doi: 10.15389/agrobiology.2020.4.784rus
УДК 636.2:591.465.12:576:576.3/.7.086.83:591.04
Работа выполнена в соответствии с темой проекта № 18-016-00147А, финансируемого Российским Фондом Фундаментальных исследований (РФФИ).
ФУНКЦИОНАЛЬНАЯ АКТИВНОСТЬ МИТОХОНДРИЙ И СТАТУС ХРОМАТИНА НАТИВНЫХ И ДЕВИТРИФИЦИРОВАННЫХ ООЦИТОВ Bos taurus ПОД ВОЗДЕЙСТВИЕМ НАНОЧАСТИЦ
ВЫСКОДИСПЕРСНОГО КРЕМНЕЗЕМА
Т.И. КУЗЬМИНА1, И.В. ЧИСТЯКОВА1, Д.Н. ТАТАРСКАЯ2
Митохондрии — единственные клеточные компартменты, генерирующие и трансформирующие энергию в клетке. На изменение экстра- и внутриклеточных условий (ионный гомеостаз, степень дегидратации, температура) эти органеллы реагируют одними из первых. При обработке сверхнизкими температурами вследствие перекисного окисления липидов нарушается работа АТФ-синтетазного комплекса (Е.А. Новодержкина с соавт., 2016), а также структура генетического материала. В качестве цитопротекторных соединений могут быть предложены наночастицы высокодисперсного кремнезема (нВДК). Аморфная форма диоксида кремния, или высокодисперсный кремнезем, проявляющий свою биологическую активность через высокую адсорбирующую способность, снижает концентрацию ионов и биополимеров во время дегидратации клетки при криоконсервации (Т.Т. Туров с соавт., 2011). В настоящей работе впервые показано, что при использовании наночастиц высокодисперсного кремнезема в концентрации 0,001 % в технологии витрификации и экстракорпорального созревания девитрифицированных ооцитов (ДВ) коров наблюдается повышение митохондриального потенциала ДВ ооцитов и снижение числа дегенерированных клеток. Цель исследования — идентифицировать характер воздействия наночастиц высокодисперсного кремнезема на функциональную активность митохондрий и статус хроматина в нативных и девитрифицированных ооцитах коров при экстракорпоральном созревании. В экспериментах использовали ооцит-кумулюсные комплексы (ОКК) голштинизированного крупного рогатого скота (Bos taurus). Витрификации подвергались ооциты с гомогенной цитоплазмой, окруженные пятью и более слоями кумулюсных клеток. ОКК, предназначенные для витрификации, обрабатывали тремя растворами криопротекторных агентов (КПА), приготовленными на среде Т-199 с добавлением 10 % сыворотки крови плодов коров (FBS, «HyClone», Великобритания). В состав растворов КПА для витрификации ооцитов в контроле входили: в КПА-1 — 0,7 M диметилсульфоксид (DMSO) и 0,9 M этиленгликоль (EG); КПА-2 — 1,4 M DMSO и 1,8 M EG; КПА-3 — 2,8 M DMSO, 3,6 M EG и 0,65 M трегалоза. Растворы КПА, растворы для девитрификации и отмывания ооцитов в опыте дополняли наночастицами высокодисперсного кремнезема (нВДК, 4-17 нм, массовая концентрация 0,001 %), синтезированными посредством высокотемпературного гидролиза. В контроле нативные и девитрифицированные ооциты культивировали в течение 24 ч при 38,5 °C и 90 % влажности, в атмосфере, содержащей 5 % СО2. Среда имела следующий состав: Т-199 + 10 % FBS + 106 клеток/мл гранулезы + 50 нг/мл бычьего пролактина. Для культивирования нативныхи девитрифицированных ооцитов в опыте в эту среду добавляли нВДК (массовая концентрация 0,001 %). Для оценки функционального состояния митохондрий в нативных и ДВ ооцитах использовали зонд MitoTracker Orange CMTMRos («Thermo Fisher Scientific», Великобритания). В серии экспериментов по выявлению воздействия нВДК на ядерное созревание женских гамет ооциты помещали на 5-10 мин в 0,9 % раствор цитрата натрия и с помощью препаровальной иглы механически очищали от кумулюса. Затем клетки переносили на сухое обезжиренное стекло и фиксировали смесью метанол:уксусная кислота (3:1). Суховоздушные препараты окрашивали азур-эозином по Романовскому-Гимзе. При воздействии нВДК в ДВ ооцитах возрастала интенсивность флуоресценции MitoTracker Orange CMTMRos (77±6,3 против 169±12,8 мкA, p < 0,05). Функциональная активность митохондрий ДВ ооцитов, обработанных нВДК, в период от стадии диплотены до стадии метафазы I увеличилась с 169±12,8 до 181±7,7 мкА (p < 0,05), в дальнейшем происходило ее снижение до 141±11,2 мкА, что, вероятно, связано с завершением ядерно-цитоплазматического созревания ооцитов. При оценке статуса хроматина ДВ ооцитов в группе, обработанной нВДК, было отмечено снижение числа клеток с признаками дегенерации хроматина на стадии диплотены и метафазы II по сравнению с интактными ДВ ооцитами (40 против 21 % и 59 против 38 %, p < 0.01), что, возможно, обусловлено процессами репарации ДНК. В целом выявлен положительный эффект 0,001 % нВДК в отношении показателей функциональной активности митохондрий и статуса хроматина в ДВ женских гамет Bos taurus.
