doi: 10.15389/agrobiology.2019.4.723rus
УДК 636.5:591.463.12:591.8.086.2
Работа выполнена при финансовой поддержке РНФ, грант № 16-16-04104.
ДИНАМИКА ГИСТОЛОГИЧЕСКОЙ СТРУКТУРЫ СЕМЕННИКОВ У ПЕРЕПЕЛОВ (Coturnix coturnix) В ПРОЦЕССЕ СПЕРМАТОГЕНЕЗА
И.П. НОВГОРОДОВА, Н.А. ВОЛКОВА, А.Н. ВЕТОХ, Л.А. ВОЛКОВА, В.А. БАГИРОВ, Н.А. ЗИНОВЬЕВА
Половые клетки самцов — уникальный объект для генетических и физиологических исследований как биологической основы современного животноводства. Процессы созревания и дифференцировки половых клеток самцов животных и птиц представляют огромный интерес для сравнительной эмбриологии, биологии развития, медицины и биотехнологии. Перспективный вид для таких экспериментальных работ — перепела, которые характеризуются ранней половой зрелостью и коротким периодом генерации. Наиболее интересны стволовые клетки семенников — сперматогонии, которые в настоящее время активно изучаются в связи с тем, что их рассматривают в качестве перспективных мишеней для введения рекомбинантной ДНК с целью получения трансгенных особей. Однако морфология половых клеток самцов домашних птиц в процессе их формирования освещена неполно. В этом исследований мы впервые подробно описываем гистологические особенности ткани сперматогенного эпителия перепелов, находящихся на разных стадиях сперматогенеза и динамику популяций сперматогеных клеток семенников. Целью работы было выявление возрастных особенностей сперматогенеза, связанных с динамикой развития разных типов клеток эпителиосперматогенного слоя семенных канальцев у перепелов. Для этого у перепелов (Coturnix coturnix) эстонской породы исследовали гистологическую структура семенников в возрасте 1, 2, 3, 4, 5, 6, 12 и 24 нед (по 10 самцов в каждой возрастной группе). Ткань семенников фиксировали в растворе Буэна, дегидратировали в спиртах возрастающей концентрации и заливали в парафин. Готовили гистологические срезы толщиной 5-6 мкм и окрашивали гематоксилином и эозином. Определяли состав сперматогенных клеток и их соотношение в семенных канальцах. Просматривали не менее 30 семенных канальцев каждого самца. Показано, что диаметр семенных канальцев семенников изменялся в процессе онтогенеза и составил в возрасте 1, 2, 3, 4, 5, 6, 12 и 24 нед соответственно 42±1, 71±2, 91±2, 117±2, 237±4, 278±5, 282±7 и 291±6 мкм. Популяция клеток внутри семенных канальцев была представлена клетками Сертоли и генеративными клетками (сперматогонии, сперматоциты, сперматиды и спермии) на разных стадиях дифференцировки в зависимости от возраста. Их общее число в семенных канальцах достоверно увеличивалось с возрастом (соответственно 18±1, 24±1, 58±4, 80±6, 249±16, 587±34, 658±24 и 540±41 в возрасте 1, 2, 3, 4, 5, 6, 12 и 24 нед; p < 0,01). В возрасте 1 нед в семенных канальцах доминировали клетки Сертоли (среднее число в одном семенном канальце 12,0±0,9), в то время как сперматогонии были представлены единичными клетками (от 1 до 4). Максимальный процент сперматогониев от общего числа сперматогенных клеток (76±2 %) отмечали в возрасте 3 нед. У 4-недельных самцов в семенных канальцах визуализировались сперматоциты 1-го и 2-го порядка, а с 5-недельного возраста — сперматиды. В возрасте 5 нед в семенных канальцах перепелов выявлялись единичные спермии, число которых увеличивалось к 6 нед. Таким образом, у перепелов возраст от 1 до 3 нед — это оптимальный период для манипуляций со сперматогониями при их использовании в качестве биологического материала для сохранения генетических ресурсов сельскохозяйственной птицы в условиях криобанков, а также как мишеней для введения рекомбинантной ДНК с целью генетической модификации половых клеток самцов и последующего получения трансгенного потомства.
