doi: 10.15389/agrobiology.2019.4.744rus
УДК 636.294:579.62:577.2
Исследование выполнено при поддержке гранта РНФ для реализации научного проекта № 17-76-20026 «Микробиоценоз рубца Rangifer tarandus Арктических регионов России как фундаментальная основа получения перспективных биотехнологий для сельскохозяйственных животных».
МИКРОБИОТА РУБЦА У СЕВЕРНЫХ ОЛЕНЕЙ (Rangifer tarandus) С КЛИНИЧЕСКИМИ ПРОЯВЛЕНИЯМИ НЕКРОБАКТЕРИОЗОВ
К.А. ЛАЙШЕВ1, Л.А. ИЛЬИНА2, Е.А. ЙЫЛДЫРЫМ2, В.А. ФИЛИППОВА2, Т.П. ДУНЯШЕВ2, А.В. ДУБРОВИН2,
Д.В. СОБОЛЕВ2, Н.И. НОВИКОВА2, Г.Ю. ЛАПТЕВ2, А.А. ЮЖАКОВ1, Т.М. РОМАНЕНКО3, Ю.П. ВЫЛКО3
Некробактериоз — инфекционное заболевание, поражающее многие виды домашних и диких млекопитающих, птиц, человека. Основные клинические проявления болезни связывают с развитием гнойно-некротических поражений кожи, слизистых, внутренних органов, конечностей в результате инфицирования строго анаэробными патогенными фузобактериями — Fusobacterium necrophorum. Для оленеводства некробактериоз северных оленей (Rangifer tarandus) относится к одной из наиболее серьезных проблем, поскольку он становится причиной существенных потерь в экономической и хозяйственной деятельности населения в районах Крайнего Севера. В представленном исследовании нами впервые проанализированы различия между составом микробиоты рубца у клинически здоровых северных оленей и животных с проявлениями некробактериоза. Целью исследования была характеристика микробиоты рубца северного оленя при некробактериозе с использованием молекулярных методов. Объектом исследования были северные олени ненецкой породы (здоровые и с признаками некробактериоза), в том числе телята (4-6 мес) и взрослые животные (3-6 лет). Образцы содержимого рубца отбирали в летне-осенний период в 2017 году (n = 3 для каждой возрастной группы) в Ямало-Ненецком автономном округе (п.г.т. Харп, лесотундровая природно-климатическая зона). Общее число бактерий и грибов класса Neocallimastigales анализировали методом количественной ПЦР, состав бактериального сообщества — методом Т-RFLP (terminal restriction fragment length polymorphism). У особей с клиническими проявлениями некробактериоза детектировано достоверно более высокое содержание фузобактерий: у взрослых оленей в 1,79 раза (p < 0,05), у телят — в 2,65 раза (p < 0,05). У больных животных обеих возрастных групп отмечали более значительное присутствие бактерий рода Staphylocoссus (p < 0,05) и семейства Pseudomonadaceae (p < 0,05), часть видов которых могут становиться причиной гнойно-некротических поражений. У 4-6-месячных телят выявлено достоверное увеличение (p < 0,05) численности кампилобактерий (семейство Сampylobacteriaceae) и энтеробактерий (семейства Enterobacteriaceae) по сравнению с клинически здоровыми животными. При этом у здоровых особей обнаружено больше целлюлозолитических бактерий класса Clostridia (в том числе представителей семейств Lachnospiraceae, Eubacteriaceae и Clostridiaceae), филума Bacteroidetes и бактерий порядка Negativicutes, ферментирующих кислоты. Отмечено, что в целом микробиом рубца телят с клиническими признаками некробактериоза характеризовался большими изменениями по сравнению со взрослыми животными. В частности, у молодняка северных оленей с симптомами некробактериоза наблюдали достоверное увеличение индекса видового разнообразия Шеннона микробиоты рубца (р < 0,05), что свидетельствует о большей неоднородности бактериального сообщества по сравнению с таковым у здоровых особей. Также в рубце животных из этой группы было выявлено снижение (р < 0,05) численности целлюлозолитических грибов-хитридиомицетов класса Neocallimastigales. На основании полученные нами результатов могут быть разработаны рекомендации для повышения эффективности мероприятий против некробактериоза северных оленей и снижения падежа животных в летне-осенний период.
Ключевые слова: Rangifer tarandus, северный олень, некробактериоз, заболевания копыт, фузобактерии, микробиом рубца, Российская Арктика, T-RFLP-анализ, количественная ПЦР.
В естественном ареале северных оленей (Rangifer tarandus) постоянно регистрируются инфекционные и инвазионные болезни (1). Они наносят значительный урон оленеводству, которое стратегически значимо для населения Крайнего Севера. Особо выделяются паразитарные болезни, передаваемые кровососущими насекомыми, оводовые инвазии (эдемагеноз, цефеномийоз) и инфекционное бактериальное заболевание некробактериоз. Доля больных некробактериозом животных в среднем составляет от 7 до 33 %. Наиболее сильно некробактериоз проявляется в летний период, когда доля больных животных может достигать 65-70 % (2).
