doi: 10.15389/agrobiology.2019.4.798rus
УДК 36.52/.58:636.084.416:579.6
Исследование выполнено при поддержке гранта РНФ для реализации научного проекта 16-16-04089-П «Изучение физиологических и микробиологических особенностей пищеварения кур мясных пород в эмбриональный и постэмбриональный периоды для создания новых технологий кормления, обеспечивающих максимально полную реализацию генетического потенциала птицы».
ЗАМЕЩЕНИЕ КОРМОВЫХ АНТИБИОТИКОВ В РАЦИОНАХ. Сообщение II. МИКРОБИОТА КИШЕЧНИКА И ПРОДУКТИВНОСТЬ МЯСНЫХ КУР (Gallus gallus L.) НА ФОНЕ ФИТОБИОТИКА
И.А. ЕГОРОВ1, Т.А. ЕГОРОВА1, Т.Н. ЛЕНКОВА1, В.Г. ВЕРТИПРАХОВ1, В.А. МАНУКЯН1, И.Н. НИКОНОВ1, А.А. ГРОЗИНА1, В.А. ФИЛИППОВА2, Е.А. ЙЫЛДЫРЫМ2, Л.А. ИЛЬИНА2, А.В. ДУБРОВИН2, Г.Ю. ЛАПТЕВ2
Реализовать генетический потенциал продуктивности можно только у здоровой птицы. В связи с запретом использования кормовых антибиотиков в странах ЕС ведется поиск альтернативы этим препаратам. Такие добавки, как пробиотики, пребиотики, синбиотики, симбиотики, подкислители и фитобиотики, не уступают антибиотикам по эффективности, но исключают негативные последствия. В настоящей работе впервые представлены результаты выращивания молодняка мясных кур (Gallus gallus domesticus) исходных линий Б5 и Б9 селекции СГЦ «Смена» с использованием фитобиотика Интебио на основе эфирных масел чеснока, лимона, чабреца и эвкалипта. Целью работы было изучение влияния фитобиотика на основе эфирных масел на рост и половое развитие молодняка кур, активность пищеварительных ферментов и состояние микробиоценоза кишечника в сравнении с кормовым антибиотиком. Для эксперимента отбирали 1-су-точных цыплят (две группы по 50 гол. в каждой), которых выращивали до 21-недельного возраста (СГЦ «Загорское ЭПХ», Московская обл.). Комбикорма для обеих групп птицы были растительного типа, их балансировали по всем питательным веществам. Контрольная группа получала комбикорма с добавкой кормового антибиотика Бацитрацина-30 (активность 42 EД/мг) в количестве 100 г/т в течение всего опытного периода. Птица опытной группы получала кормовую добавку Интебио (ООО «БИОТРОФ», г. Санкт-Петербург) в количестве 1000 г/т корма. Установлено, что в опытных группах живая масса петушков и курочек обеих линий в 21-недельном возрасте не различалась с контролем и составила в линии Б5 3172 и 2318 г против 3169 и 2316 г в контрольной группе, в линии Б9 — 2590 и 1917 г против 2589 и 1920 г. Развитие репродуктивных органов самцов (семенники) и самок (яичники и яйцеводы) было в пределах нормы и не имело существенных различий между опытными и контрольными группами. Результаты физиологического (балансового) опыта, проведенные в 4- и 7-недельном возрасте птицы, согласуются с данными по живой массе молодняка. Существенных различий по переваримости сухого вещества корма, жира, утилизации азота корма между птицей контрольных и опытных групп не установлено. Однако петушки и курочки линии Б5 на 3,11 % лучше переваривали сухое вещество корма при более высокой (на 2,12 %) степени использования азота. Переваримость жира также была выше на 2,95 %. Исследования на фистулированной птице показали, что добавка в комбикорма фитобиотика Интебио способствовала повышению активности липазы в дуоденальном содержимом у молодняка линий Б5 и Б9 соответственно на 30,9 (р ≤ 0,05) и 98,3 % (р ≤ 0,01), а также протеаз на 36,4 % (р ≤ 0,05) в линии Б5. При этом у кур линии Б9 липолитическая активность была ниже как в содержимом кишечника (на 59,9 %, р ≤ 0,001), так и в плазме крови (на 48,3 %, р ≤ 0,001). T-RFLP-анализ бактериального сообщества дуоденального содержимого позволил установить присутствие значительного количества филотипов микроорганизмов, общее число которых в зависимости от породы птицы и введения в рацион фитобиотика составляло от 125,65±3,12 до 170,36±6,09. По результатам оценки таксономической принадлежности большинство выявленных филотипов были отнесены к филумам Firmicutes, Bacteroidetes и Proteobacteria, суммарная доля которых в дуоденальном содержимом кишечника птиц составляла не менее 77,37±4,29 и максимально достигала 95,69±6,15 %. В меньшей степени были представлены бактерии филума Actinobacteria, в минорном количестве обнаруживались представители филумов Tenericutes и Fusobacteria. В ответ на применение фитобиотика наблюдалось достоверное увеличение представленности членов филума Bacteroidetes,а также класса Clostridiales, включающего представителей семейств Eubacteriaceae, Clostridiaceae, Lachnospiraceae (p ≤ 0,05), что свидетельствует о повышении потенциала ферментации растительных полисахаридов микробным сообществом.
Ключевые слова: Gallus gallus domesticus, молодняк, исходные линии, живая масса, пищеварительные ферменты, фитобиотик, микрофлора кишечника.
Мясная птица может достичь высоких продуктивных и воспроизводительных качеств только при условии полноценного нормированного кормления. От этого в значительной степени зависит результативность работы генетиков и селекционеров. Питательность кормов, их количество и качество должны обеспечить запланированные селекционируемые показатели по неделям жизни, начиная с 7-суточного возраста птицы. На всех этапах выращивания молодняка исходных линий необходимо скармливать комбикорма, сбалансированные по обменной энергии, питательным, минеральным и биологически активным веществам с учетом их доступности. Они должны соответствовать существующим ветеринарно-санитарным и гигиеническим требованиям и быть нетоксичными (1).
В настоящее время большинство стран Европы ввели запрет на включение в корма для птицы кормовых антибиотиков. Основное внимание уделяется кормовым добавкам, которые могли бы заменить кормовые антибиотики без значительных изменений рецептов комбикормов (2-4).
Широкое применение антибиотиков и химических антибактериальных средств нередко приводит к ухудшению здоровья птицы, связанному с развитием неконтролируемых секундарных инфекций — сальмонеллезов, кампилобактериозов, стафилококкозов, клостридиозов, а также полимикробных заболеваний (5, 6). Патогенные микроорганизмы вызывают нарушение состава кишечной микробиоты, приводят к изменению толщины, внешнего вида, мышечного тонуса, прочности и повышенной парацеллюлярной проницаемости стенок кишечника для токсических метаболитов, что в итоге негативно отражается на состоянии здоровья и продуктивности стада. Актуальным вопросом остается и контаминация продукции птицеводства различными патогенами — возбудителями инфекционных заболеваний человека (7, 8).