Ключевые слова: перепел, семенники, клетки Сертоли, сперматогенез, сперматогонии.
Изучение процессов созревания и дифференцировки половых клеток самцов животных и птиц представляет огромный интерес для сравнительной эмбриологии, биологии развития, медицины и биотехнологии в целом. В особенности это касается сперматогониев как предшественников зрелых половых клеток — спермиев. Сперматогонии рассматриваются в качестве перспективных мишеней для введения рекомбинантной ДНК с целью получения трансгенных особей (1). Такая технология предусматривает выделение и трансформацию in vitro сперматогониев с их последующей трансплантацией в семенники самцов-реципиентов, у которых предварительно блокируют собственный сперматогенез (2, 3). Трансплантированные сперматогонии впоследствии дифференцируются в спермии, которые используют для получения трансгенного потомства. Подобная возможность получения химерных и генетически модифицированных особей показана в ряде работ на лабораторных животных (4, 5), свиньях (6, 7), овцах (8), а также на петухах (9, 10). Клетки гонад самцов также служат ценным генетическим материалом для создания криобанков в рамках сохранения и поддержания генофонда ценных пород животных и птицы (11, 12).
В отличие от млекопитающих, у птиц, в том числе перепелов, семенники остаются в брюшной полости (на месте их развития) в течение всей жизни (13, 14). Особенности половых органов кур, индеек, уток и перепелов достаточно глубоко описаны с точки зрения анатомии, в то время как морфология клеток самцов домашней птицы освещена неполно (15).
У птиц, как и у млекопитающих, сперматогенез представляет собой длительный процесс постепенной трансформации зародышевых клеток в сперматозоиды в пределах границ семенных канальцев семенника и включает три последовательных этапа — стадии сперматоцитогенеза, сперматидогенеза и спермиогенеза. Физиологические особенности и анатомические характеристики, связанные со сперматогенезом у птиц, составляют предмет обширных исследований (16, 17). При сперматогенезе происходит клеточная пролиферация с повторными митотическими делениями, дублированием хромосом, мейотическим делением клеток и т.д. для дальнейшего образования гаплоидных сперматид с их последующей дифференциацией в зрелые спермии (18).
В представленной работе мы впервые подробно изучили гистологические особенности ткани сперматогенного эпителия перепелов на разных стадиях сперматогенеза и охарактеризовали популяции сперматогеных клеток семенников в динамике.
Нашей целью было изучение возрастных особенностей сперматогенеза у перепелов в связи с динамикой развития разных типов клеток эпителио-сперматогенного слоя семенных канальцев.
Методика. В экспериментах использовали самцов перепелов (Coturnix coturnix) эстонской породы в возрасте 1, 2, 3, 4, 5, 6, 12 и 24 нед (группы по восьми возрастным категориям, в каждой группе по 10 гол.). Биоматериалом служили семенники, полученные при убое.
Отобранные ткани семенников фиксировали в течение 48 ч в растворе Буэна, состоящем их пикриновой, уксусной кислот и формалина в соотношении 15:1:5, после чего образцы заливали в парафин и готовили гистологические срезы толщиной 5-6 мкм (12, 13). Препараты окрашивали гематоксилином и эозином («BioVitrum», Россия).
При проведении гистологического анализа просматривали семенные канальцы, имеющие округлую форму и просвет (поперечный срез). Типы клеток сперматогенного эпителия идентифицировали по морфологическим характеристикам (14, 15). Гистологические препараты изучали методом световой микроскопии (Ni-U, «Nikon», Япония; микроскоп оснащен пакетом программ для обработки и анализ изображений NIS-Elements, «Nikon», Япония). Оценивали диаметр семенных канальцев, число и состав находящихся в них сперматогенных клеток.
Статистическую обработку данных проводили с использованием пакета для анализа данных MS Excel 2016 (t-test). В таблицах представлены средние арифметические значения (M) и ошибки средних (±SEM). Различия считали статистически значимыми при р < 0,01.
Результаты. Гистологическая структура семенников у перепелов была аналогична таковой у млекопитающих. Паренхиматозную ткань органа формировала система извитых семенных канальцев, содержащих разные типы клеток (клетки Сертоли, сперматогонии, сперматоциты, сперматиды и спермии).