Некробактериоз поражает многие виды домашних и диких млекопитающих животных, птиц, а также человека. Основные клинические проявления болезни — гнойные некрозы кожи, слизистых, внутренних органов в результате инфицирования строго анаэробными патогенными фузобактериями, преимущественно Fusobacterium necrophorum (3, 4). У жвачных животных страдают конечности. У северных оленей гнойные поражения копыт отмечают только в летне-осенний период (2, 5), преимущественно в июле, августе, с наступлением жары, в сентябре число заболеваний резко снижается, а в октябре новые случаи уже не регистрируются. Это связано с созданием оптимальных для развития инфекций сезонных условий — плюсовых температур воздуха, истощения животных, активности насекомых. Вспышки некробактериоза происходят как на территории Российской Арктики (5), так и в других ареалах северных оленей (6).
Более всего некробактериоз изучен у крупного рогатого скота (КРС) (6, 7), тогда как исследований, касающихся северных оленей, намного меньше. В последнее время наиболее активно обсуждается возможность взаимосвязей между возбудителем некробактериоза и микрофлорой других экотопов организма животного, в частности рубца (8, 9).
Многие авторы полагают, что здоровье жвачных животных в значительной степени зависит от рубцового пищеварения и, следовательно, обеспечивается наличием в рубце определенных микроорганизмов (10). По современным представлениям, F. necroforum относится к нормофлоре пищеварительного тракта жвачных, в особенности рубца, и может распространяться в окружающей среде через экскременты (6, 11). Известно, что фузобактерии способны выделять ряд токсинов, которые приводят к некрозам тканей и возникновению вторичных инфекций, вызываемых актинобактериями. Сообщалось, что фузобактерии способны проникать только в поврежденные ткани, например при некротических поражениях конечностей (12). Также установлено, что фузобактерии попадают в организм животного через геморрагические повреждения в пищеварительном тракте (7).
Причины, повышающие подверженность северных оленей заражению фузобактериями, изучены мало. По-видимому, его может провоцировать стресс, повышенная или пониженная температура, перенаселенность и скудное питание (12, 13). Так, норвежские исследователи, изучив вспышку некробактериоза у северных оленей, обитающих на севере Норвегии (2007-2008 годы), подтвердили, что возбудителем инфекции была бактерия F. necroforum, и пришли к выводу, что заболевание спровоцировано более высокими (по сравнению со средними) значениями температуры и влажности (6).
Разнообразие патогенных и условно-патогенных бактерий в рубце северных оленей при некробактериозах практически не описано.
В представленном исследовании нами впервые проанализированы различия между составом микробиоты рубца у клинически здоровых северных оленей Арктической зоны Российской Федерации и животных с проявлениями некробактериоза. У особей с симптомами некробактериоза детектировано меньшее количество целлюлозолитических и утилизирующих кислоты бактерий, а также большее — фузобактерий и других патогенов (стафилококков, псевдомонад и др.), часть видов которых могут становиться причиной гнойно-некротических поражений животных. Наиболее выраженные изменения в составе микробиома рубца в результате некробактериоза были детектированы у 4-6-месячных телят, у которых бактериальное сообщество рубца оказалось более неоднородным и характеризовалось повышенной численностью кампилобактерий, энтеробактерий и снижением представленности грибов-хитридиомицетов.
Целью работы было исследование микробиоты рубца северного оленя с клиническими проявлениями некробактериоза с использованием молекулярно-биологических методов.
Методика. Образцы содержимого рубца отбирали с помощью зонда у 4-6-месячных телят и 3-6-летних взрослых северных оленей (Rangifer tarandus) ненецкой породы (клинически здоровые животные и особи с проявлениями некробактериоза; n = 3 из каждой возрастной группы) в летне-осенний период в 2017 году в Ямало-Ненецком автономном округе (п.г.т. Харп, лесотундровая природно-климатическая зона). Образцы замораживали при температуре -20 °С до проведения анализа.
Тотальную ДНК из образцов выделяли при помощи набора Genomic DNA Purification Kit («Fermentas, Inc.», Литва) в соответствии с рекомендациями производителя. Конечную концентрацию выделенной тотальной ДНК в растворе определяли на флуориметре Qubit («Invitrogen, Inc.», США) с применением набора Qubit dsDNA BR Assay Kit («Invitrogen, Inc.», США) в соответствии с инструкцией производителя.
Общую численность бактерий и грибов-хитридиомицетов класса Neocallimastigales анализировали методом количественной ПЦР (амплификатор ДТ Lite-4; ООО «НПО ДНК-Технология», Россия) с набором реактивов для ПЦР-РВ в присутствии интеркалирующего красителя EVA Green (ЗАО «Синтол», Россия). Использовали праймеры F — 5´-ACTCCTAC-GGGAGGCAGCAG-3´, R — 5´-ATTACCGCGGCTGCTGG-3´ (бактерии); F — 5´-GCACTTCATTGTGTGTACTG-3´, R — 5´-GGATGAAACTCGTTG-ACTTC-3´ (грибы). Режим амплификации: 3 мин при 95 °С (1 цикл); 13 с при 95 °С, 13 с при 57 °С, 30 с при 72 °С (40 циклов).