Ключевые слова: ооцит, витрификация, наночастицы высокодисперсного кремнезема, функциональная активность митохондрий, MitoTracker Orange CMTMRos, хроматин, Bos taurus.
Совершенствование криопротекторных сред для витрификации ооцитов посредством введения различных соединений, необходимых для сохранения жизнеспособности после процедуры размораживания, продолжает оставаться актуальной проблемой технологии криоконсервации. В последнее время при создании комбинированных криоконсервантов широко применяются биологически активные вещества с наноразмерной структурой, синтезированные из различных соединений микроэлементов и минералов, например гидроксиапатита (HA), диоксида кремния (SiO2), оксида алюминия (Al2O3) и диоксида титана (TiO2) (1, 2). Включение наночастиц в состав криопротекторных агентов повышает эффективность витрификации за счет увеличения теплопроводности витрифицирующих растворов и снижения последствий рекристаллизационных процессов, наблюдаемых при размораживании клеток с использованием смесей стандартных криопротекторных агентов, таких как этиленгликоль и диметилсульфоксид (1, 2).
Высокодисперсный (пирогенный) кремнезем (ВДК) — аморфная форма диоксида кремния с рассеянием размеров сферической формы частиц 4-17 нм (90 % интервала) (3). Ввиду выраженной адсорбционной активности, обусловленной значительной удельной поверхностью (Sуд. = 200 м2/г), наночастицы ВДК способны снижать концентрацию полимеров и ионов во время дегидратации клетки (3, 4), а также связывать внеклеточную воду (5). При этом большой размер агрегатов тонкодисперсного кремнезема и малая плотность частиц в этих агломератах обеспечивают связывание значительного количества внеклеточной воды, которая не подвержена изменениям при взаимодействиях с клетками, в том числе в процессах замораживания/оттаивания (5).
В условиях оксидативного стресса при замораживании наблюдается денатурирующее влияние свободно-радикальных процессов на ион-транспортирующие белки, нарушение синтеза ДНК и образования термотропных дефектов митохондрий, что в конечном счете приводит к гибели клеток (6). Митохондрии обеспечивают клетку АТФ, необходимой для завершения мейотического созревания. Особенности их функционирования детерминируют качество гаметы (7, 8). Аномалии преобразования хроматина ядра клетки при температурозависимом окислительном стрессе главным образом обусловлены нарушением биосинтеза ядерных белков и их связи с ДНК, разрушением нуклеотидов, одно- и двухнитевыми разрывами ДНК (9).
В настоящей работе впервые показано, что при использовании наночастиц высокодисперсного кремнезема в концентрации 0,001 % в технологии витрификации и экстракорпорального созревания девитрифицированных ооцитов (ДВ) коров повышается митохондриальный потенциал ДВ ооцитов и снижается число дегенерированных клеток.
Цель исследования — идентифицировать характер воздействия на-ночастиц высокодисперсного кремнезема на функциональную активность митохондрий и статус хроматина в нативных и девитрифицированных ооцитах коров при экстракорпоральном созревании.
Методика. В экспериментах использовали ооцит-кумулюсные комплексы (ОКК) голштинизированного крупного рогатого скота (Bos taurus). Яичники коров после овариоэктомии доставляли в лабораторию в растворе 0,9 % NaCl, содержащем пенициллин (100 МЕ/мл), стрептомицин (100 мкг/мл) и амфотерицин B (0,25 нг/мл), при температуре 30-35 °С. ОКК аспирировали из фолликулов диаметром 3-8 мм.
Витрификации подвергались ооциты с гомогенной цитоплазмой, окруженные пятью и более слоями кумулюсных клеток. ОКК, предназначенные для витрификации, обрабатывали тремя растворами криопротек-торных агентов (КПА), приготовленными на среде Т-199 с добавлением 10 % fetal bovine serum (FBS, «HyClone», Великобритания). В состав растворов КПА для витрификации ооцитов в контроле (без обработки нано-частицами высокодисперсного кремнезема — нВДК) входили: в КПА-1 — 0,7 M диметилсульфоксид (DMSO) и 0,9 M этиленгликоль (EG); в КПА-2 — 1,4 M DMSO и 1,8 M EG; в КПА-3 — 2,8 M DMSO, 3,6 M EG и 0,65 M трегалоза (Trehalose). Все использованные реагенты, за исключением специально обозначенных в тексте, произведены компанией «Sigma-Aldrich» (США), пластиковая лабораторная посуда — BD Falcon™ (Becton Dickinson and Co., BD Biosciences», США). В опыте в растворы КПА для витрификации ооцитов вносили нВДК (4-17 нм, массовая концентрация 0,001 %), синтезированные в ИХП им. А.А. Чуйко НАН Украины посредством высокотемпературного гидролиза. Наночастицы ВДК использовали в виде стабильной суспензии, состоящей преимущественно из субмикронных агрегатов. При выборе концентрации основывались на данных, представленных разработчиками (10).
