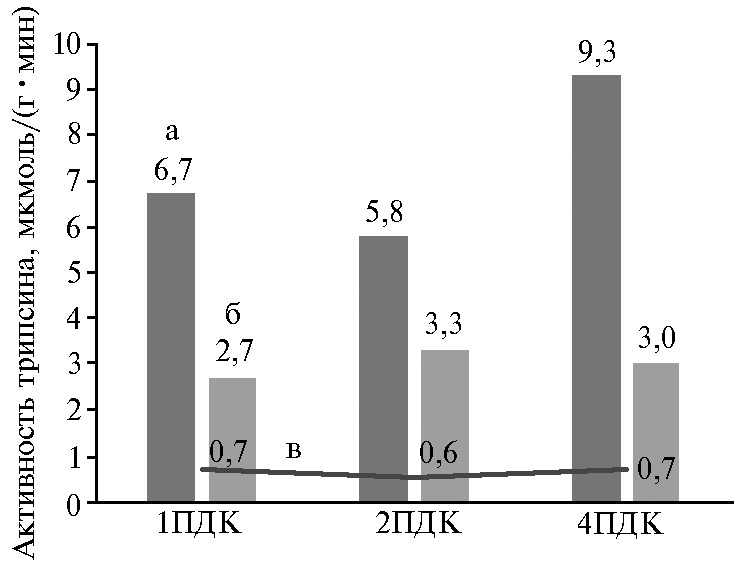
Рис. 2. Активность трипсина в разных биологических средах при экспериментальном Т-2 токсикозе цыплят-бройлеров (Gallus gallus L.) кросса Смена 8 (среднее за опыт для дозы токсина): а — дуоденальный химус, б — помет, в — кровь; 1ПДК — 100 мкг/кг корма, 2ПДК — 200 мкг/кг корма, 4ПДК — 400 мкг/кг корма (виварий ФНЦ ВНИТИП РАН, 2020 год).
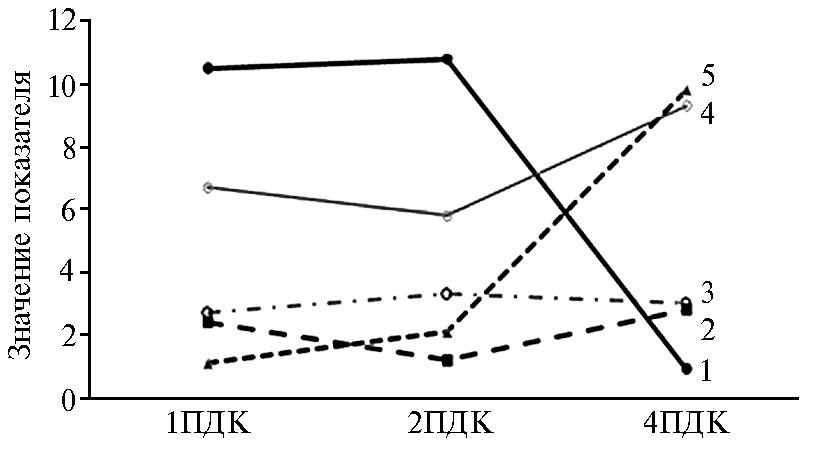
Рис. 3. Количественное соотношение НТ-2 токсин/Т-2 токсин в дуоденальном химусе (1) и помете (2), активность трипсина в помете, ед/мл (3) и дуоденальном химусе, ед/мл (4) и фосфатазно-протеазный индекс крови (5) у цыплят-бройлеров (Gallus gallus L.) кросса Смена 8 при экспериментальном Т-2 токсикозе: 1ПДК — 100 мкг/кг корма, 2ПДК — 200 мкг/кг корма, 4ПДК — 400 мкг/кг корма (виварий ФНЦ ВНИТИП РАН, 2020 год).
ЛИТЕРАТУРА
- Awuchi C.G., Ondari E.N., Ogbonna C.U., Upadhyay A.K., Baran K., Okpala C.O.R., Korzeniowska M., Guiné R.P.F. Mycotoxins affecting animals, foods, humans, and plants: types, occurrence, toxicities, action mechanisms, prevention, and detoxification strategies — a revisit. Foods, 2021, 10(6): 1279 (doi: 10.3390/foods10061279).
- Иванов А.В., Фисинин В.И., Тремасов М.Я., Папуниди К.Х. Микотоксикозы (биологические и ветеринарные аспекты). М., 2010.
- Hollander D., Croubels S., Lauwers M., Caekebeke N., Ringenier M., Meyer F.D., Reisinger N., Immerseel F.V., Dewulf J., Antonissen G. Biomonitoring of mycotoxins in blood serum and feed to assess exposure of broiler chickens. Journal of Applied Poultry Research, 2021, 30(1): 100111 (doi: 10.1016/j.japr.2020.10.010).
- Pereira C.S., Cunha S.C., Fernandes J.O. Prevalent mycotoxins in animal feed: occurrence and analytical methods. Toxins (Basel), 2019, 11(5): 290 (doi: 10.3390/toxins11050290).