Состав бактериального сообщества рубца северных оленей анализировали методом Т-RFLP (terminal restriction fragment length polymor-phism) (14). ПЦР осуществляли на ДНК-амплификаторе Verity («Life Technologies, Inc.», США) с эубактериальными праймерами 63F (5´-CAGGCCTAACACATGCAAGTC-3´) с меткой на 5´-конце (флуорофор WellRed D4, «Beckman Coulter», США) и 1492R (5´-TACGGHTACCTTG-TTACGACTT-3´), позволяющими амплифицировать фрагмент гена 16S pРНК (от 63-й до 1492-й позиции; нумерация указана для гена 16S pРНК Esherichia coli), в следующем режиме: 3 мин при 95 °С (1 цикл); 30 с при 95 °С, 40 с при 55 °С, 60 с при 72 °С (35 циклов); 5 мин при 72 °С.
Флуоресцентно меченные ампликоны гена 16S pРНК очищали по стандартной методике (15). Их обработку (30-50 нг ДНК) проводили рестриктазами HaeIII, HhaI и MspI («Fermentas», Литва), в течение 2 ч при 37 °С. Продукты рестрикции осаждали этанолом, затем добавляли 0,2 мкл маркера молекулярной массы Size Standart-600 («Beckman Coulter», США) и 10 мкл формамида Sample Loading Solution («Beckman Coulter», США). Анализ проводили на CEQ 8000 («Beckman Coulter», США) согласно рекомендациям производителя (погрешность прибора не более 5 %). Полученные данные обрабатывали в программе Fragment Analysis («Beckman Coulter», США). Вычисляли размеры пиков и их площади, на основании чего выделяли подтипы (филотипы) с принятой в исследовании погрешностью в 1 нуклеотид и оценивали их долю в микробном сообществе.
Таксономическую принадлежность бактерий определяли с использованием базы данных (http://mica.ibest.uidaho.edu/trflp.php).
Статистическую обработку результатов осуществляли методом дисперсионного анализа с использованием программного обеспечения Microsoft Excel 2010. Рассчитывали средние (M), стандартные ошибки средних (±SEМ), достоверность различий между изучаемыми группами оценивали по критерию t-Стьюдента. В программе Past рассчитывали индексы биоразнообразия Шеннона и Симпсона (http://folk.uio.no/ohammer/past/).
Результаты. Согласно оценке бактериального сообщества методом T-RFLP, большая часть филотипов относилась к филумуFirmicutes (их общая доля достигала у взрослых северных оленей 61 %). В меньшей степени в микробиоме рубца клинически здоровых животных и особей с симптомами некробактериоза оказались представлены бактерии филумов Bacteroidetes, Actinobacteria и Proteobacteria. В минорном количестве выявлялись представители филумов Fusobacteria, Acidobacteria, Cyanobacteria. Также существенной оказалась доля филотипов, которые не удалось идентифицировать по базам данных. Наибольшее присутствие неидентифицированных таксонов отмечали у взрослых особей (до 22,18 %).
Микробиом рубца животных с клиническими проявлениями некробактериоза имел значимые отличия от такового у здоровых особей. Прежде всего у северных оленей с симптомами некробактериоза следует отметить достоверно более высокое содержание представителей филума Fusobacteria (рис. 1): у взрослых особей — в 1,79 раза (p < 0,05), у телят — в 2,65 раза (p < 0,05). Полученные результаты в очередной раз подтверждают роль бактерий рода Fusobacterium (в том числе F. necrophorum) в этиологии некробактериоза.
Тем не менее имеются сообщения, авторы которых высказывали сомнения по этому поводу или приводили доказательства основной роли других видов микроорганизмов в этиологии развития некробактериоза у жвачных (16). В связи с этим интересен выявленный нами факт совместного присутствия и роста численности ряда патогенов, включая возбудителей гнойно-некротических инфекций, в рубце у обследованных особей с симптомами некробактериоза. Так, в обеих возрастных группах северных оленей с некробактериозными проявлениями достоверно повышалась доля бактерий рода Staphylocoссus (p < 0,05) и семейства Pseudomonadaceae (p < 0,05), часть видов которых могут становиться причиной гнойно-некротических инфекций животных. Кроме того, у 4-6-месячных телят выявили рост присутствия кампилобактерий (сем. Сampylobacteriaceae) и энтеробактерий (сем. Enterobacteriaceae) — соответственно в 1,96 (р < 0,05) и 2,38 раза (р < 0,05) по сравнению с клинически здоровыми животными.