Размер семенных канальцев у перепелов изменялся в течение онтогенеза (табл. 1). В ранний период постнатального развития этот показатель с возрастом незначительно повышался. В период от 1 до 2 нед диаметр семенных канальцев возрастал на 69 % (p < 0,01), со 2-й по 3-ю нед — на 28 % (p < 0,01), с 3-й по 4-ю нед — на 29 % (p < 0,01). Значительный рост размеров семенных канальцев отмечали с 4-й до 5-й нед. Диаметр и площадь семенных канальцев у 5-недельных перепелов были в 2 раза больше аналогичных показателей, установленных для особей в возрасте 4 нед. С 6-й до 24-й нед размеры семенных канальцев практически не изменялись.
Увеличение диаметра и площади семенных канальцев у перепелов с возрастом было обусловлено ростом и дифференцировкой сперматогенных клеток (табл. 1). У 1-недельных самцов число сперматогенных клеток в одном семенном канальце варьировало от 11 до 26 и составило в среднем 18±1. На 2-й нед этот показатель возрастал по отношению к значениям, установленным у 1-недельных особей, на 33 % (р < 0,01), в период с 2 до 3 нед — на 142 % (p < 0,01), с 3 до 4 нед — на 86 % (p < 0,01). Значительный рост и дифференцировка сперматогенных клеток отмечались с 4-й до 6-й нед. В семенных канальцах 5-недельных самцов по сравнению с 4-недельными число сперматогенных клеток увеличивалось в 3,1 раза, с 5-й до 6-й нед — 2-кратно. После достижения половозрелости (6 нед) рост и развитие самцов практически не сопровождались изменениями в числе сперматогенных клеток в семенных канальцах семенников. Различия в этом показателе, установленном на 6-й нед и в более позднем возрасте (3 и 6 мес), не превышали 1,8 %.
Наличие, число и соотношение сперматогенных клеток внутри семенных канальцев семенников варьировали в зависимости от возраста перепелов (табл. 2). У 1-недельных перепелов базальную мембрану семенных канальцев семенников выстилали клетки Сертоли и единичные сперматогонии (рис. 1, А), при этом по числу клетки Сертоли превалировали с процентным соотношением соответственно 71 % против 29 %. Клетки Сертоли имели темноокрашенное ядро пирамидальной формы, расположенное на базальной мембране. Сперматогонии располагались по семенному канальцу и не встречались на базальной мембране, были представлены преимущественно типом А (стволовые клетки семенника). Клетки этого типа были крупными и характеризовались наличием ядра эллиптической или округлой формы, расположенного, как правило, на базальной мембране канальца. Ядерный хроматин в ядре концентрировался в одной зоне.
У 2-недельных перепелов клетки сперматогенного эпителия семенных канальцев также были представлены двумя типами — клетками Сертоли и сперматогониями. Число сперматогониев разных типов в семенном канальце увеличивалось до 10±1 (р < 0,01). При этом число клеток Сертоли изменялось незначительно (см. табл. 2). Сперматогонии встречались как на периферии, так и внутри семенного канальца. Наряду со сперматогониями типа А выявлялись сперматогонии промежуточного типа и типа В. Сперматогонии промежуточного типа были чуть меньше сперматогониев типа А, хроматин в их ядре сливался в одно или два ядрышка и имел более темный цвет. Сперматогонии типа В характеризовались наличием большого ядра круглой или эллиптической формы, хлопья хроматина в ядре распределялись по всей эндоплазме. На указанной стадии развития просвет семенного канальца отсутствовал.
Формирование в семенных канальцах просвета, необходимого для выхода спермиев у взрослых особей, отмечали в возрасте 3 нед (см. рис. 1, Б). Сперматогонии располагались на базальной мембране. Их число по сравнению с предыдущим периодом увеличилось в 4,4 раза (р < 0,01).