Изучение свойств экстрактов и эфирных масел растений считается одним из наиболее перспективных подходов для создания препаратов для профилактики заболеваний и увеличения продуктивности сельскохозяйственной птицы. В отличие от антибиотиков и лекарств, полученных посредством химического синтеза, вещества, выделенные из растений, характеризуются меньшей токсичностью, не аккумулируются в организме и могут стать идеальным сырьем для создания лекарственных препаратов (9). Кроме того, растения представляют собой неограниченный возобновляемый источник биологически активных веществ, включающих до 12 тыс. соединений, из которых описано около 2 тыс. (10). Интерес к ним обусловлен их антибактериальным действием и возможностью применения вместо лечебных и кормовых антибиотиков, а также другими свойствами, влияющими на обмен веществ и продуктивность животных (11-15). Однако данные о многих аспектах воздействия эфирных масел все еще остаются противоречивыми, что может объясняться различием питательного рациона, условий содержания, породными особенностями птицы.
Потребность в племенной продукции для бройлерного производства в стране удовлетворяется в основном за счет зарубежных кроссов птицы. Однако последние годы в СГЦ «Смена» совместно с учеными Всероссийского научно-исследовательского и технологического института птицеводства РАН ведется работа по созданию нового отечественного кросса мясных кур (16, 17). К его основным преимуществам над зарубежными аналогами относятся высокие жизнеспособность и генетический потенциал продуктивности, адаптированность к местным условиям кормления и содержания.
Значительный интерес представляет изучение воздействия фитобиотиков на микробиом кишечника птицы различных генетических линий. Снижение риска развития инфекционных патологий связывают с формированием здоровой микробиоты пищеварительного тракта, которая способна обеспечить высокую резистентность к колонизации кишечника патогенами (18, 19) благодаря синтезу летучих жирных кислот (ЛЖК), бактериоцинов и других соединений, сдерживающих рост и развитие патогенных видов (20, 21). Известно, что микроорганизмы, взаимодействуя между собой, а также с организмом хозяина, способны оказывать глубокое воздействие на иммунитет, неспецифическую резистентность к инфекциям и общие процессы жизнедеятельности птицы (22). Кроме того, показано активное участие микробного сообщества кишечника птиц в пищеварительных процессах при расщеплении сложных полисахаридов и белков (23, 24), в использовании и образовании питательных веществ, синтезе витаминов (25), развитии ворсинок кишечника, увеличивающих всасываемую поверхность (26, 27).
Наиболее перспективны современные подходы, основанные на применении молекулярно-генетических методов, — NGS-секвенирование (next generation sequencing) и T-RFLP-анализ (terminal restriction fragment length polymorphism) (27-30), при проведении которых отсутствует необходимость в стадии культивирования микроорганизмов.
В представленной работе впервые приведены данные по активности пищеварительных ферментов у фистулированной птицы, получавшей фитобиотик Интебио (разработка и производство ООО «БИОТРОФ», г. Санкт-Петербург), которые подтверждают, что этот препарат может служить заменой кормовому антибиотику.
Целью нашей работы было изучение влияния фитобиотика на основе эфирных масел на рост и половое развитие молодняка мясных кур (Gallus gallus domesticus), активность пищеварительных ферментов и состояние микробиоценоза кишечника в сравнении с кормовым антибиотиком.
Методика. Зоотехнический и физиологический опыты проводили на исходных линиях птицы Б5 (отцовская линия отцовской родительской породы формы корниш) и Б9 (материнская линия материнской родительской формы плимутрок) в Селекционно-генетическом центре «Загорское ЭПХ» (г. Сергиев Посад, Московская обл.) в 2017 году. С 1-суточного до 21-недельного возраста птицу содержали в клетках (по 50 гол. в группе). Влажностный, температурный и световой режимы, фронт кормления и поения соответствовали рекомендациям ВНИТИП (31). Учитывали сохранность и живую массу птицы, а также массу репродуктивных органов (семенники и яичники с яйцеводами).
В 1-ю нед выращивания молодняк получал корма вволю, без ограничений по количеству. Затем еженедельно выдачу корма фиксировали, то есть осуществляли нормированное кормление. Контрольная группа получала комбикорма растительного типа, сбалансированные по всем питательным веществам соответственно возрастным периодам, с добавкой кормового антибиотика Бацитрацина-30 (активность 42 EД/мг) в количестве 100 г/т в течение всего опытного периода. Птица опытной группы получала кормовую добавку Интебио (ООО «БИОТРОФ», г. Санкт-Петербург) в количестве 1000 г/т корма. Интебио — фитобиотик (ТУ 9362-011-50932298-2011), состоящий из носителя (пшеничные отруби, ГОСТ 7169-66), на который нанесена смесь эфирных масел (чеснока, лимона, чабреца и эвкалипта). Птицу исходных линий кормили рассыпными комбикормами следующей питательности: 1-21-е сут — 280 ккал/100 г обменной энергии, 20 % сырого протеина, 1,0 % кальция, 0,7 % фосфора, 1,15 % общего лизина, 0,95 % доступного лизина, 0,45 % общего метионина, 0,39 % доступного метионина; 22-35-е сут — 275 ккал/100 г, соответственно 18; 1,0; 0,7; 0,9; 0,76; 0,38; 0,32 %; 36-105-е сут — 265 ккал/100 г, 14; 1,0; 0,65; 0,65; 0,58; 0,30; 0,26 %; 106-147-е сут — 270 ккал/100 г, 15; 1,5; 0,7; 0,64; 0,57; 0,30; 0,26 %.