Полученные результаты подтверждают сообщения о полиэтиологической природе некробактериоза. Так, G.R. Smith с соавт. (17, 18) показали присутствие сопутствующих фузобактериям микроорганизмов, в числе которых отмечены энтеробактерии и актинобактерии, осложняющие течение некробактериоза. Аналогичного мнения придерживались М. Петров и С. Ташев (цит. по 3), которые при обследовании 134 северных оленей с гнойными поражениями конечностей выявили синегнойную палочку (Pseudomonas aeruginosa), стрептококки (род Streptococcus), стафилококки (род Staphylococcus) и их ассоциации. А.X. Лайшев с соавт. (5) сообщают о выделении вместе с фузобактериями микрококка, кишечной палочки, стафилококка, протея при экспериментальном заражении оленей чистой культурой возбудителя некробактериоза. По данным J.E. Nocek (7), размножение патогенных микроорганизмов, обитающих на конечностях КРС (фузобактерий, стафилококков, стрептококков, энтеробактерий), наряду с повышением в крови содержания эндотоксинов и гистамина (продуктов лизиса рубцовых микроорганизмов) и повреждением кровеносных сосудов вызывает воспаление и некрозы дермальных слоев копыт. Н.С. Островский с соавт. (19) объясняли массовое распространение болезней конечностей у северных оленей нарушением технологии содержания, ведущим к травматизму и мацерации кожи, ослаблению ее защитных свойств, с последующим внедрением микрофлоры в ткани.
В нашей работе бактериальное сообщество рубца клинически здоровых животных отличалось от такового у особей с симптомами некробактериоза большим процентным содержанием микроорганизмов, участвующих в ферментации углеводов растительных кормов. Так, у телят с симптомами некробактериоза была меньше (в 1,32 раза, р < 0,05) долябактерий класса Clostridia (в том числе представителей семейств Lachnospiraceae, Eubacteriaceae и Clostridiaceae), потенциально обладающих способностью к ферментации полисахаридов растительных кормов с образованием летучих жирных кислот. Кроме того, у взрослых и молодых больных особей (по сравнению со здоровыми животными) регистрировали достоверно меньшую (р < 0,05) численность бактерий филума Bacteroidetes (включающего роды Bacteroides, Prevotella), ферментирующих крахмал, клетчатку, ряд других углеводов, белков, а также дезаминирующих аминокислоты. У телят в меньшем количестве присутствовали бактерий порядка Negativicutes (родов Megasphaera,Selenomonas), которые препятствуют снижению рН и развитию лактатного ацидоза за счет утилизации органических кислот (пропионовой, уксусной, масляной, молочной и др.), образуемых в процессе ферментации растительной клетчатки (7, 20, 21).
Расчет экологических индексов также продемонстрировал общее снижение биоразнообразия в рубце животных с клиническими проявлениями некробактериоза (рис. 3). В частности, у молодняка с симптомами некробактериоза наблюдалось увеличение индекса Шеннона (р < 0,05), что свидетельствуют о большей неоднородности бактериального сообщества их рубца по сравнению со здоровыми особями.
Следует отметить, что общее количество бактерий у здоровых и заболевших особей достоверно не различалось (рис. 4). Однако у молодняка северных оленей в рубце выявили достоверное (р < 0,05) снижение численности грибов-хитридиомицетов, которые, как известно, вырабатывают широкий спектр многофункциональных полисахаридных ферментов (22).
Ранее сообщалось, что нарушение баланса микробиоты в пищеварительном тракте на фоне заражения некробактериозом приводит к более быстрому размножению патогенного штамма фузобактерии (23, 24). Кроме того, инфицирование организма КРС фузобактериями может происходить в результате развития синдрома лактатного ацидоза, который характеризуется нарушением состава рубцовой микробиоты (4, 7). При этом в рубце КРС наблюдается снижения численности целлюлозолитических и утилизирующих кислоты бактерий, а также увеличение количества микроорганизмов родов Streptococcusи Lactobacillus, синтезирующих лактат, который снижает рН в рубце (25). Развитие лактатного ацидоза приводит к повреждению эпителия рубца и проникновению патогенных фузобактерий через слизистую в кровь. Это обусловливает дальнейшее инфицирование организма, при котором в зависимости от биотипа возбудителя (А, В или АВ) наблюдается соответствующая клиническая картина (поражения копыт, слизистых или кожи, абсцессы печени или других внутренних органов). Так, для КРС неоднократно показано одновременное выделение фузобактерий из других органов (печени, почек) и рубца (7, 8). Интересно, что у исследованных нами северных оленей не наблюдали увеличения представленности в рубце лактобактерий, что свидетельствует о необходимости продолжить изучение связи микробиома рубца с развитием некробактериоза. Так, в качестве причин развития некробактериоза у северных оленей рассматривают потребление животными зараженного корма, а также проникновение микроорганизмов через респираторный тракт, что показано при исследовании вспышки некробактериоза в США, вызванного поражением респираторного тракта фузобактериями F. necrophorum и F. varium у белохвостого оленя (Odocoileus virginianus) (8, 26).
Таким образом, у северных оленей с проявлениями некробактериоза по сравнению с клинически здоровыми животными существенно нарушается структура рубцового микробиома. У последних выявлялось больше целлюлозолитических и утилизирующих кислоты бактерий и меньше — фузобактерий и иных патогенов (стафилококков, псевдомонад и др.). Микробиом рубца у телят с клиническими признаками некробактериоза характеризовался более значительными изменениями, чем у больных взрослых особей. Эти различия выражались в большей неоднородности бактериального сообщества, росте представленности кампилобактерий, энтеробактерий, а также в снижении численности грибов-хитридиомицетов. Следовательно, выявленные закономерности могли быть обусловлены физиологическими особенностями этого этапа развития у северных оленей. Полученные в исследовании результаты могут быть использованы в качестве основы для разработки рекомендаций по повышению эффективности мероприятий против некробактериоза северных оленей и снижению падежа животных в летне-осенний период.