ОКК помещали на 30 с в КПА-1, затем на 30 с в КПА-2 и на 20 с в КПА-3. После этого пайеты с ооцитами погружали в сосуды Дьюара с жидким азотом. ОКК извлекали из пайет не ранее, чем через 1 ч, размораживали, последовательно помещая в 0,25 М раствор трегалозы, приготовленный на среде Т-199 с добавлением 10 % FBS (3 мин, 37 °С), 0,19 М раствор трегалозы (3 мин, 37 °С) и 0,125 М раствор трегалозы (3 мин, 37 °С), после чего трижды отмывали в среде Т-199 с 10 % FBS. В контроле перечисленные растворы не содержали нВДК, в опыте их дополняли нВДК с конечной концентрацией 0,001 %.
Нативные и девитрифицированные (ДВ) ооциты культивировали в течение 24 ч при 38,5 °C и 90 % влажности в атмосфере, содержащей 5 % СО2 в cреде следующего состав: Т-199 + 10 % FBS + 106 клеток/мл гранулезы + 50 нг/мл бычьего пролактина. При культивировании нативных и девитрифицированных ооцитов в контроле эта среда не содержала нВДК, в опыте — к ней добавляли нВДК с массовой концентрацией 0,001 % (11).
Для оценки функционального состояния митохондрий в нативных и ДВ ооцитах на разных стадиях мейоза использовали зонд MitoTracker Orange CMTMRos («Thermo Fisher Scientific», Великобритания). ОКК по 15-20 шт. помещали в капли 500 нМ раствора MitoTracker Orange CMTMRos объемом 500 мкл и инкубировали 30 мин в темноте при температуре 37 °С. Затем ооциты отмывали от красителя в фосфатном буферном растворе (ФБР) с добавлением 0,3 % бычьего сывороточного альбумина. Отмытые ооциты очищали от кумулюсных клеток, инкубируя в 0,1 % растворе трипсина при 37 °С в течение 5-10 мин, переносили в раствор Хенкса, содержащий 3,7 % параформальдегида, а затем фиксировали 15 мин при 37 °С. После фиксации ооциты отмывали от параформальдегида в ФБР и помещали на стекла Super frost в капли раствора Hoechst 33258 (2,5 мкг/мл, «Thermo Fisher Scientific», Великобритания).
Измерение интенсивности флуоресценции (ИФ) MitoTracker Orange CMTMRos и оценку ядерного созревания с использованием красителя Hoechst 33258 проводили с помощью флуоресцентного микроскопа ZEISS Axio Lab.A1 и фотометрической насадки («Karl Zeiss», Германия). Длина волн возбуждения для MitoTracker Orange CMTMRos составляла 554 нм, излучения — 576 нм; для Hoechst 33258 — соответственно 352 нм и 461 нм. ИФ MitoTracker Orange CMTMRos измеряли в мкА.
В серии экспериментов по выявлению влияния нВДК на ядерное созревание женских гамет ооциты помещали на 5-10 мин в 0,9 % раствор цитрата натрия и с помощью препаровальной иглы механически очищали от кумулюса. Затем клетки переносили на сухое обезжиренное стекло и фиксировали смесью метанол:уксусная кислота (3:1). Суховоздушные препараты окрашивали азур-эозином по Романовскому-Гимзе.
Результаты обрабатывали методом двухфакторного (two-way) дисперсионного анализа ANOVA c помощью пакета статистической программы SigmaStat («Jandel Scientific Software», США). Результаты представлены в виде средних значений (M) и стандартных ошибок средних (±SEM). Для оценки достоверности различий между сравниваемыми средними значениями использовали t-критерий Стьюдента и критерий χ2 Пирсона с поправкой на правдоподобие. Значимость различий сравниваемых средних значений оценивали при р < 0,05, р < 0,01 и р < 0,001 для 3-5 независимых экспериментов.
Результаты. Ранее мы выявили положительное влияние 0,001 % нВДК на статус хроматина соматических клеток овариальных фолликулов коров (12), характер ооцит-кумулюсных взаимодействий при созревании женских гамет (13), развитие доимплантационных эмбрионов (11). В настоящем исследовании было обнаружено, что введение в состав криопротекторных сред нВДК обусловливало повышение показателей флуоресценции зонда MitoTracker Orange CMTMRos в ДВ ооцитах по сравнению с гаметами, не подвергшимися воздействию нВДК (169±12,8 против 77±6,3 мкА, p < 0,05) (табл., рис. 1).