Для получения содержимого 12-перстной кишки молодняк оперировали в возрасте 6 нед, вживляя Т-образную канюлю в 1 см от места впадения трех панкреатических и двух желчных протоков в кишечник. Через 5 сут после хирургической операции, когда птица восстанавливалась, ее использовали в экспериментах. Для опытов из контрольной и опытной группы отбирали по 5 гол., учетный период составлял 10 сут. Утром после 14-часового голодания птица получала по 30 г комбикорма, через 1 ч после кормления у нее брали пробы дуоденального химуса (5,0 мл), центрифугировали при 5000 об/мин в течение 3 мин (ДМ0412, «Dragonlab», КНР), разводили надосадочную жидкость в 10 раз раствором Рингера и определяли активность пищеварительных ферментов. Активность амилазы оценивали по Смит-Рою-Уголеву (32), колориметрию выполняли на приборе КФК-3 (ОАО «Загорский оптико-механический завод», Россия) при λ = 670 нм и выражали, как количество расщепленного крахмала (мг) в расчете на 1 мл химуса в течение 1 мин (мг·мл-1·мин-1). Активность протеолитических ферментов определяли по количеству расщепленного казеина (мг·мл-1·мин-1) при фотометрическом контроле (33), оптическую плотность устанавливали на КФК-3 ЗОМЗ (λ = 450 нм), активность липазы оценивали на полуавтоматическом биохимическом анализаторе BS3000P («SINNOWA Medical Science & Technology Co., Ltd», КНР), используя набор для определения липазы («ДИАКОН-ВЕТ», Россия). Биохимические исследования крови, взятой натощак из подкрыльцовой вены, выполняли на автоматическом биохимическом анализаторе Chem Well 2900 (T) («Awareness Technology», США) с использованием соответствующих наборов реагентов («Human GmbH», Германия), а также на полуавтоматическом биохимическом анализаторе BS3000P («SINNOWA Medical Science & Technoligy Co., Ltd», КНР).
Пробы содержимого 12-перстного отдела кишечника для анализа микрофлоры у самок обеих линий опытной и контрольной групп (n = 3) отбирали в конце эксперимента (по три повторности из каждой группы) со строгим соблюдением стерильности (33) и немедленно замораживали.
Состав микрофлоры исследовали методом T-RFLP. Тотальную ДНК из образцов выделяли с помощью набора DNA Purification Kit («Fermen-tas, Inc.», Литва), следуя рекомендациям производителя. ПЦР проводили на ДНК-амплификаторе Verity («Life Technologies, Inc.», США) с использованием эубактериальных праймеров 63F (5´-CAGGCCTAACACATGCA-AGTC-3´) с меткой на 5´-конце (флуорофор D4 — WellRed) и 1492R (5´-TACGGHTACCTTGTTACGACTT-3´), которые позволяют амплифицировать фрагмент гена 16S pРНК с позициями от 63-й до 1492-й (нумерация указана для гена 16S pРНК Esсherichia coli), в следующем режиме: 3 мин при 95 °С (1 цикл); 30 с при 95 °С, 40 с при 55 °С, 60 с при 72 °С (35 циклов); 5 мин при 72 °С. Флуоресцентно меченные ампликоны гена 16S pРНК очищали по стандартной методике (35). Концентрацию очищенных фрагментов ДНК гена 16S pРНК определяли с помощью флуориметра Qubit 2.0 («Invitrogen», ФРГ). По 30-50 нг ампликонов 16S pРНК обрабатывали рестриктазами HaeIII, HhaI и MspI («Fermentas», Литва). Продукты рестрикции анализировали с помощью секвенатора CEQ 8000 («Beckman Co-ulter», США) согласно рекомендациям производителя.Принадлежность бактерий к определенной филогенетической группе определяли с использованием программы Fragment Sorter и базы данных (http://www.oardc.ohiostate.edu/trflpfragsort/index.php).
Статистическую обработку результатов выполняли с помощью компьютерной программы Microsoft Excel, определяя средние значения (M) и стандартные ошибки средних (±SEM). Достоверность различий оценивали по t-критерию Стьюдента. Различия считали статистически значимыми при p < 0,05. В программе Past рассчитывали индексы биоразнообразия Шеннона (H) и Симпсона (D) (http://folk.uio.no/ohammer/past/).
Результаты. Исследования показали, что как опытная, так и контрольная группа характеризовались 100 % сохранностью поголовья. При этом живая масса молодняка изучаемых линий в контрольной и опытной группах была практически идентичной, что свидетельствует о положительном влиянии фитобиотика на рост птицы при сравнении с кормовым антибиотиком (табл. 1). Так, в линии Б5 петушки контрольной группы имели живую массу в конце выращивания 3169 г, опытной группы — 3172 г. У курочек в 21-недельном возрасте наблюдалась та же тенденция. В линии Б9 у петушков обеих групп живая масса также не различалась, хотя по абсолютным значениям она была ниже, чем в линии Б5, что обусловлено направлением селекции этой птицы. Аналогичные результаты были получены по курочкам. Расход корма на 1 гол. за весь период выращивания по линии Б5 составил 11,305 кг, по линии Б9 — 10,934 кг. Конверсия корма в опытных группах не имела достоверных различий с контролем по обеим изученным линиям.
Добавление к рациону мясных кур препарата Интебио способствовало повышению активности липазы по сравнению с контролем (на 30,9 % при р ≤ 0,05 у кур линии Б5, на 98,3 % при р ≤ 0,001 — у линии Б9) и протеаз (у кур линии Б5 на 36,4 % при р ≤ 0,05) в дуоденальном содержимом кишечника (табл. 2). Можно предположить, что эфирные масла обладают стимулирующим действием на выработку желудочного сока птицы, который усиливает активность панкреатических ферментов. Липолитическая активность в содержимом кишечника и плазме крови у птицы линии Б9 была ниже, чем у кур линии Б5, соответственно на 59,9 и 48,3 %.
Результаты по живой массе птицы при использовании кормового антибиотика и фитобиотика, свидетельствующие о практически одинаковом влиянии изученных препаратов, были подтверждены данными о переваримости и использовании питательных веществ корма. Показатели переваримости сухого вещества, жира, а также использования азота между опытными и контрольными группами как самцов, так и самок не имели значительных различий. Отмечено только, что петушки и курочки линии Б5 лучше переваривали сухое вещество корма (на 3,11 %), жир (на 2,95 %) и лучше использовали азот (на 2,12 %).
Ремонтный молодняк должен иметь не только живую массу, соответствующую нормативам, но и хорошо развитые репродуктивные органы (семенники, яичники и яйцеводы). Масса семенников у 21-недельных петушков линии Б5 в контрольной группе составляла 7,5-9,1 г, в опытной — 7,7-9,3 г, то есть различия были незначительными. В линии Б9 в контрольной группе показатели составляли 7,2-8,7 г, в опытной — 7,4-9,0 г, что практически равнозначно. Аналогичная тенденция прослеживалась и по массе репродуктивных органов самок. Так, масса яичников и яйцеводов в контрольной группе линии Б5 составила 1,75-1,86 и 5,52-5,61 г, в опытной — 1,74-1,91 и 5,64-5,70 г; для линии Б9 — соответственно 1,73-1,95 и 5,61-5,64 г (контрольная группа), 1,77-1,95 и 5,67-5,72 г (опытная группа).