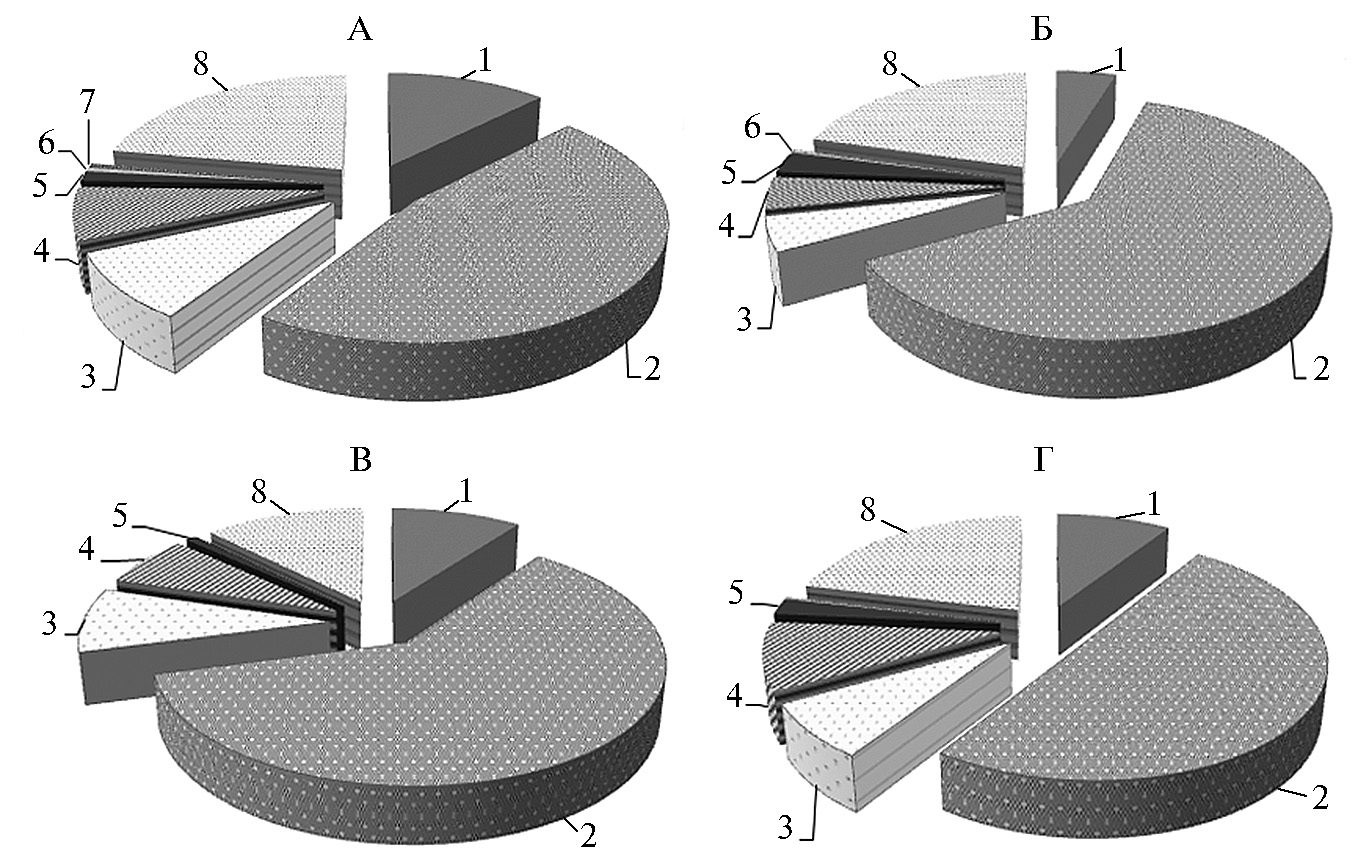
Рис. 1. Представленность филумов бактерий в рубце северных оленей (Rangifer tarandus): А — взрослые особи, Б — взрослые особи с клиническими признаками некробактериоза, В — телята, Г — телята с клиническими признаками некробактериоза; 1 — филум Bacteroidetes, 2 — филум Firmicutes, 3 — филум Actinobacteria, 4 — филум Proteobacteria, 5 — филум Fusobacteria, 6 — филум Cyanobacteria, 7 — филум Acidobacteria, 8 — неклассифицированные нуклеотидные последовательности (Ямало-Ненецкий автономный округ, п.г.т. Харп, 2017 год).