- Marin S., Ramos A.J., Cano-Sancho G., Sanchis V. Mycotoxins: occurrence, toxicology, and exposure assessment. Food Chem. Toxicol., 2013, 60: 218-237 (doi: 10.1016/j.fct.2013.07.047).
- Kovalsky P., Kos G., Nährer K., Schwab C., Jenkins T., Schatzmayr G., Sulyok M., Krska R. Co-occurrence of regulated, masked and emerging mycotoxins and secondary metabolites in finished feed and maize — an extensive survey. Toxins, 2016, 8: 363 (doi: 10.3390/toxins8120363).
- Streit E., Schatzmayr G., Tassis P., Tzika E., Marin D., Taranu I., Tabuc C., Nicolau A., Aprodu I., Puel O., Oswald I. P. Current Situation of mycotoxin contamination and co-occurrence in animal feed—focus on Europe. Toxins (Basel), 2012, 4(10): 788-809 (doi: 10.3390/toxins4100788).
- Biomin® mycotoxin prediction. Режим доступа: https://www.biomin.net/science-hub/. Дата обращения 20.08.2021.
- Labuda R., Parich A., Berthiller F., Tančinová D. Incidence of trichothecenes and zearalenone in poultry feed mixtures from Slovakia. Int. J. Food Microbiol.,2005, 105: 19-25 (doi: 10.1016/j.ijfoodmicro.2005.06.005).
- Ibáñez-Vea M., Lizarraga E., González-Peñas E., López de Cerain A. Co-occurrence of type-A and type-B trichothecenes in barley from a northern region of Spain. Food Control, 2012, 25: 81-88 (doi: 10.1016/j.foodcont.2011.10.028).
- Головня Е.Я., Лунегова И.В., Свиридова А.В. Мониторинг и определение микотоксинов в комбикормах в Ленинградской области. Международный вестник ветеринарии, 2016, 4: 62-65.
- Джатдоева А.А., Селимов Р.Н., Грачева Т.С., Метальников П.С., Комаров А.А. Исследование загрязненности микотоксинами кормов и кормового сырья на территории Российской Федерации. Успехи медицинской микологии, 2018, 19: 297-298.
- Jing J.L., Zhang Y., Sun H., Wei J.T., Khalil M.M., Wang Y.W., Dai J.F., Zhang N.Y., Qi D.S., Sun L.H. The response of glandular gastric transcriptome to T-2 toxin in chicks. Food Chem. Toxicol., 2019, 132: 110658 (doi: 10.1016/j.fct.2019.110658).
- Sun Y.X., Yao X., Shi S.N., Zhang G.J., Xu L.X., Liu Y.J., Fang B.H. Toxicokinetics of T-2 toxin and its major metabolites in broiler chickens after intravenous and oral administration. J. Vet. Pharmacol. Ther., 2015, 38(1): 80-85 (doi: 10.1111/jvp.12142).
- Yang L., Tu D., Zhao Z., Cui J. Cytotoxicity and apoptosis induced by mixed mycotoxins (T-2 and HT-2 toxin) on primary hepatocytes of broilers in vitro. Toxicon, 2017, 129: 1-10 (doi: 10.1016/j.toxicon.2017.01.001).
- Wan Q., He Q., Deng X., Hao F., Tang H., Wang Y. Systemic metabolic responses of broiler chickens and piglets to acute t-2 toxin intravenous exposure. J. Agric. Food Chem., 2016, 64(3): 714-723 (doi: 10.1021/acs.jafc.5b05076).
- Yang L., Yu Z. Hou J., Deng Y., Zhou Z., Zhao Z., Cui J. Toxicity and oxidative stress induced by T-2 toxin and HT-2 toxin in broilers and broiler hepatocytes. Food Chem. Toxicol., 2016, 87: 128-137 (doi: 10.1016/j.fct.2015.12.003).
- Chen Y., Han S., Wang Y., Li D., Zhao X., Zhu Q., Yin H. Oxidative stress and apoptotic changes in broiler chicken splenocytes exposed to t-2 toxin. BiomedReserch International, 2019, 25: 5493870 (doi: 10.1155/2019/5493870).
- Lauwers M., De Baere S., Letor B., Rychlik M., Croubels S., Devreese M. Multi LC-MS/MS and LC-HRMS methods for determination of 24 mycotoxins including major phase I and II biomarker metabolites in biological matrices from pigs and broiler chickens. Toxins, 2019, 11(3): 171 (doi: 10.3390/toxins11030171).
- Xue C.Y., Wang G.H., Chen F., Zhang X.B., Bi Y.Z., Cao Y.C. Immunopathological effects of ochratoxin A and T-2 toxin combination on broilers. Poultry Science, 2010. 89(6): 1162-1166 (doi: 10.3382/ps.2009-00609).
- Kubena L.F., Edrington T.S., Harvey R.B., Buckley S.A., Phillips T.D., Rottinghaus G.E., Casper H.H. Individual and combined effects of fumonisin B1 present in Fusarium moniliforme culture material and T-2 toxin or deoxynivalenol in broiler chicks. Poultry Science, 1997, 76(9): 1239-1247 (doi: 10.1093/ps/76.9.1239).