Через 14 ч (после достижения девитрифицированными ооцитами стадии метафазы I) в присутствии нВДК мы отмечали статистически значимые различия в показателях ИФ MitoTracker Orange CMTMRos между клетками, обработанны-ми и не подвергшимися действию нВДК (181±7,7 против 139±11,7 мкA, p < 0,05). При этом к моменту достижения ооцитами стадии метафазы I ИФ нативных ооцитов не отличалась от показателя ДВ ооцитов, подвергшихся обработке нВДК (соответственно 188±11,3 и 181±7,7 мкA). В ДВ ооцитах отмечалось усиление активности митохондрий со стадии диплотены до метафазы I (p < 0,05) (см. табл.). На завершающих этапах созревания гамет наблюдали общее снижение ИФ MitoTracker Orange CMTMRos во всех исследуемых группах, а показатели митохондриальной активности были минимальными.
При оценке состояния хроматина нативных и ДВ ооцитов на стадии диплотены выявили повышение доли ДВ интактных гамет с признаками дегенерации хромосом по сравнению с клетками, обработанны-ми нВДК (40 против 21 %, p < 0,01) (рис. 2). При последующем культивировании до стадии метафазы I мы не обнаружили значимых различий в количестве ооцитов с нормальным или дегенерированным хроматином между контрольной и обработанными нВДК нативными и ДВ группами гамет (рис. 3, А).
Однако через 24 ч по достижении ДВ ооцитами стадии метафазы II (рис. 4) отмечалось повышение числа клеток с признаками дегенерации ядерного материала среди тех, которые не были обработаны нВДК (59 % против 38 %, p < 0,01, см. рис. 3, Б).
При замораживании функциональная активность митохондрий снижается, что обусловлено нарушением не только работы антиоксидантной системы, но и образованием неселективных митохондриальных пор (МП) (7, 14). Витрификация индуцирует изменения в кальциевых рецепторах на мембране эндоплазматического ретикулума, приводящие к атипичной флуктуации содержания Ca2+, что вызывает нерегулируемое открытие МП (15). При такой спонтанной пермеабилизации мембраны происходит запуск каскада событий апоптоза (7). Полученные нами результаты позволяют предположить, что нВДК оптимизирует процессы, связанные с кальциевой регуляцией функционирования МП и усилением ионообмена между цитоплазмой клетки (3, 6). Оксидативный стресс митохондрий при криоконсервации связан с окислительной модификацией белков (16) и образованием белковых фибриллярных комплексов (17, 18). Эти процессы провоцируют окисление клеточной мембраны, нарушают ионный гомеостаз, функционирование ядерного аппарата, меж- и внутриклеточную передачу сигналов (18). Ранее было показано, что наночастицы размером от 3 до 9 нм, проявляющие свойства квантовых точек, способны тормозить образования продуктов окисления белков (16) и ингибировать формирование агрегатных белковых структур (17, 19). Ультрамалые частицы кремнезема предположительно обладают антиоксидантным и антиагрегационным эффектами, за счет чего происходит сохранение клеточных функций, в том числе функций ядерного аппарата. Также следует отметить, что влияние нВДК на митохондрии клеток может быть обусловлено концентрацией частиц в растворе (20, 21). Увеличение функциональной активности митохондрий в обработанных нВДК и контрольных ДВ ооцитах при переходе от стадии диплотены к стадии метафазы I, возможно, объясняется восстановлением электрохимического градиента и повышенной продукцией АТФ (22), необходимой для завершения ядерно-цитоплазматического созревания ДВ ооцита.
Наночастицы ВДК не оказывают генотоксичного действия (23) и способны инициировать антистрессовый ответ клеток на действие сверхнизких температур через активацию белков репарации ДНК (24, 25). Важно отметить, что нВДК в отношении генотоксичности, как и в случае с цито-токсичностью, обладают размеро- и дозозависимым эффектом (25, 26). В наших опытах воздействие нВДК оказывало положительный эффект на ядерное созревание ДВ ооцитов, что, вероятно, было связано с индукцией антистрессового ответа и ослаблением последствий окислительного стресса, обусловленными ультрамалыми размерами частиц ВДК и их небольшой концентрацией в средах для созревания.
Наблюдаемый эффект снижения митохондриальной активности во всех исследованных группах при культивировании, может быть следствием завершения ядерно-цитоплазматического созревания ооцита с дальнейшим блоком мейоза на стадии метафазы II перед активацией яйцеклетки сперматозоидом. Однако уменьшение ИФ MitoTracker Orange CMTMRos и увеличение доли клеток с дегенерированным хроматином в группе ДВ ооцитов на стадии метафазы II по сравнению с другими экспериментальными группами, по-видимому, свидетельствует также о значительном количестве криоповреждений в структуре митохондрий и хроматина.