В возрасте 4 нед небольшие формирующиеся просветы имелись во всех семенных канальцах. В некоторых из них клетки сперматогенного эпителия выстраивались в 5-6 рядов. Наряду с клетками Сертоли и сперматогониями выявлялись сперматоциты 1-го и 2-го порядка, которые располагались ближе к центру канальца. Сперматоциты 1-го порядка представляли собой большие клетки с крупным овальным ядром. Сперматоциты 2-го порядка были меньше по размеру (см. рис. 1, В). Среднее число сперматоцитов 1-го и 2-го порядка в семенном канальце не превышало соответственно 11±1 и 5±1. Число клеток Сертоли и сперматогониев в этом возрасте по сравнению с предыдущим периодом заметно не изменялось: различия по показателям не превышали соответственно 6 и 13 %.
У 5-недельных особей популяция сперматогенных клеток в семенных канальцах была представлена клетками Сертоли, сперматогониями, находящимися на базальной мембране, сперматоцитами 1-го и 2-го порядка (см. табл. 2). Вблизи просвета отмечали наличие сперматид в виде незрелых мелких образований округлой формы с отчетливо видным ядром. В просвете некоторых семенных канальцев появлялись единичные сперматозоиды.
К 6-недельному возрасту ближе к просвету семенного канальца выявлялись сперматиды удлиненной формы с хвостиками, их число по сравнению с предыдущим периодом увеличивалось в 2,0 раза (р < 0,01, см. табл. 2, рис. 1, Г). Отмечалось наличие сперматид на разных этапах спермиогенеза, что обусловило их неодинаковую форму. Так, с приближением к просвету семенных канальцев сперматиды уменьшались в размере и представляли собой клетки с овальной удлиненной го-ловкой темного цвета и длинными хвостиками. В единичных семенных ка-нальцах встречались спе-рматиды, располагающиеся пучками. В просветах большинства семенных канальцев выявлялись сформировавшиеся сперматозоиды. Клетки Сертоли зачастую имели вытянутую форму (в виде тяжей, располагающихся практически от базальной мембраны до просвета канальца). Общее число клеток в семенных канальцах по сравнению с аналогичным показателем в возрасте 5 нед повышалось в 2,4 раза, преимущественно за счет увеличения числа сперматоцитов, сперматид и спермиев (р < 0,01).
У взрослых самцов в возрасте 3 и 6 мес в семенных канальцах присутствовали все клетки сперматогенного эпителия; средние значения этих показателей были практически одинаковыми. Различия по содержанию в семенных канальцах сперматогенных клеток разных типов, установленные у перепелов в 6-недельном возрасте и в 3 и 6 мес, не превышали 11 %.
Таким образом, проведенные исследования показали, что у разновозрастных перепелов число разных типов сперматогенных клеток и их соотношение в семенных канальцах семенников варьировали. Доля клеток Сертоли от общего числа сперматогенных клеток в семенном канальце с возрастом снижалась за счет относительного содержания сперматоцитов и сперматид (рис. 2). В ранний период постнатального развития самцов доля сперматогониев повышалась по мере взросления особи, достигая максимального значения (до 76,6 %) к 3-недельному возрасту. В последующий период этот показатель снижался до 13 % в возрасте 6 мес.
Результаты настоящего исследования согласуются с данными, полученными нами ранее при изучении возрастных особенностей сперматогенеза у кроликов (19), петухов (20), цесарей (21). У самцов перечисленных видов показано изменение состава сперматогенных клеток в семенных канальцах семенников в онтогенезе. В течение постнатального периода отмечалась неравномерность в увеличении размеров семенных канальцев, разница по наличию, числу и соотношению сперматогенных клеток в них. В ранний период онтогенеза размеры семенных канальцев и число сперматогенных клеток в них постоянно и существенно возрастали. Интенсивный рост и дифференцировка сперматогенных клеток и, как следствие, увеличение размеров семенных канальцев выявлено в период, предшествующий достижению половозрелости: у самцов кроликов в возрасте от 5 до 6 мес, у петухов и цесарей — от 4 до 5 мес. Отметим, что подобных исследований, выполненных другими авторами, нам обнаружить не удалось.