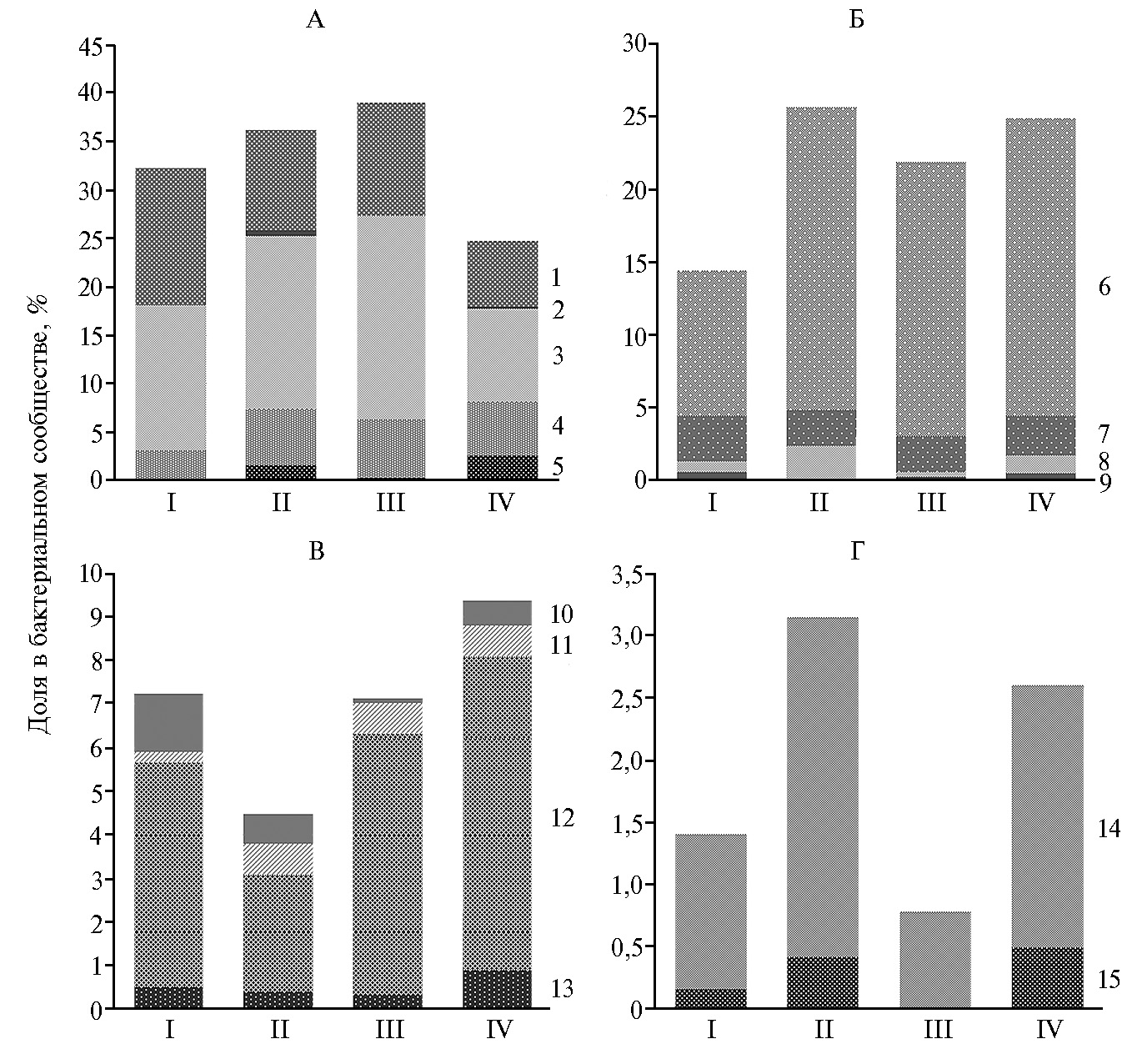
Рис. 2. Представленность бактериальных таксонов в рубце северных оленей (Rangifer tarandus): I — взрослые особи, II — взрослые особи с клиническими признаками некробактериоза, III — телята, IV — телята с клиническими признаками некробактериоза (Ямало-Ненецкий автономный округ, п.г.т. Харп, 2017 год).
А — класс Clostridia филума Firmicutes: 1 — cем. Thermoanaerobacteraceae, 2 — сем. Lachnospiraceae, 3 — сем. Eubacteriaceae, 4 — сем. Ruminococcaceae, 5 — сем. Clostridiaceae.
Б — другие представители класса Clostridia филума Firmicutes: 6 — пор. Negativicutes, 7 — р. Bacillus,8 — р. Lactobacillus, 9 — р. Peptococcus.
В — филум Proteobacteria: 10 — сем. Burkholderiaceae; 11 — сем. Pseudomonadaceae, 12 — сем. Сampylobacteriaceae, 13 — сем. Enterobacteriaceae.