Таким образом, при экстракорпоральном созревании девитрифицированных ооцитов коров было выявлено повышение митохондриального потенциала ДВ ооцитов и снижение числа дегенерированных клеток при воздействии наночастиц высокодисперсного кремнезема, что может свидетельствовать об усилении ионообменных и репарационных процессов. Увеличение митохондриальной активности при достижении ДВ ооцитами стадии метафазы I указывает на усиление энергоснабжения клеток в этот период. Общее снижение трансмембранного потенциала в нативных и ДВ ооцитах на стадии метафазы II может быть связано либо с завершением ядерно-цитоплазматического созревания, либо с наличием большого количества криоповреждений в структуре митохондрий (мембранах) и хроматина. Следовательно, использование при витрификации женских гамет Bostaurus 0,001 % нВДК положительно влияет на показатели ядерно-цитоплазматического созревания ДВ ооцитов. Проблема создания криобанка ооцитов приобретает особую актуальность в условиях крупномасштабной селекции и снижения репродуктивных качеств высокопродуктивных коров для сохранения генофонда элитных особей и конструирования новых генотипов, в том числе с использованием технологии CRISPR-Cas9. Полученные нами данные могут быть использованы при совершенствовании этапов экстра- и интраовариальной витрификации ооцитов других видов животных, в том числе и при решении проблем бесплодия у человека.
Интенсивность флуоресценции (мкА) зонда MitoTracker Orange CMTMRos («Thermo Fisher Scientific», Великобритания) в нативных и девитрифицированных ооцитах голштинизированных коров на разных стадиях мейоза при воздействии наночастиц высокодисперсного кремнезема (нВДК) (М±SEM, число экспериментов — 5, общее число ооцитов — 529, культура in vitro) |
|||||
Группа |
Обработка |
Число |
Интенсивность флуоресценции MitoTracker Orange CMTMRos на соответствующей стадии мейоза |
||
диплотена |
метафаза I |
метафаза II |
|||
Нативные ооциты |
− |
133 |
331±16,2a |
188±11,3e |
143±10,1i |
+ |
139 |
309±15,6b |
203±11,8f |
149±14,7j |
|
Девитрифицированные ооциты |
− |
128 |
77±6,3c |
139±11,7g |
101±9,7k |
+ |
129 |
169±12,8d |
181±7,7h |
141±11,2l |
|
a:c; c:d; b:d; g:h; e:g; i:k; k:l; a:d; b:c; f:g; j:kРазличия в парах статистически значимы при p < 0,05 (t-критерий Стьюдента). |
|||||
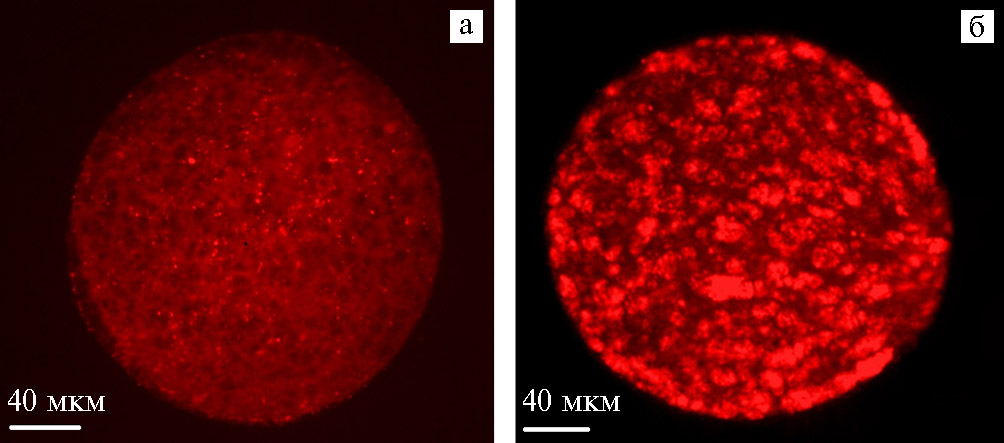
Рис. 1. Репрезентативное изображение митохондрий в девитрифицированных ооцитах голштинизированных коров на стадии диплотены с низкой (а) и высокой (б) интенсивностью флуоресценции зонда MitoTracker Orange MTMRos («Thermo Fisher Scientific», Великобритания): а — ооциты, не обработанные наночастицами высокодисперсного кремнезема (нВДК), б — ооциты, обработанные 0,001 % нВДК (культура in vitro; микроскоп ZEISS Axio Lab.A1, «Karl Zeiss», Германия).