- Venkatesh P.K., Vairamuthu S., Balachandran C., Manohar B.M., Raj G.D. Induction of apoptosis by fungal culture materials containing cyclopiazonic acid and T-2 toxin in primary lymphoid organs of broiler chickens. Mycopathologia, 2005, 159(3): 393-400 (doi: 10.1007/s11046-004-6271-x).
- Huff W.E., Harvey R.B., Kubena L.F., Rottinghaus G.E. Toxic synergism between aflatoxin and T-2 toxin in broiler chickens. Poultry Science, 1988, 67(10): 1418-1423 (doi: 10.3382/ps.0671418).
- Кононенко Г.П., Буркин А.А. Фузариотоксины в зерновых кормах. Ветеринарная патология, 2002 (2): 128-132.
- Кононенко Г.П., Буркин А.А., Зотова Е.В. Микотоксикологический мониторинг. Сообщение 1. Полнорационные корма для свиней и птицы (2009-2018 гг.). Ветеринария сегодня, 2020, 32: 60-65 (doi: 10.29326/2304-196X-2020-1-32-60-65).
- Кононенко Г.П., Буркин А.А., Зотова Е.В. Микотоксикологический мониторинг. Сообщение 2. Зерно пшеницы, ячменя, овса и кукурузы. Ветеринария сегодня, 2020, 33: 139-145 (doi: 10.29326/2304-196X-2020-1-32-60-65).
- Грозина А.А., Гогина Н.Н., Круглова Л.М. Данные мониторинга содержания микотоксинов в кормах, полученные в период с 2010 по 2018 год при помощи метода иммуноферментного анализа. Материалы XX Международной конференции. Российское отделение Всемирной научной ассоциации по птицеводству (ВНАП РФ). Сергиев Посад, 2020: 211-213.
- Lauwers M., De Baere S., Letor B., Rychlik M., Croubels S., Devreese M. Multi LC-MS/MS and LC-HRMS methods for determination of 24 mycotoxins including major phase I and II biomarker metabolites in biological matrices from pigs and broiler chickens. Toxins, 2019, 11(3): 171 (doi: 10.3390/toxins11030171).
- Yang L., Zhao Z., Wu A., Deng Y., Zhou Z., Zhang J., Hou J. Determination of trichothecenes A (T-2 toxin, HT-2 toxin, and diacetoxyscirpenol) in the tissues of broilers using liquid chromatography coupled to tandem mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci., 2013, 942-943: 88-97 (doi: 10.1016/j.jchromb.2013.10.034).
- Вертипрахов В.Г., Грозина А.А., Кислова И.В. Способ оценки адаптации пищеварения птицы к ингредиентному составу рациона. Патент на изобретение 2742175 C1, 02.02.2021. Заявка № 2019142448 от 19.12.2019.
- European Convention for the Protection of Vertebrate Animals used for Experimental and other Scientific Purposes (ETS № 123) (Strasburg, 18.03.1986). Режим доступа: https://norecopa.no/legislation/council-of-europe-convention-ets-123. Accessed: 20.08.2021. Дата обращения 20.08.2021.
- Борисенко К.В., Вертипрахов В.Г. Активность пищеварительных ферментов при добавке в корм бройлеров протеазы. Птицеводство, 2018, 10: 20-23.
- Руководство по оптимизации рецептов комбикормов для сельскохозяйственной птицы /Под ред. В.И. Фисинина. Сергиев Посад, 2014: 3-4.
- Батоев Ц.Ж. Физиология пищеварения птиц. Улан-Удэ, 2001.
- Вертипрахов В.Г., Грозина А.А. Оценка состояния поджелудочной железы методом определения активности трипсина в крови птицы. Ветеринария, 2018, 12: 51-54 (doi: 10.30896/0042-4846.2018.21.12.51-54).
- Вертипрахов В.Г., Титов В.Ю., Гогина Н.Н., Грозина А.А. Изменение активности пищеварительных ферментов и развитие воспаления у цыплят-бройлеров при экспериментальном микотоксикозе. Ветеринария, 2017, 10: 60-63.
- Вертипрахов В.Г., Гогина Н.Н., Грозина А.А., Хасанова Л.В., Ребракова Т.М. Пищеварение и обмен веществ у мясных кур при экспериментальном микотоксикозе. Ветеринария и кормление,2017, 6: 17-20.
- Rothman S.S., Liebow C., Isenman L. Conservation of digestive enzymes. Physiological Review,2002, 82(1): 1-18 (doi: 10.1152/physrev.00022.2001).
- Коротько Г.Ф. Рециркуляция ферментов пищеварительных желез. Российский журнал гастроэнтерологии, гепатологии, колопроктологии,2011, 4: 14-21.
- Вертипрахов В.Г., Кислова И.В. Вопросы минерального обмена с участием щелочной фосфатазы у кур-несушек. Птица и птицепродукты, 2020, 1:44-46(doi: 10.30975/2073-4999-2020-22-1-44-46).