Среди доступных информационных источников мы также не нашли публикаций, касающихся возрастных изменений морфометрических характеристик семенных канальцев семенников у перепелов и количественного состава сперматогенных клеток. Работы на птице, в том числе перепелах, были связаны преимущественно с изучением морфометрических показателей (диаметр, площадь и др.) сперматогенных клеток и их структурных единиц в процессе дифференцировки (22, 23). Имеется ряд работ других авторов по изучению анатомического строения и морфометрических данных половых органов у самцов перепелов (24-27). Так, была выполнено оценка морфометрических показателей семенников у 60-суточных перепелов, в частности определены размеры семенников, их анатомическое строение и гистологическая структура (24). Представленные в указанной работе результаты гистологических исследований согласуются с полученными нами. T.A. Kannan с соавт. (25) изучали анатомические и морфологические особенности семенников перепелов в возрастной динамике: у молодых перепелов, по достижении половозрелости и у взрослых самцов. Было показано увеличение размеров семенников до 22-недельного возраста. Аналогичные данные получены К.В. Баусовой (26). В ряде работ отмечается влияние на сперматогенез перепелов различных кормов, гормонов, гербицидов и других веществ (28-30), а также естественного и искусственного освещения (31).
Итак, у перепелов (Coturnix coturnix) размер семенных канальцев, число и состав сперматогенных клеток в них варьирует в зависимости от возраста. В возрасте 1-3 нед популяция клеток эпителио-сперматогенного слоя семенных канальцев представлена двумя типами — клетками Сертоли и сперматогониями. У 3-недельных особей начинает формироваться семенной просвет, с 4-й нед на гистологических срезах семенников выявляются сперматоциты 1-го и 2-го порядка, а с 5-й нед — сперматиды. В семенных канальцах 6-недельных перепелов присутствуют все типы клеток эпителио-сперматогенного слоя: клетки Сертоли, сперматогонии разных типов, сперматоциты 1-го и 2-го порядка, сперматиды, спермии. Эти данные расширяют представления о морфологии формирующихся половых клеток у самцов домашней птицы и позволяют заключить, что у перепелов возраст от 1 до 3 нед — это оптимальный период для манипуляций со сперматогониями при их использовании в качестве биологического материала для сохранения генетических ресурсов сельскохозяйственной птицы в условиях криобанков, а также как мишеней для введения генных конструкций с целью получения трансгенной птицы.