T-RFLP-анализ бактериального сообщества кишечника позволил установить присутствие значительного количества филотипов микроорганизмов, общее число которых составило в зависимости от породы птицы и введения в рацион фитобиотика от 125,65±3,12 до 170,36±6,09 (табл. 3). Часть филотипов, составляющую от 3,98±0,42 до 24,88±1,61 % в зависимости от опытной группы, не удалось идентифицировать до уровня филума, что свидетельствует о наличии в содержимом кишечника птицы абсолютно неизвестных микроорганизмов, нуклеотидные последовательности которых не имеют аналогов с описанными таксонами. Неидентифицированные последовательности выявили и на более низких таксономических уровнях.
По результатам оценки таксономической принадлежности большинство выявленных филотипов было отнесено к трем филумам — Fir-micutes, Bacteroidetes и Proteobacteria, суммарная доля которых в дуоденальном содержимом кишечника птиц составляла не менее 77,37±4,29 и максимально достигала 95,69±6,15 % (рис.). В меньшей степени были представлены бактерии филума Actinobacteria, в минорном количестве обнаружены представители филумов Tenericutes и Fusobacteria.
В бактериальном сообществе кишечника птицы детектировано значительное количество условно-патогенных и патогенных микроорганизмов, доминирующими среди которых были представители семейства Сampylobacteriaceae. Этот факт вызывает интерес, поскольку вопрос о присутствии и распространении в химусе 12-перстной кишки возбудителей инфекционных заболеваний изучен слабо.
Отметим, что полученные данные в целом соответствуют современным представлениям о микробиоте кишечника птицы (24, 36-39). Так, по результатам анализа таксономического состава около 5000 последовательностей из баз данных GenBank (https://www.ncbi.nlm.nih.gov/genbank/), Ribosomal Database Project (https://rdp.cme.msu.edu/) и Silva (https://www.arb-silva.de/) для сообщества кишечника курицы и индейки установлено присутствие представителей 13 бактериальных филумов, более 90 % которых относились к Firmicutes, Bacteroidetes и Proteobacteria (24).
В целом сравнение экологических индексов между породами птицы выявило более низкие показатели биоразнообразия для линии кур породы плимутрок, что указывает на меньшее накопление энтропии, большую степень организации и однородность состава бактериального сообщества по сравнению с таковым у породы корниш.
Сравнительный анализ бактериального сообщества содержимого 12-перстного отдела кишечника птицы позволил установить статистически значимые различия в составе микробиоты, связанные с применением в рационе птиц фитобиотика. Отмечены некоторые различия в изменении структуры микробиоценоза пищеварительного тракта в зависимости от породы птицы. Судя по индексам биоразнообразия Шеннона и доминирования Симпсона, у птиц породы корниш фитобиотик способствовал достоверному (p ≤ 0,05) снижению неоднородности состава микробиоты кишечника. По результатам таксономической оценки, в бактериальном сообществе 12-перстного отдела кишечника птицы обеих линий были отмечены существенные изменения в ответ на применение фитобиотика. Прежде всего, наблюдалось достоверное увеличение представленности членов филума Bacteroidetes,а также класса Clostridiales, включающего представителей семейств Eubacteriaceae, Clostridiaceae, Lachnospiraceae (p ≤ 0,05), что свидетельствует о повышении потенциала ферментации растительных полисахаридов микробным сообществом, поскольку перечисленные микроорганизмы обладают способностью расщеплять крахмал, клетчатку и некоторые другие углеводы, белки и дезаминировать аминокислоты.
Полученные результаты анализа состава микробиоты кишечника вполне закономерны и имеют наглядную взаимосвязь с физиологическим состоянием птиц. Так, увеличение содержания микробиоты, обладающей ферментативной активностью, согласуется с приведенными выше данными о повышении активности панкреатических ферментов в дуоденальном содержимом кишечника. Наши данные подтверждают сообщения о том, что некоторые облигатные обитатели кишечника птицы способны оказывать непосредственное влияние на продуктивность птицы. Так, V.A. Torok с соавт. (37) провели серию экспериментов, в которых обнаружили достоверную корреляцию между составом микроорганизмов в слепых отростках кишечника и эффективностью усвоения кормов. Авторы прежде всего отмечали наличие взаимосвязи для бактерий филума Firmicutes (40), включая представителей родов Eubacterium (семейство Eubacteriaceae), Roseburia (семейство Lachnospiraceae), Faecalibacterium (семейство Ruminococcaceae) (41).
Метаболизм этих микроорганизмов связан с синтезом различных летучих жирных кислот (масляной, уксусной и др.), которые необходимы птице как источник энергии. Некоторые кислоты (например, масляная) увеличивают размер эпителия кишечника (42, 43), обеспечивая тем самым барьер для токсичных агентов (44).
Отметим, что увеличение представленности ЛЖК-синтезирующих микроорганизмов в наших экспериментах при введении в рацион птиц фитобиотика положительно отражалось и на представленности в кишечнике бактерий порядка Selenomonadales,которые обладают способностью трансформировать органические кислоты до различных полезных соединений, принимая активное участие в метаболизме.
Кроме того, интересные изменения, связанные с введением в рацион птицы фитобиотика, были отмечены в отношении облигатных представителей кишечника птицы — молочнокислых бактерий родов Lactobacillus, Enterococcus, Bacillus и бифидобактерий рода Bifidobacterium, которые благодаря синтезу ими различных органических кислот и бактериоцинов способны к антагонистическому вытеснению из кишечника патогенов, таких как сальмонеллы, протеи, стафилококки, кишечная палочка, псевдомонады, стрептококки (21, 41). Установлено, что в ответ на применение в рационе фитобиотика у птиц обеих пород достоверно увеличивалась представленность бактерий родов Bacillus и Bifidobacterium (p ≤ 0,05) наряду со снижением численности других микроорганизмов, имеющих сходные свойства.
Среди бактерий, вызывающих инфекционные заболевания, у птиц детектированы возбудители кампилобактериоза (семейства Campylobacteraceae — Arcobacter, Campylobacter), пастереллеза (семейство Pasteurellaceae — Pasteurella, Haemophilus), микоплазмоза (филум Tenericutes — Mycoplasma), некротического энтерита (филум Fusobacteria), гнойно-некротических инфекций (род Staphylocoссus), клостридии (виды Clostridium novyi и C. perfringens). Большинство перечисленных микроорганизмов в сообществе кишечника птиц было минорным, за исключением кампилобактерий.
Максимальное содержание условно-патогенных бактерий выявили у птиц породы корниш, что также свидетельствует о некоторой несбалансированности микробного сообщества их кишечника (см. рис.). У кур материнской линии фиксировали меньшее присутствие бактерий родов Campylobacter, Fusobacterium, а также видов Clostridium novyi и C. perfringens. Закономерности, характеризующей сходные изменения количества патогенов в дуоденальном содержимом кишечника птицы материнской и отцовской линий в ответ на применение фитобиотика, мы не выявили. Возможно, это связано с генетическими различиями птицы и требует дополнительных исследований.