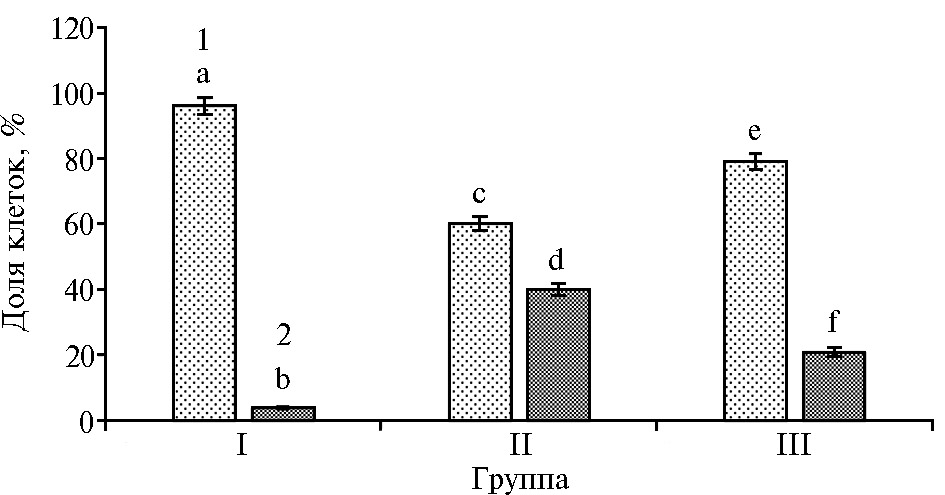
Рис. 2. Доля ооцитов голштинизированных коров с нормальным (1) и дегенерированным (2) хроматином на стадии диплотены при воздействии наночастиц высокодисперсного кремнезема (нВДК): I — нативные ооциты (контроль), II — девитрифицированные ооциты (контроль), III — девитрифицированные ооциты, обработанные 0,001 % нВДК. Вертикальные отрезки — стандартные ошибки средних значений (±SEM) (0 ч культивирования, число ооцит
Различия в парах статистически значимы: для a:b; a:c; e:f; b:d; b:f; a:d; a:f; c:b; c:f; e:b; d:e — при p < 0,001; для c:d; c:e; d:f — при p < 0,01 (критерий χ2 Пирсона).
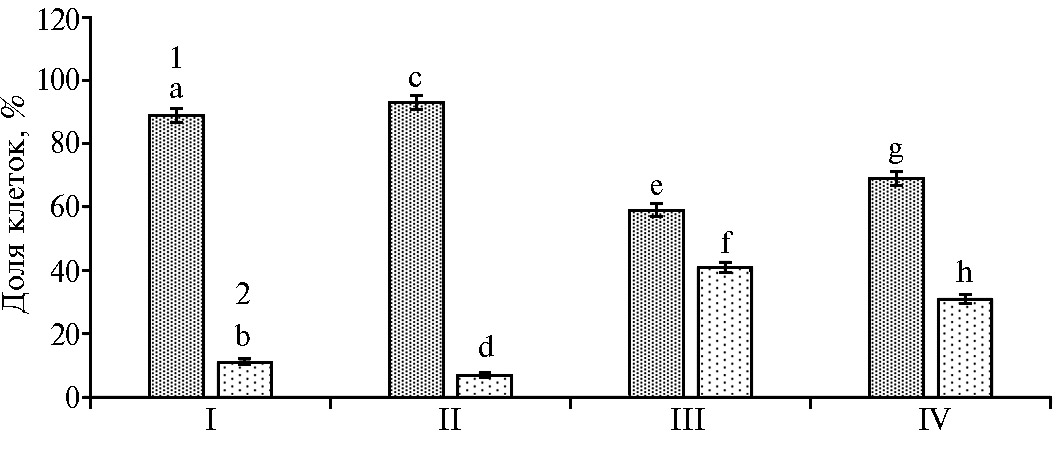
Рис. 3. Доля ооцитов голштинизированных коров с нормальным (1) и дегенерированным (2) хроматином на стадиях метафазы I (А) и метафазы II (Б) при культивировании с наночастицами высокодисперсного кремнезема (нВДК): I — нативные ооциты (контроль), II — нативные ооциты, обработанные 0,001 % нВДК, III — девитрифицированные ооциты (контроль), IV — девитрифицированные ооциты, обработанные 0,001 % нВДК. Вертикальные отрезки — стандартные ошибки средних значений (±SEM) (24 ч культивирования, число ооцитов — 419, число экспериментов — 4; культура in vitro).
А: a:b; c:d; g:h; a:e; a:g; c:e; c:g; b:f; b:h; d:f; d:h; a:d; a:f; a:h; c:b; c:f; c:h; e:b; e:d; e:h; g:b; g:d; g:f — различия в парах статистически значимы при p < 0,001; различия e:f статистически значимы при p < 0,01.