1. Возрастная динамика морфологических показателей, характеризующих гистоструктуру семенника у перепелов (Coturnix coturnix) эстонской породы (n = 80, М±SEM) |
|||
Возраст, нед |
Диаметр семенных |
Площадь семенных |
Число сперматогенных клеток |
1 |
42±1 |
1329±55 |
18±1 |
2 |
71±2ab |
4117±222ab |
24±1ab |
3 |
91±2ab |
6172±177ab |
58±4ab |
4 |
117±2ab |
9932±310ab |
80±6ab |
5 |
237±4ab |
40688±993ab |
249±16ab |
6 |
278±5ab |
56232±868ab |
587±34ab |
12 |
282±7ac |
57023±766ac |
598±23ac |
24 |
291±6ac |
55985±812ac |
570±41ac |
a, b Различия по отношению к предыдущей возрастной группе статистически значимы при p < 0,01. |
|||
2. Состав популяции клеток сперматогенного эпителия в семенных |
|||||
Возраст |
Тип клеток |
||||
клетки Сертоли |
сперматогонии |
сперматоциты |
сперматоциты |
сперматиды |
|
1 |
12±1 |
5±1 |
0 |
0 |
0 |
2 |
13±1 |
10±1ab |
0 |
0 |
0 |
3 |
15±1 |
44±4ab |
0 |
− |
0 |
4 |
14±1 |
49±3 |
11±1 |
5±1 |
0 |
5 |
16±1 |
92±5ab |
21±3ab |
25±3ab |
96±13 |
6 |
16±1 |
112±4ab |
61±5ab |
84±2ab |
192±3ab |
3 |
18±3 |
113±9 |
65±5 |
95±8 |
194±5 |
6 |
18±2 |
119±6 |
63±4 |
93±4 |
192±3 |
12 |
12±1 |
5±1 |
0 |
0 |
0 |
24 |
13±1 |
10±1ab |
0 |
0 |
0 |
a, b Различия по отношению к предыдущей возрастной группе статистически значимы при p < 0,01. |
|||||
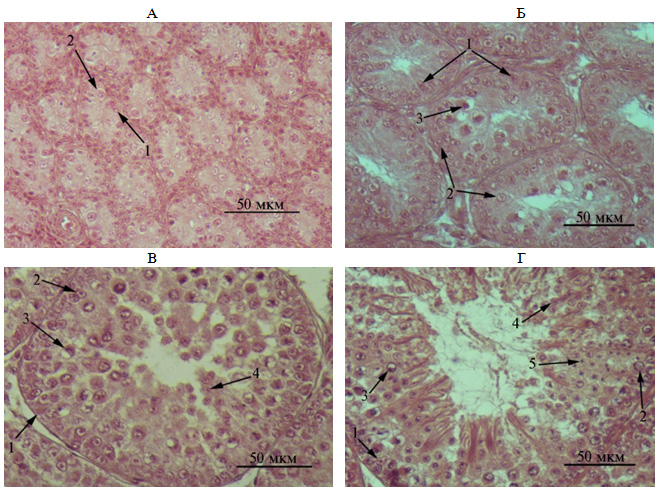
Рис. 1. Гистологическая структура семенных канальцев семенников перепелов (Coturnix coturnix) эстонской породы в возрасте 1 нед (А), 3 нед (Б), 4 нед (В) и 6 нед (Г): 1 — клетки Сертоли, 2 — сперматогонии, 3 — сперматоциты 1-го порядка, 4 — сперматоциты 2-го порядка, 5 — сперматиды. Окрашивание гематоксилином и эозином, световая микроскопия (Ni-U, «Nikon», Япония), увеличение ×400.
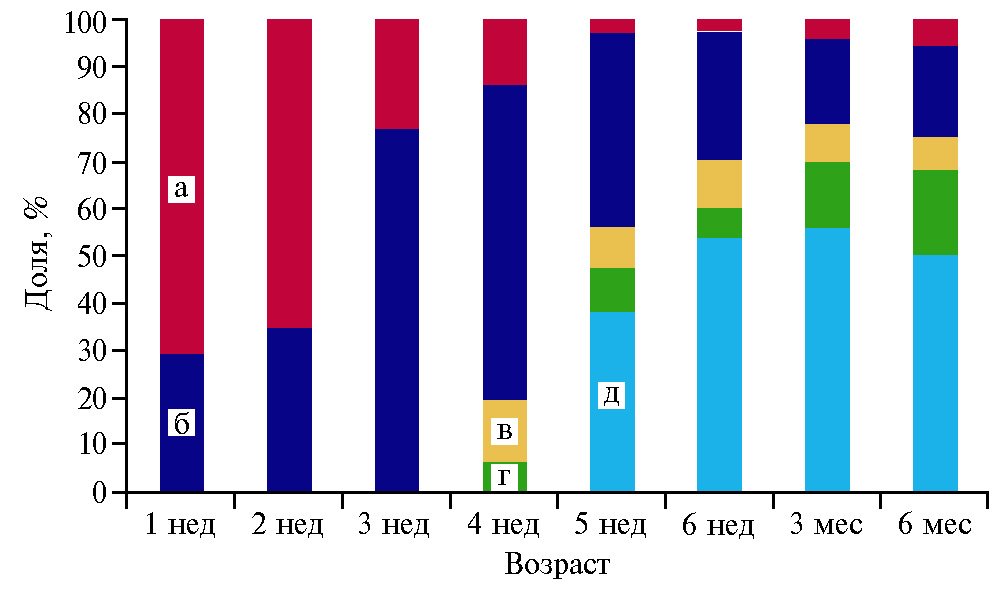
Рис. 2. Соотношение клеток сперматогенного эпителия семенных канальцев в семенниках разновозрастных перепелов (Coturnix coturnix) эстонской породы: а — клетки Сертоли, б — сперматогонии, в — сперматоциты 1-го порядка, г — спрматоциты 2-го порядка, д — сперматиды.
ЛИТЕРАТУРА
- Olive V., Cuzin F. The spermatogonial stem cell: from basic knowledge to transgenic technology. The International Journal of Biochemistry & Cell Biology, 2005, 37: 246-250 (doi: 10.1016/j.biocel.2004.07.017).
- Takashima S. Biology and manipulation technologies of male germline stem cells in mammals. Reproductive Medicine and Biology, 2018, 17(4): 398-406 (doi: 10.1002/rmb2.12220).