Анализируя полученные результаты, следует отметить у птиц породы корниш тенденцию к снижению представленности в дуоденальном содержимом патогенных фузобактерий — возбудителей некротического энтерита с поражением внутренних органов, суставов.
Таким образом, проведенные исследования позволяют сделать заключение, что замена кормового антибиотика эфирными маслами растений в комбикормах молодняка мясных кур (исходные линии Б5 и Б9) позволяет получить практически одинаковую живую массу птицы. Развитие репродуктивных органов самцов (семенников) и самок (яичники и яйцеводы) в обеих группах оставалось в пределах нормы без существенных различий. Отмечено увеличение активности липазы в химусе 12-перстной кишки на 30,9 % у линии Б5 и на 98,3 % — у линии Б9, а также протеаз у птицы линии Б5 на 36,4 %, что согласуется с данными по переваримости жира и азота корма в этих группах. Результаты проведенных исследований свидетельствуют о наличии у кур обеих пород заметных изменений в бактериальном сообществе дуоденального содержимого, связанных прежде всего с повышением доли представителей микроорганизмов, обладающих ферментативными свойствами в отношении сложных полисахаридов (класс Clostridia, филум Bacteroidetes), а также бактерий с высокой антагонистической активностью (Bifidobacterium, Bacillus и др.).
1. Живая масса (г) молодняка мясных кур (Gallus gallus domesticus) двух линий и потребление корма (г·гол.-1·сут-1) при включении в рацион кормового антибиотика Бацитрацина-30 или фитобиотика на основе эфирных масел Интебио в зависимости от возраста (M±SEM, СГЦ «Загорское ЭПХ», г. Сергиев Посад, Московская обл., 2017 год) |
||||||||||
Возраст, |
Линия |
|||||||||
Б5 |
Б9 |
|||||||||
♂ |
♀ |
ПК |
♂ |
♀ |
ПК |
|||||
К |
О |
К |
О |
К |
О |
К |
О |
|||
1 |
229±5,4 |
219±6,2 |
220±4,8 |
215±4,3 |
266 |
209±3,8 |
201±5,0 |
213±2,9 |
207±4,5 |
259 |
2 |
340±6,3 |
335±6,5 |
332±5,9 |
327±5,8 |
294 |
187±6,0 |
180±6,5 |
270±5,6 |
260±4,8 |
280 |
3 |
630±8,6 |
629±8,8 |
605±7,8 |
600±6,9 |
350 |
527±7,5 |
521±6,8 |
492±6,6 |
489±7,2 |
336 |
4 |
780±16,3 |
775±15,8 |
700±15,2 |
689±16,0 |
434 |
679±12,5 |
667±13,4 |
605±10,6 |
600±11,3 |
420 |
5 |
938±20,3 |
940±21,8 |
807±18,8 |
800±18,1 |
469 |
849±18,6 |
835±20,1 |
718±17,7 |
710±16,5 |
455 |
6 |
1110±21,6 |
1115±20,8 |
980±17,6 |
982±18,0 |
476 |
992±19,3 |
990±18,4 |
850±17,0 |
842±16,8 |
462 |
7 |
1260±22,3 |
1262±222,1 |
1005±118,4 |
1001±18,6 |
504 |
1222±20,3 |
1215±18,6 |
1039±16,6 |
1030±16,8 |
490 |
8 |
1450±23,6 |
1440±24,0 |
1170±20,7 |
1162±20,1 |
511 |
1390±21,4 |
1382±19,9 |
1127±18,8 |
1122±19,2 |
497 |
9 |
1595±25,2 |
1589±24,8 |
1245±23,3 |
1240±22,9 |
518 |
1450±22,6 |
1455±23,1 |
1200±20,4 |
1207±21,1 |
504 |
10 |
1790±28,3 |
1794±26,9 |
1440±27,9 |
1437±26,8 |
525 |
1590±27,6 |
1587±28,0 |
1295±25,2 |
1290±26,4 |
511 |
11 |
1900±30,2 |
1910±31,0 |
1550±28,3 |
1555±27,8 |
546 |
1605±28,8 |
1600±30,3 |
1375±25,5 |
1367±24,8 |
532 |
12 |
2020±32,3 |
2015±30,8 |
1700±26,6 |
1692±27,0 |
553 |
1810±25,6 |
1812±26,1 |
1460±24,4 |
1462±23,7 |
539 |
13 |
2110±30,6 |
2117±31,2 |
1740±27,6 |
1747±26,5 |
560 |
1890±28,0 |
1884±27,4 |
1530±22,6 |
1525±23,1 |
546 |
14 |
2275±31,2 |
2269±29,9 |
1875±28,3 |
1880±27,6 |
574 |
1940±25,6 |
1932±24,8 |
1600±20,7 |
1592±21,4 |
560 |
15 |
2495±28,8 |
2490±28,0 |
1910±26,4 |
1905±25,9 |
602 |
1995±27,7 |
1990±26,3 |
1687±24,4 |