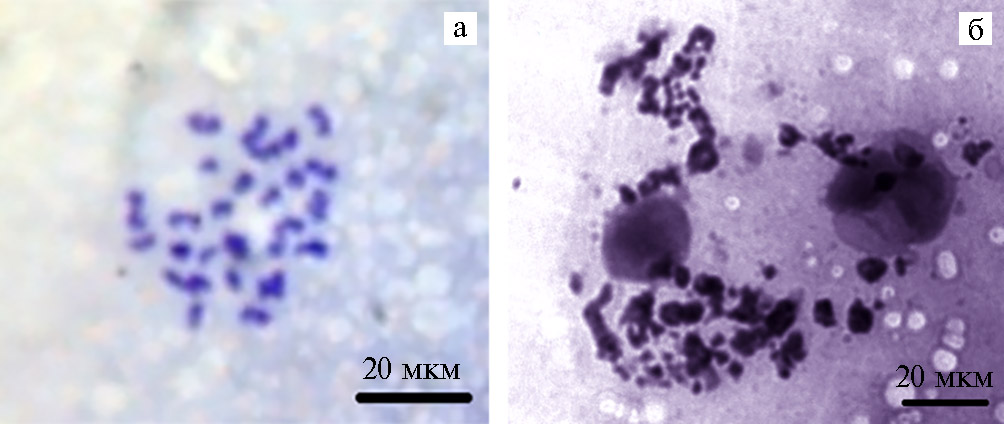
Рис. 4. Репрезентативное изображение нормального (а) и дегенерированного (б) хроматина на стадии метафазы II в девитрифицированных ооцитах голштинизированных коров при обработке наночастицами высокодисперсного кремнезема (0,001 %) ( культура in vitro; цитогенетический препарат, окрашивание азур-эозином по Романовскому-Гимзе; микроскоп ZEISS Axio Lab.A1, «Karl Zeiss», Германия).
ЛИТЕРАТУРА
- Xu H.F., Hao B.T., Liu L.J., Tang L.L., Liu B.L. Calorimetric studies on thermal properties of nano-cryoprotectant solutions during vitrification. CryoLetters, 2016, 37(6): 406-410.
- Li W., Zhou X., Dai J., Zhang D., Liu B., Wang H., Xu L. Effect of hydroxyapatite nanoparticles on MII-stage porcine oocytes vitrification and the study of its mechanism. Biochemical Engineering Journal, 2013, 30(4): 789-793.
- Туров В.В., Барвинченко В.Н., Крупская Т.В., Гунько В.М., Чехун В.Ф. Гидратные свойства композитного материала на основе высокодисперсного кремнезема и ДНК. Бioтехнологiя, 2011, 4(4): 34-48.
- Rubio L., Pyrgiotakis G., Beltran-Huarac J., Zhang Y., Gaurav J., Deloid G., Spyrogianni A., Sarosiek K., Bello D., Demokritou P. Safer-by-design flame-sprayed silicon dioxide nanoparticles: the role of silanol content on ROS generation, surface activity and cytotoxicity. Particle and Fibre Toxicology, 2019, 16(1): 40 (doi: 10.1186/s12989-019-0325-1).
- Галаган Н.П., Гунько В.М., Порхун Н.Г., Новикова Е.А., Туров В.В. Влияние дисперсности нанокремнеземов на их биоактивность по отношению к гаметам быка. Доповiдi Нацiональної академiї наук України, 2012, 5: 126-133.
- Савченко Д.С. Изучение антиоксидантных свойств нанокомпозита высокодисперсного кремнезема с наночастицами серебра. Медицина и образование в Сибири, 2013, 6: 23-30.
- Al-Zubaidi U., Liu J., Cinar O., Robker R.L., Adhikari D., Carroll J. The spatio-temporal dynamics of mitochondrial membrane potential during oocyte maturation. Molecular Human Reproduction, 2019, 25(11): 695-705 (doi: 10.1093/molehr/gaz055).
- Пожилова Е.В., Новиков В.Г., Левченкова О.С. Регуляторная роль митохондриальной поры и возможности ее фармакологической модуляции. Обзоры по клинической фармакологии и лекарственной терапии, 2014, 12(3): 13-19.
- Yoon S.Y., Eum J.H., Cha S.K., Yoon T.K., Lee D.R., Lee W.S. Prematuration culture with phosphodiesterase inhibitors after vitrification may induce recovery of mitochondrial activity in vitrified mouse immature oocytes. Biopreservation and Biobanking, 2018, 16(4): 296-303 (doi: 10.1089/bio.2018.0010).
- Galagan N.P., Klymenko N.Y., Orel I.L., Novikova E.A., Turov V.V. Biofunctional nanomaterials based on ultra-fine silica, protein and aminocarbohydrates. Biopolymers and Cell, 2010, 26(3): 205-213 (doi: 10.7124/bc.000158).
- Chistyakova I.V., Kuzmina T.I., Stanislavovich T.I., Kovtun S.V. Effects of highly dispersed silica nanoparticles on the cryoresistance of the bovine cumulus-oocyte complexes. Cryobiology, 2018, 85: 176 (doi: 10.1016/j.cryobiol.2018.10.215).
- Кузьмина Т.И., Чистякова И.В. Влияние наночастиц высокодисперсного кремнезема на апоптоз в нативных и девитрифицированных клетках гранулезы Bos taurus. Актуальные вопросы ветеринарной биологии, 2019, 3(43): 8-12 (doi: 10.24411/2074-5036-2019-10031).