- Sofikitis N., Kaponis A., Mio Y., Makredimas D., Giannakis D., Yamamoto Y., Kanakas N., Kawamura H., Georgiou J., Schrader M., Lolis E., Giannakopoulos X., Loutradis D., Tarlatzis V., Miyagawa I. Germ cell transplantation: a review and progress report on ICSI from spermatozoa generated in xenogeneic testes. Human Reproduction Update, 2003, 9(3): 291-307 (doi: 10.1093/humupd/dmg015).
- Dobrinski I., Avarbock M.R., Brinster R.L. Germ cell transplantation from large domestic animals into mouse testes. Molecular Reproduction and Development, 2000, 57: 270-279 (doi: 10.1095/biolreprod66.1.21).
- Brinster R.L. Germline stem cell transplantation and transgenesis. Science, 2002, 296: 2174-2176 (doi: 10.1126/science.1071607).
- Honaramooz A.1., Megee S.O., Dobrinski I. Germ cell transplantation in pigs. Biology of Reproduction, 2002, 66(1): 21-28 (doi: 10.1095/biolreprod66.1.21).
- Savchenkova I.P., Korjikova S.V., Kostereva N.V., Ernst L.K. Cultivation and transfer of porcine type A spermatogonia. Russian Journal of Developmental Biology, 2006, 37(4): 242-249 (doi: 10.1134/S1062360406040060).
- Rodriguez-Sosa J.R., Dobson H., Hahnel A. Isolation and transplantation of spermatogonia in sheep. Theriogenology, 2006, 66: 2091-2103 (doi: 10.1016/j.theriogenology.2006.03.039).
- Yu F., Ding L.J., Sun G.B., Sun P.X., He X.H., Ni L.G., Li B.C. Transgenic sperm produced by electrotransfection and allogeneic transplantation of chicken fetal spermatogonial stem cells. Molecular Reproduction and Development, 2010, 77: 340-347 (doi: 10.1002/mrd.21147).
- Li B., Sun G., Sun H., Xu Q., Gao B., Zhou G., Zhau W., Wu X., Bao W., Yu F., Wang K., Chen G. Efficient generation of transgenic chickens using the SSCs in vivo and ex vivo transfection. Science China Life Sciences, 2008, 51: 734-742 (doi: 10.1007/s11427-008-0100-2).
- Zheng Y., Zhang Y., Qu R., He Y., Tian X., Zeng W. Spermatogonial stem cells from domestic animals: progress and prospects. Reproduction, 2014, 147: 65-74 (doi: 10.1530/REP-13-0466).
- Nakamura Y., Usui F., Miyahara D., Mori T., Ono T., Takeda K., Nirasawa K., Kagami H., Tagami T. Efficient system for preservation and regeneration of genetic resources in chicken: concurrent storage of primordial germ cells and live animals from early embryos of a rare indigenous fowl (Gifujidori). Reproduction Fertility Devevelopment, 2010, 22: 1237-1246 (doi: 10.1071/RD10056).
- Deviche P., Hurley L.L., Fokidis H.B. Function, avian testicular structure and regulation. In: Hormones and Reproduction of Vertebrates /D. Norris, K. H. Lopez (eds.).Elsevier Inc., 2011: 27-70 (doi: 10.1016/B978-0-12-374929-1.10002-2).
- Wei L., Peng K.-M., Liu H., Song H., Wang Y., Tang L. Histological examination of testicular cell development and apoptosis in the ostrich chick. Turkish Journal of Veterinary and Animal Sciences, 2011, 35(1): 7-14 (doi: 10.3906/vet-0806-2).
- Thurston R.J., Korn N. Spermiogenesis in commercial poultry species: anatomy and control. Poultry Science, 2000, 79: 1650-1668 (doi: 10.1093/ps/79.11.1650).
- Aire T.A., Ozegbe P.C. The testicular capsule and peritubular tissue of birds: morphometry, histology, ultrastructure and immunohistochemistry. Journal of Anatomy, 2007, 210: 731-740 (doi: 10.1111/j.1469-7580.2007.00726.x).