1680±22,6 |
588 |
16 |
2530±32,4 |
2537±29,9 |
1947±27,7 |
1951±26,8 |
623 |
2140±28,5 |
2147±29,6 |
1710±26.6 |
1712±25,7 |
609 |
17 |
2650±31,7 |
2649±28,8 |
2005±30,3 |
2001±30,0 |
630 |
2267±30,1 |
2260±31,2 |
1775±28,0 |
1771±27,8 |
616 |
18 |
2795±32,5 |
2790±28,4 |
2190±29,6 |
2185±27,7 |
658 |
2368±32,5 |
2351±33,0 |
1804±26,5 |
1801±24,7 |
644 |
10 |
2940±30,8 |
2947±31,1 |
2210±29,6 |
2215±29,1 |
714 |
2478±33,4 |
2480±34,6 |
1843±30,2 |
1845±28,8 |
672 |
20 |
3075±32,6 |
3077±33,2 |
2235±28,8 |
2240±29,0 |
728 |
2505±36,2 |
2500±35,8 |
1885±27,7 |
1890±27,0 |
728 |
21 |
3169±35,0 |
3172±35,7 |
2316±30,9 |
2318±30,7 |
770 |
2589±38,8 |
2590±39,2 |
1920±29,8 |
1917±28,7 |
756 |
Примечание. К и О — соответственно контроль и опыт, ПК — потребление корма за 1 нед. Описание групп см. в разделе «Методика». |
||||||||||
2. Ферментативная активность дуоденального химуса и активность панкреатических ферментов в плазме крови у мясных кур (Gallus gallus domesticus) двух линий при включении в рацион кормового антибиотика Бацитрацина-30 или фитобиотика на основе эфирных масел Интебио(M±SEM, СГЦ «Загорское ЭПХ», г. Сергиев Посад, Московская обл., 2017 год) |
|||
Показатель |
Группа |
К контролю, % |
|
контроль (n = 5) |
опыт (n = 5) |
||
Линия Б5 |
|||
Активность ферментов в химусе |
|||
Амилаза, мг·мл-1·мин-1 |
219±21,1 |
231±25,5 |
105,5 |
Липаза, Ед/л |
750±54,7 |
982±76,5* |
130,9 |
Протеазы, мг·мл-1·мин-1 |
22±1,8 |
30±1,7* |
136,4 |
Активность ферментов в плазме крови |
|||
Амилаза, Ед/л |
395±43,5 |
322±20,5 |
81,5 |
Липаза, Ед/л |
29±2,2 |
28±2,1 |
96,5 |
Трипсин, Ед/л |
35±5,4 |
34±3,5 |
97,1 |
Линия Б9 |
|||
Активность ферментов в химусе |
|||
Амилаза, мг·мл-1·мин-1 |
266±31,0 |
305±41,0 |
114,7 |
Липаза, Ед/л |
301±37,5 |
597±50,3** |
198,3 |
Протеазы, мг·мл-1·мин-1 |
36±0,8 |
36±1,0 |
100,0 |
Активность ферментов в плазме крови |
|||
Амилаза, Ед/л |
290±25,1 |
263±6,5 |
90,7 |
Липаза, Ед/л |
15±0,9 |
19±0,6* |
126,7 |
Трипсин, Ед/л |
29±0,5 |
30±0,9 |
103,4 |
Примечание. Описание групп см. в разделе «Методика». |
|||
3. Индексы биоразнообразия бактериального сообщества в содержимом 12-перстной кишки у двух линий мясных кур (Gallus gallus domesticus) при включении в рацион кормового антибиотика Бацитрацина-30 или фитобиотика на основе эфирных масел Интебио(M±SEM, СГЦ «Загорское ЭПХ», г. Сергиев Посад, Московская обл., 2017 год) |
||||
Показатель |
Линия Б9 |
Линия Б9 |
||
контроль |
опыт |
контроль |
опыт |
|
Индекс биоразнообразия Шеннона (Н) |
2,53±0,11 |
2,43±0,09 |
3,39±0,15 |
1,67±0,07 |
Индекс биоразнообразия Симпсона (D) |
0,82±0,04 |
0,82±0,06 |
0,91±0,02 |
0,68±0,08 |
Число филотипов |
149,05±5,23 |
130,82±3,68 |
125,65±3,12 |
170,36±6,09 |
Примечание. Описание групп см. в разделе «Методика». |
||||
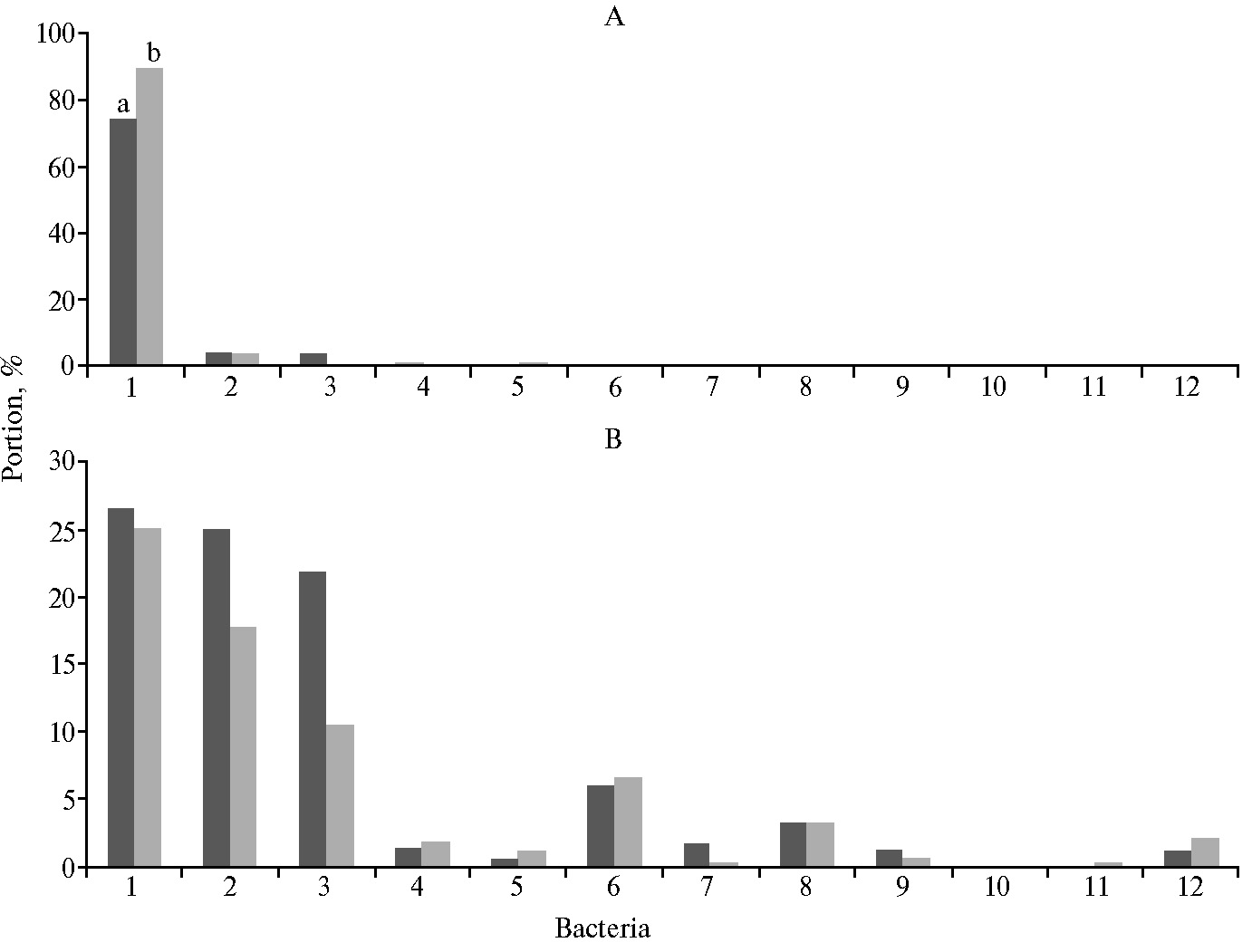
Представленность разных групп бактерий в содержимом 12-перстной кишки у родительских линий цыплят-бройлеров (Gallus gallus domesticus) Б9 (а) и Б5 (б) в контрольных (А) и опытных (Б) группах: 1 — порядок Lactobacillales, 2 — неидентифицируемые бактерии, 3 — род Bacil-lus, 4 — филум Actinobacteria, 5 — семейство Campylobacteriaceae, 6 — филум Bacteroidetes, 7 — класс Clostridiales, 8 — порядок Selenomonadales, 9 — род Bifidobacterum, 10 — филум Fusobacteria, 11 — род Staphylococcus, 12 — порядок Pseudomonadales. Описание групп см. в разделе «Методика».