Г — прочие: 14 — филум Fusobacteria, 15 — р. Staphylococcus
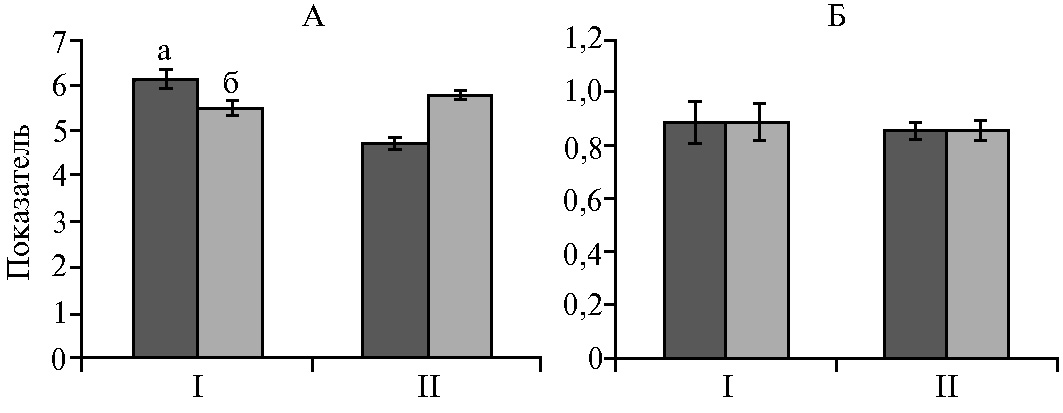
Рис. 3. Показатели биоразнообразия бактериального сообщества рубца у взрослых особей (I) и телят (II) северных оленей (Rangifer tarandus): А — индекс Шеннона, Б — индекс Симпсона; а — клинически здоровые животные, б — особи c проявлениями некробактериоза (M±SEM, Ямало-Ненецкий автономный округ, п.г.т. Харп, 2017 год).
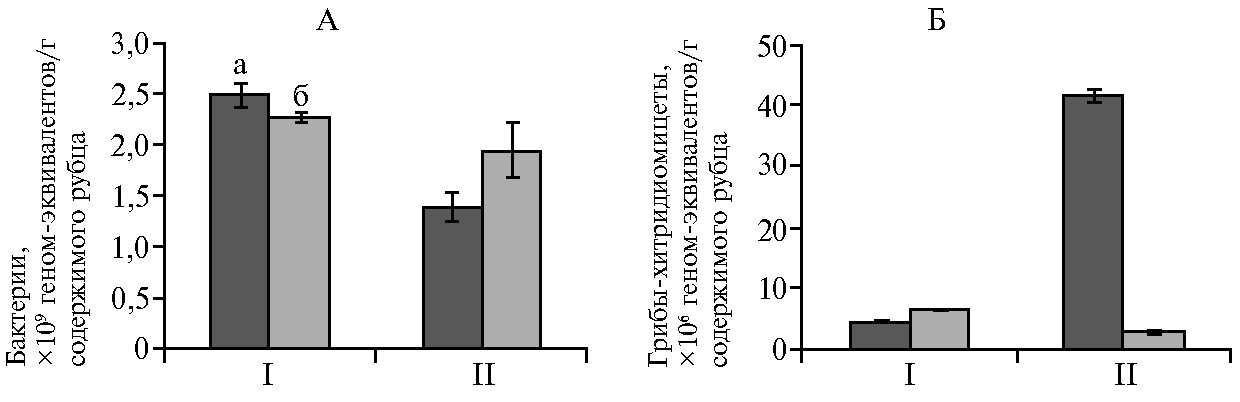
Рис. 4. Количество микроорганизмов в рубце у взрослых особей (I) и телят (II) северных оленей (Rangifer tarandus): А — бактерии, Б — грибы-хитридиомицеты; а — клинически здоровые животные, б — особи c проявлениями некробактериоза (M±SEM, Ямало-Ненецкий автономный округ, п.г.т. Харп, 2017 год).
ЛИТЕРАТУРА
- Haigh J., Berezowski J., Woodbury M.R. A cross-sectional study of the causes of morbidity and mortality in farmed white-tailed deer. Can. Vet. J., 2005, 46(6): 507-512.
- Самандас А.М., Лайшев К.А. Изучение этиологических особенностей некробактериоза северных оленей. Сибирский вестник< сельскохозяйственной науки, 2010, 10(214): 48-52.
- Самоловов А.А. Некробактериоз животных. Новосибирск, 1993.
- Tadepalli S., Narayanan S.K., Stewart G.C., Chengappa M.M., Nagaraja T.G. Fusobacterium necrophorum: a ruminal bacterium that invades liver to cause abscesses in cattle. Anaerobe, 2009, 15(1-2): 36-43 (doi: 10.1016/j.anaerobe.2008.05.005).
- Лайшев А.Х., Маслухина А.Г. О наличии «фактора распространения» у возбудителя некробациллеза оленей. Труды НИИСХ КрайнегоСевера, 1966, 13: 37-38.
- Handeland K., Boye M., Bergsjø B., Bondal H., Isaksen K., Agerholm J.S. Digital necrobacillosis in Norwegian wild tundra reindeer (Rangifer tarandus tarandus). Journal of Comparative Pathology, 2010, 143(1): 29-38 (doi: 10.1016/j.jcpa.2009.12.018).
- Nocek J.E. Bovine acidosis: implications on laminitis. J. Dairy Sci., 1997, 80: 1005-1028 (doi: 10.3168/jds.S0022-0302(97)76026-0).
- Brooks J.W., Kumar A., Narayanan S., Myers S., Brown K., Nagaraja T.G., Jayarao B.M. Characterization of Fusobacterium isolates from the respiratory tract of white-tailed deer (Odocoileus virginianus). Journal of Veterinary Diagnostic Investigation, 2014, 26(2): 213-220 (doi: 10.1177/1040638714523613).