- Jamieson B.G.M. Avian spermatozoa: structure and phylogeny. In: Reproductive Biology and Phylogeny of Birds /B.G.M. Jamieson (ed.). Science Publishers, Enfield, New Hampshire, U.S.A. Jersey, Plymouth, U.K., 2007: 349-511.
- Peruquetti R.L., Taboga S.R., Vilela de Azeredo-Oliveira M.T. Morphological changes of mammalian nucleoli during spermatogenesis and their possible role in the chromatoid body assembling. International Scholarly Research Notices. Cell Biology, 2012, 2012: Article ID 829854 (doi: 10.5402/2012/829854).
- Ендовицкая И.П., Зиновьева Н.А., Эрнст Л.К. Динамика сперматогенеза у кролика Oryctolagus сuniculus. Цитология, 2005, 47(1): 44-48.
- Белоглазова Е.В., Котова Т.О., Волкова Н.А., Волкова Л.А., Зиновьева Н.А., Эрнст Л.К. Возрастная динамика сперматогенеза у петухов в связи с оптимизацией сроков биоинженерных манипуляций. Сельскохозяйственная биология, 2011, 6: 60-64.
- Volkova N.A., Vetokh A.N., Novgorodova I.P., Dotsev A.V., Zinovieva N.A. The dynamics of spermatogenesis in guinea fowls. Reproduction, Fertility and Development, 2017, 30(1): 211-211 (doi: 10.1071/RDv30n1Ab143).
- Abdul-Rahman I., Obese F.Y., Robinson J.E. Spermatogenesis and cellular associations in the seminiferous epithelium of Guinea cock (Numida meleagris). Canadian Journal of Animal Science, 2017, 97(2): 241-249 (doi: 10.1139/cjas-2016-0068).
- Aire T.A. Spermatogenesis in birds. Spermatogenesis, 2014, 4(3): e959392 (doi: 10.4161/21565554.2014.959392).
- Савельева А.Ю. Микроскопическое строение половых желез домашнего японского перепела. Международный научно-исследовательский журнал, 2016, 4(46): 59-62 (doi: 10.18454/IRJ.2016.46.165).
- Kannan T.A., Ramesh G., Sivakumar M. Age related changes in the gross and histoarchitecture of testis in Japanese quails (Coturnix coturnix japonica). International Journal of Livestock Research, 2015, 5(6): 26-33 (doi: 10.5455/ijlr.20150614083747).
- Баусова К.В. Постынкубационный онтогенез семенников эстонского перепела. Птицеводство, 2016, 8: 48-52.
- Kouatcho F.D., Kenfack A., Ngoula F., Teguia A. Sexual maturity prediction based on hormonal profiles, testes and semen characteristics in male Coturnix quail (Garsault, 1764) in the Western Highlands of Cameroon. International Journal of Agronomy and Agricultural Research (IJAAR), 2015, 7(4): 143-154.
- Bello U.M., Madekurozwa M.-C., Groenewald H.B., Aire T.A., Arukwe A. The effects on steroidogenesis and histopathology of adult male Japanese quails (Coturnix coturnix japonica) testis following pre-pubertal exposure to di(n-butyl) phthalate (DBP). Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 2014, 166: 24-33 (doi: 10.1016/j.cbpc.2014.06.005).
- Hussain R., Khan A., Mahmood F., Rehan S., Ali F. Clinico-hematological and tissue changes induced by butachlor in male Japanese quail (Coturnix japonica). Pesticide Biochemistry and Physiology, 2014, 109: 58-63 (doi: 10.1016/j.pestbp.2014.01.005).
- Türk G., Simsek U.G., Çeribasi A.O., Çeribasi S., Kaya S.O., Güvenç M., Çiftçi M., Sönmez M., Yüce A., Bayrakdar A., Yaman M., Тonbak F. Effect of cinnamon (Cinnamomum zeylanicum) bark oil on heat stress-induced changes in sperm production, testicular lipid peroxidation, testicular apoptosis, and androgenic receptor density in developing Japanese quails. Theriogenology, 2015, 84: 365-376 (doi: 10.1016/j.theriogenology.2015.03.035).
- Yadav S., Chaturved C.M. Light colour and intensity alters reproductive/seasonal responses in Japanese quail. Physiology & Behavior, 2015, 147: 163-168 (doi: 10.1016/j.physbeh.2015.04.036).