- Кузьмина Т.И., Станиславович Т.И., Молчанов А.В. Влияние наночастиц высокодисперсного кренезема на показатели криорезистентности девитрифицированных ооцит-кумулюсных комплексов Bos taurus. Аграрный научный журнал, 2019, 3: 29-34 (doi: 10.28983/asj.y2019i3pp29-34).
- Новодережкина Е.А., Животовский Б.Д., Гогвадзе В.Г. Индукция неспецифической проницаемости митохондриальной мембраны и ее роль в гибели клеток. Молекулярная биология, 2016, 50(1): 51-68 (doi: 10.7868/S002689841601016X).
- Kim B., Yoon S.-Y., Cha S.K., Kwak K.H., Fissore R.A., Parys J.B., Yoon T.K., Lee D.R. Alterations in calcium oscillatory activity in vitrified mouse eggs impact on egg quality and subsequent embryonic development. Pflügers Archiv — European Journal of Physiology, 2011, 461: 515-526 (doi: 10.1007/s00424-011-0955-0).
- Mostek A., Słowińska M., Judycka S., Karol H., Ciereszko A., Dietrich M.A. Identification of oxidatively modified proteins due to cryopreservation of carp semen. Journal of Animal Science, 2018, 96(4): 1453-1465 (doi: 10.1093/jas/sky063).
- Xiao L., Zhao D., Chan W.-H., Choi M.M.F., Li H.-W. Inhibition of beta 1-40 amyloid fibrillation with N-acetyl-L-cysteine capped quantum dots. Biomaterials, 2010, 31(1): 91-98 (doi: 10.1016/j.biomaterials.2009.09.014).
- Shin W.S., Di J., Cao Q., Li B., Seidler P.M., Murray K.A., Bitan G., Jiang L. Amyloid β-protein oligomers promote the uptake of tau fibril seeds potentiating intracellular tau aggregation. Alzheimers Research and Therapy, 2019, 11(1): 86 (doi: 10.1186/s13195-019-0541-9).
- Sukhanova A., Poly S., Bozrova S., Lambert E., Ewald M., Karaulov A., Molinari M., Nabiev I. Nanoparticles with a specific size and surface charge promote disruption of the secondary structure and amyloid-like fibrillation of human insulin under physiological conditions. Frontiers in Chemistry, 2019, 7: 480-486 (doi: 10.3389/fchem.2019.00480).
- Desai J., Foresto-Neto O., Honarpisheh M., Steiger S., Nakazawa D., Popper B., Buhl E.M., Boor P., Mulay S.R., Anders H.-J. Particles of different sizes and shapes induce neutrophil necroptosis followed by the release of neutrophil extracellular trap-like chromatin. Scientific Reports, 2017, 7(1): 15003 (doi: 10.1038/s41598-017-15106-0).
- Sun L., Li Y., Liu X., Jin M., Zhang L., Du Z., Guo C., Huang P., Sun Z. Cytotoxicity and mitochondrial damage caused by silica nanoparticles. Toxicology in Vitro, 2011, 25(8): 1619-1629 (doi: 10.1016/j.tiv.2011.06.012).
- Barros V.R.P., Monte A.P.O., Santos J.M.S., Lins T.L.B.G., Cavalcante A.Y.P, Gouveia B.B., Müller M.C., Oliveira Junior J.L., Barberino R.S., Donfack N.J., Araújo V.R., Matos M.H.T. Effects of melatonin on the in vitro growth of early antral follicles and maturation of ovine oocytes. Domestic Animal Endocrinology, 2019, 71: 106386 (doi: 10.1016/j.domaniend.2019.106386).
- Савченко Д.С. Изучение генотоксичности и цитотоксичности нанокомпозита высокодисперсного кремнезема с наночастицами серебра. Вестник новых медицинских технологий, 2013, 20(4): 44-47.
- Yang Y., Du X., Wang Q., Liu J., Zhang E., Sai L., Peng C., Lavin M.F., Yeo A.J., Yang X., Shao H., Du Z. Mechanism of cell death induced by silica nanoparticles in hepatocyte cells is by apoptosis. International Journal of Molecular Medicine, 2019, 44(3): 903-912 (doi: 10.3892/ijmm.2019.4265).
- Murugadoss S ., Lison D., Godderis L ., Van Den Brule S., Mast J., Brassinne F., Sebaihi N., Hoet P.H. Toxicology of silica nanoparticles: an update. Archives of Toxicology, 2017, 91(9): 2967-3010 (doi: 10.1007/s00204-017-1993-y).
- Li Y., Duan J., Chai X., Yang M., Wang J., Chen R., Sun Z. Microarray-assisted size-effect study of amorphous silica nanoparticles on human bronchial epithelial cells. Nanoscale, 2019, 11(47): 22907-22923 (doi: 10.1039/c9nr07350g).