- Kupca A.M., Rettinger A., Zimmermann P., Hörmansdorfer S., Konrad R., Hafner-Marx A. Severe purulent and necrotizing glossitis in a fallow deer (Dama dama) due to an infection with the involvement of Mannheimia granulomatis. Berl. Munch. Tierarztl. Wochenschr., 2015, 128(7-8): 285-288.
- Zeineldin M., Barakat R., Elolimy A., Salem A.Z.M., Elghandour M.M.Y., Monroy J.C. Synergetic action between the rumen microbiota and bovine health. Microbial Pathogenesis, 2018, 124: 106-115 (doi: 10.1016/j.micpath.2018.08.038).
- Aagnes T.H., Sørmo W., Mathiesen S.D. Ruminal microbial digestion in free-living, in captive lichen-fed, and in starved reindeer (Rangifer tarandus tarandus) in winter. Appl. Environ. Microbiol., 1995, 61(2): 583-591.
- Woodbury M.R., Chirino-Trejo M. Necrobacillosis in white-tailed deer. Proceedings of the 1st World Deer Veterinary Congress and the Deer Branch of the New Zealand Veterinary Association. The Deer Branch New Zealand Veterinary Association, Wellington, 2004: 21-23.
- Haigh J.C., Robert J.H. Farming wapiti and red deer. Mosby, St. Louis, 1993.
- Лаптев Г.Ю., Новикова Н.И., Ильина Л.А., Йылдырым Е.А., Нагорнова К.В., Думова В.А., Солдатова В.В., Большаков В.Н., Горфункель Е.П., Дубровина Е.Г., Соколова О.Н., Никонов И.Н., Лебедев А.А. Нормы содержания микрофлоры в рубце крупного рогатого скота. СПб, 2016.
- Маниатис Т., Фрич Э., Сэмбрук Дж. Молекулярное клонирование. М., 1984.
- Handeland K., Boye M., Bergsjø B., Bondal H., Isaksen K., Agerholm J.S. Digital necrobacillosis in Norwegian wild tundra reindeer (Rangifer tarandus tarandus). Journal of Comparative Pathology, 2010, 142(1): 29-38 (doi: 10.1016/j.jcpa.2009.12.018).
- Smith G.R., Till D., Wallace L.M., Noakes D.E. Enhancement of the infectivity of Fusobacterium necrophorum by other bacteria. Epidemiol. Infect., 1989, 102(3): 447-458.
- Li Y., Hu X., Yang S., Zhou J., Qi L., Sun X., Fan M., Xu S., Cha M., Zhang M1, Lin S., Liu S., Hu D. Comparison between the fecal bacterial microbiota of healthy and diarrheic captive musk deer. Front Microbiol., 2018, 9: 300 (doi: 10.3389/fmicb.2018.00300).
- Островский Н.С., Мажуга Е.П. Лечение и профилактика гнойно-некротических заболеваний пальцев крупного рогатого скота. В сб.:Профилактика незаразных болезней сельскохозяйственных животных. М., 1977: 231-234.
- Church D.C. The ruminant animal: digestive physiology and nutrition. Prentice Hall, New Jersey, 1993.
- Hungate R.E. The rumen and its microbes. Academic Press, NY, 1966.
- Wang T.Y., Chen H.L., Lu M.J., Chen Y.C., Sung H.M., Mao C.T., Cho H.Y., Ke H.M., Hwa T.Y., Ruan S.K., Hung K.Y., Chen C.K., Li J.Y., Wu Y.C., Chen Y.H., Chou S.P., Tsai Y.W., Chu T.C., Shih C.A., Li W.H., Shih M.C. Functional characterization of cellulases identified from the cow rumen fungus Neocallimastix patriciarum W5 by transcriptomic and secretomic analyses. Biotechnology for Biofuels, 2011 4: 24. (doi: 10.1186/1754-6834-4-24).
- Smith G.R., Thornton E.A. Effect of disturbance of the gastrointestina microflora on the faecal excretion of Fusobacterium necrophorum biovar A. Epidemiology and Infection, 1993, 110(2): 333-337.
- Nagaraja T.G., Narayanan S.K., Stewart G.C., Chengappa M.M. Fusobacterium necrophorum infections in animals: pathogenesis and pathogenic mechanisms. Anaerobe, 2005, 11(4): 239-246. (doi: 10.1016/j.anaerobe.2005.01.007).
- Chen L., Shen Y., Wang C., Ding L., Zhao F., Wang M., Fu J., Wang H. Megasphaera elsdenii lactate degradation pattern shifts in rumen acidosis models. Front. Microbiol., 2019, 10: 162 (doi: 10.3389/fmicb.2019.00162).
- Chirino-Trejo M., Woodbury M.R., Huang F. Antibiotic sensitivity and biochemical characterization of Fusobacterium spp. and Arcanobacterium pyogenes isolated from farmed white-tailed deer (Odocoileus virginianus) with necrobacillosis. Journal of Zoo and Wildlife Medicine, 2003, 34(3): 262-268 (doi: 10.1638/02-019).