ЛИТЕРАТУРА
- Фисинин В.И., Егоров И.А., Лаптев Г.Ю., Ленкова Т.Н., Никонов И.Н., Ильина Л.А., Манукян В.А., Грозина А.А., Егорова Т.А., Новикова Н.И., Йылдырым Е.А. Получение продукции птицеводства без антибиотиков с использованием перспективных программ кормления на основе пробиотических препаратов. Вопросы питания, 2017, 86(6): 114-124 (doi: 10.24411/0042-8833-2017-00013).
- Константинов В. Органические кислоты — отличный результат. Комбикорма, 2010, 6: 115-116.
- Smith J.A. The future of poultry production in the USA without antibiotics. Poultry International, 2002, 41: 68-69.
- Фисинин В.И., Околелова Т.М., Просвирякова О.А., Андрианова Е.Н., Крюков О.В., Кузовникова А., Нармонтиене М., Бевзюк В.Н., Папазян Т.Т., Тардатьян А.Г., Щукина С.А., Косинцев Ю.В., Тимофева Э.Н. Органическиекислотыиподкислителивкомбикормахдляптицы. Сергиев-Посад, 2008.
- Джавадов Э.Д., Дмитриева М.Е., Трефилов Б.Б, Новикова О.Б., Титова Т.Г. Инфекционная патология в промышленном птицеводстве. Ветеринария и кормление, 2016, 2: 24-27.
- Collier C.T., Hofacre C.L., Payne A.M., Anderson D.B., Kaiser P., Mackie R.I., Gaskins H.R. Coccidia induced mucogenesis promotes the onset of necrotic enteritis by supporting Clostridium perfringens growth. Veterinary Immunology and Immunopathology, 2008, 122(1-2): 104-115 (doi: 10.1016/j.vetimm.2007.10.014).
- Hermans D., Pasmans F., Messens W., Martel A., Van Immerseel F., Rasschaert G., Heyndrickx M., Van Deun K., Haesebrouck F. Poultry as a host for the zoonotic pathogen Campylobacter jejuni. Vector-Borne and Zoonotic Disease, 2012, 12(2): 89-98 (doi: 10.1089/vbz.2011.0676).
- Seys S.A., Sampedro F., Hedberg C.W. Assessment of meat and poultry product recalls due to Salmonella contamination: product recovery and illness prevention. Journal of Food Protection, 2017, 80(8): 1288-1292 (doi: 10.4315/0362-028X.JFP-16-424).
- Hashemi S.R., Zukifli I., Hair Bejo M., Farida A., Somchit M.N. Acute toxicity study and phytochemical screening of selected herbal aqueous extract in broiler chickens. International Journal of Pharmacology, 2008, 4(5): 352-360 (doi: 10.3923/ijp.2008.352.360).
- Крюков В.С., Глебова И.В. Антибактериальное действие эфирных масел лекарственных растений (обзор). Проблемы биологии продуктивных животных, 2017, 3: 5-25.
- Nazzaro F., Frantianni F., De Marino L., Coppala R., De Feo V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals, 2013, 6(12): 1451-1474 (doi: 10.3390/ph6121451).
- Christaki E., Bonos E., Giannenas I., Florou-Paneri P. Aromatic plants as a source of bioactive comppouds. Agriculture, 2012, 2(3): 228-243 (doi: 10.3390/agriculture2030228).
- Zeng Z., Zhang S., Wang H., Piao X. Essential oil and aromatic plants as feed additives in non-ruminant nutrition: a review. Journal of Animal Science and Biotechnology, 2015, 6(1): Article 7 (doi: 10.1186/s40104-015-0004-5).
- Odoemalam V.U., Etuk I.F., Ndelekwute E.K., Iwuji T.C., Ekwe Ch.C. Herbs and spices: option for sustainable animal production. Journal of Biology, Agriculture and Healthcare, 2013, 3(7): 116-124.
- Costa L.B., Luciano F.B., Miyada V.S., Gois F.D. Herbal extracts and organic acids as natural feed additives in pig diets. South African Journal of Animal Science, 2013, 43(2): 181-193 (doi: 10.4314/sajas.v43i2.9).
- Егорова А.В. Мясные куры родительского стада: оценка, отбор и подбор птицы. Птицеводство, 2012, 12: 8-10.
- Егорова А.В., Тучемский Л.И., Емануйлова Ж.В. Продуктивность родительских форм мясных кур селекции селекционного-генетического центра «Смена». Зоотехния, 2015, 6: 2-4.
- Yeoman C.J., Chia N., Jeraldo P., Sipos M., Goldenfeld N.D., White B.A. The microbiome of the chicken gastrointestinal tract. Animal Health Research Reviews, 2012, 13(1): 89-99 (doi: 10.1017/S1466252312000138).
- Kerr A.K., Farrar A.M., Waddell L.A., Wilkins W., Wilhelm B.J., Bucher O., Wills R.W., Bailey R.H., Varga C., McEwe S.A., Rajić A. A systematic review-meta-analysis and meta-regression on the effect of selected competitive exclusion products on Salmonella spp. prevalence and concentration in broiler chickens. Preventive Veterinary Medicine, 2013, 111(1-2): 112-125 (doi: 10.1016/j.prevetmed.2013.04.005).
- Dobson A., Cotter P.D., Ross R.P., Hill C. Bacteriocin production: a probiotic trait? Applied and Environmental Microbiology, 2012, 78(1): 1-6 (doi: 10.1128/AEM.05576-11).
- Messaoudi S., Kergourlay G., Dalgalarrondo M., Choiset Y., Ferchichi M., Prévost H., Pilet M.F., Chobert J.M., Manai M., Dousset X. Purification and characterization of a new bacteriocin active against Campylobacter produced by Lactobacillus salivarius SMXD51. Food Microbiology, 2012, 32(1): 129-134 (doi: 10.1016/j.fm.2012.05.002).
- Sekirov I., Russell S.L., Antunes L.C.M., Finlay B.B. Gut microbiota in health and disease. Physiol. Rev., 2010, 90(3): 859-904 (doi: 10.1152/physrev.00045.2009).
- Sun H., Tang J.W., Fang C.L., Yao X.H., Wu Y.F., Wang X., Feng J. Molecular analysis of intestinal bacterial microbiota of broiler chickens fed diets containing fermented cottonseed meal. Poultry Science, 2013, 92(2): 392-401 (doi: 10.3382/ps.2012-02533).
- Wei S., Morrison M., Yu Z. Bacterial census of poultry intestinal microbiome. Poultry Science, 2013, 92(3): 671-683 (doi: 10.3382/ps.2012-02822).
- Stanley D., Geier M.S., Chen H., Hughes R.J., Moore R.J. Comparison of fecal and cecal microbiotas reveals qualitative similarities but quantitative differences. BMC Microbiology, 2015, 15(1): Article 51 (doi: 10.1186/s12866-015-0388-6).
- LeBlanc J.G., Milani C., de Giori G.S., Sesma F., van Sinderen D., Ventura M. Bacteria as vitamin suppliers to their host: a gut microbiota perspective. Curr. Opin. Biotechnol., 2013, 24(2): 160-168 (doi: 10.1016/j.copbio.2012.08.005).
- Fasina Y.O., Hoerr F.J., McKee S.R., Conner D.E. Influence of Salmonella enterica serovar Typhimurium infection on intestinal goblet cells and villous morphology in broiler chicks. Avian Diseases, 2010, 54(2): 841-847 (doi: 10.1637/9055-090809-Reg.1).
- Chae B., Ingale S., Kim J., Kim K., Sen S., Lee S., Khong C., Kim E.K., Kwon I.K. Effect of dietary supplementation of probiotics on performance, caecal microbiology and small intestinal morphology of broiler chickens. Animal Nutrition and Feed Technology, 2012, 12(1): 1-12.
- Liao N., Yin Y., Sun G., Xiang C., Liu D., Yu H.D., Wang X. Colonization and distribution of segmented filamentous bacteria (SFB) in chicken gastrointestinal tract and their relationship with host immunity. FEMS Microbiology Ecology, 2012, 81(2): 395-406 (doi: 10.1111/j.1574-6941.2012.01362.x).
- Sekelja M., Rud I., Knutsen S.H., Denstadli V., Westereng B., Naes T., Rudi K. Abrupt temporal fluctuations in the chicken fecal microbiota are explained by its gastrointestinal origin. Applied and Environmental Microbiology, 2012, 78(8): 2941-2948 (doi: 10.1128/AEM.05391-11).
- Мерина-Глузкина В.М. Сравнительная оценка сахарифицирующего и декстринирующего методов при определении активности амилазы крови здоровых и больных острым панкреатитом. Лабораторное дело, 1965, 3: 142-146.
- Батоев Ц.Ж. Фотометрическое определение активности протеолитических ферментов в поджелудочной железе, соке по уменьшению концентрации казеина. Сб. науч. тр. Бурятского СХИ (Улан-Удэ), 1971, 25: 122-126.
- Инструкции по санитарно-микробиологическому контролю тушек, мяса птицы, птицепродуктов, яиц и яйцепродуктов на птицеводческих и перерабатывающих предприятиях. М., 1990.
- Уилсон К., Уолкер Дж. Принципы и методы биохимии и молекулярной биологии. М., 2015.
- Mikhailova A.G., Khairullin R.F., Demidyuk I.V., Kostrov S.V., Grinberg N.V., Burova T.V., Grinberg V.Y., Rumsh L.D. Cloning, sequencing, expression, and characterization of thermostability of oligopeptidase B from Serratia proteamaculans, a novel psychrophilic protease. Protein Expression and Purification, 2014, 93: 63-76 (doi: 10.1016/j.pep.2013.10.011).
- Егоров И.А., Манукян В.А., Ленкова Т.Н., Околелова Т.М., Лукашенко В.С., Шевяко А.Н., Игнатова Г.В., Егорова Т.В., Андрианова Е.Н., Розанов Б.Л., Лысенко М.А., Егорова Т.А., Грозина А.А., Лаптев Г.Ю., Никонов И.Н., Александрова И.Л., Ильина Л.А., Новикова Н.И., Фисинин В.И. Методика проведения научных и производственных исследований по кормлению сельскохозяйственной птицы. Молекулярно-генетические методы определения микрофлоры кишечника. Сергиев Посад, 2013.
- Torok V.A., Hughes R.J., Mikkelsen L.L., Perez-Maldonado R., Balding K., MacAlpine R., Percy N.J., Ophel-Keller K. Identification and characterization of potential performance-related gut microbiota in broiler chickens across various feeding trials. Applied and Environmental Microbiology, 2011, 77(17): 5868-5878 (doi: 10.1128/AEM.00165-11).
- Diaz-Sanchez S., Hanning I., Pendleton S., D’Souza D. Next-generation sequencing: the future of molecular genetics in poultry production and food safety. Poultry Science, 2013, 92(2): 562-572 (doi: 10.3382/ps.2012-02741).
- Choi J.H., Kim G.B., Cha C.J. Spatial heterogeneity and stability of bacterial community in the gastrointestinal tracts of broiler chickens. Poultry Science, 2014, 93(8): 1942-1950 (doi: 10.3382/ps.2014-03974).
- Bjerrum L., Engberg R.M., Leser T.D., Jensen B.B., Finster K., Pedersen K. Microbial community composition of the ileum and cecum of broiler chickens as revealed by molecular and cellular-based techniques. Poultry Science, 2006, 85(7): 1151-1164 doi: 10.1093/ps/85.7.1151).
- Louis P., Young P., Holtrop G., Flint H.J. Diversity of human colonic butyrate-producing bacteria revealed by analysis of the butyryl-CoA:acetate CoA-transferase gene. Environmental Microbiology, 2010, 12(2): 304-314 (doi: 10.1111/j.1462-2920.2009.02066.x).
- Le Blay G., Blottiere H.M., Ferrier L., Le Foli E.C., Bonnet J.P., Galmiche C., Cherbut C. Short-chain fatty acids induce cytoskeletal and extracellular protein modifications associated with modulation of proliferation on primary culture of rat intestinal smooth muscle cells. Dig. Dis. Sci., 2000, 45(8): 1623-1630 (doi: 10.1023/a:1005529414765).
- Fukunaga T., Sasaki M., Araki Y., Okamoto T., Yasuoka T., Tsujikawa T., Fujiyama Y., Bamba T. Effects of the soluble fibre pectin on intestinal cell proliferation, fecal short chain fatty acid production and microbial population. Digestion, 2003, 67(1-2): 42-49 (doi: 10.1159/000069705).
- Niba A.T., Beal J.D., Kudi A.C., Brooks P.H. Bacterial fermentation in the gastrointestinal tract of non-ruminants: influence of fermented feeds and fermentable carbohydrates. Tropical Animal Health and Production, 2009, 41(7): 1393-1407 (doi: 10.1007/s11250-009-9327-6).