








УДК 636.086.2/.3:581.132:57.04
ТЕМПЕРАТУРА ПОЧВЫ И СО2-ОБМЕН У РАСТЕНИЙ ЛЮПИНА
С.Н. ДРОЗДОВ, Е.С. ХОЛОПЦЕВА, Э.Г. ПОПОВ
Изучали влияния температуры почвы на СО2-обмен у растений контрастных по содержанию алкалоидов сортов люпина узколистного (Lupinus angustifolius L.) в зависимости от температуры воздуха и освещенности.
Ключевые слова: люпин узколистный (сорта Сидерат 38, Кристалл), температура воздуха и почвы, освещенность, СО2-обмен у растений, планируемый многофакторный эксперимент.
Большая часть территории России (66,7 %) расположена в районах Крайнего Севера или в местностях, приравненных к ним, где одним из основных лимитирующих факторов для растений в вегетационный период служат низкие температуры почвы (1, 2). Поэтому важную роль играет подбор и селекция культур с целью получения сортов, способных давать хорошие урожаи в соответствующих почвенно-климатических условиях (3) при низкой ресурсо- и энергоемкости возделывания и усилении средообразующей (в том числе ресурсовосстанавливающей, почвозащитной и почвоулучшающей) функции (4).
С учетом острого дефицита белка в кормопроизводстве и азота в почвах Нечерноземья значительный интерес для северных регионов представляют бобовые, в частности люпин узколистный. Его высокая продуктивность связана с большой усваивающей способностью корневой системы, активной фиксацией атмосферного азота и быстрым формированием высокого урожая зеленой массы (5). Успешная интродукция люпина в этих регионах сдерживается недостаточной изученностью экофизиологических характеристик его сортов (6), в том числе влияния температуры почвы на СО2-обмен у растений (7).
Задачей нашей работы было изучение влияния температуры почвы на СО2-обмен у растений контрастных по содержанию алкалоидов сортов люпина узколистного (Lupinus angustifolius L.) в зависимости от температуры воздуха и освещенности.
Методика. Исследования проводили в Карельском научном центре РАН в 2004-2006 годах на растениях люпина узколистного двух сортов — наиболее скороспелого, высокобелкового, малоалкалоидного, заморозкоустойчивого сорта Кристалл и скороспелого, высокопродуктивного, алкалоидного, устойчивого к растрескиванию бобов и осыпанию семян сорта Сидерат 38 сидерального типа использования. Семена изучаемых сортов (по 8-10 шт.) высевали в сосуды с песком и выращивали при ежедневном поливе питательным раствором Кнопа с добавлением микроэлементов в факторостатных условиях — 14-часовой световой день, освещенность около 10 клк, температура воздуха 22/20 °С (день/ночь). Изучение влияния света и температуры на СО2-обмен интактных растений проводили в фазу 6-7 листьев: сосуды с хорошо развитыми растениями помещали в фитотрон с возможностью регулировать освещенность в диапазоне 0-40 клк при спектре, близком к естественному, и температуре воздуха от +5 до +40 °С. Сигналы датчиков температуры, освещенности и показания газоанализатора регистрировали самопишущим потенциометром КСП-4 (Россия) c точностью измерения 0,5 % (8). Эксперимент проводили по 21-31-точечному плану 2-го порядка (9) с продолжительностью воздействия факторов в каждой точке плана 40-50 мин. Газообмен определяли по разности содержания СО2 в воздухе на входе и выходе ассимиляционной камеры с помощью включенного по дифференциальной схеме газоанализатора Infralyt-4 («Junkalor», Германия) с пределами измерения 0-0,01 об% СО2; показатели пересчитывали на единицу сухой массы интактных растений. Для построения уравнения зависимости нетто-фотосинтеза от исследуемых факторов использовали регрессионный анализ (8).
Результаты. План трехфакторного эксперимента и соответствующие показатели нетто-фотосинтеза растений приведены в таблице 1.
| 1. План трехфакторного эксперимента и показатели нетто-фотосинтеза у интактных растений двух сортов люпина в фазу 6-7 листьев в регулируемых условиях среды по 21-31-точечному плану 2-го порядка | ||||||||
Точка |
E, клк |
Т 1, °С |
Т 2, °С |
PN, мг/(г·ч) |
Е, клк |
Т 1, °С |
Т 2, °С |
PN, мг/(г·ч) |
С о р т С и д е р а т 38 |
С о р т К р и с т а л л |
|||||||
1 |
25 |
17 |
17 |
11,132 |
20 |
5 |
5 |
7,869 |
2 |
10 |
27 |
17 |
2,796 |
10 |
15 |
7 |
2,098 |
3 |
40 |
7 |
24 |
13,385 |
20 |
30 |
14 |
2,623 |
4 |
40 |
27 |
10 |
11,071 |
30 |
15 |
7 |
13,37 |
5 |
40 |
7 |
10 |
15,124 |
30 |
8 |
15 |
12,59 |
6 |
40 |
27 |
24 |
10,635 |
20 |
15 |
15 |
9,181 |
7 |
10 |
17 |
24 |
3,509 |
10 |
6 |
19 |
5,508 |
8 |
25 |
7 |
24 |
10,635 |
30 |
7 |
23 |
12,06 |
9 |
25 |
17 |
17 |
11,467 |
10 |
30 |
25 |
0,524 |
10 |
10 |
7 |
17 |
6,197 |
30 |
30 |
26 |
9,181 |
11 |
10 |
17 |
10 |
5,583 |
10 |
14 |
5 |
5,511 |
12 |
20 |
5 |
5 |
7,870 |
20 |
28 |
9 |
5,118 |
13 |
10 |
15 |
7 |
2,099 |
30 |
14 |
6 |
13,97 |
14 |
20 |
30 |
14 |
2,623 |
30 |
8 |
17 |
11,02 |
15 |
30 |
15 |
7 |
13,379 |
20 |
15 |
16 |
8,070 |
16 |
30 |
8 |
15 |
12,592 |
10 |
5 |
23 |
1,968 |
17 |
20 |
15 |
15 |
9,182 |
30 |
7 |
25 |
10,03 |
18 |
10 |
6 |
19 |
5,509 |
20 |
14 |
15 |
8,267 |
19 |
30 |
7 |
23 |
12,067 |
10 |
28 |
30 |
1,181 |
20 |
10 |
30 |
25 |
0,525 |
30 |
29 |
26 |
8,858 |
21 |
30 |
30 |
26 |
9,182 |
25 |
17 |
17 |
11,13 |
22 |
– |
– |
– |
– |
10 |
27 |
17 |
2,796 |
23 |
– |
– |
– |
– |
40 |
7 |
24 |
13,38 |
24 |
– |
– |
– |
– |
40 |
27 |
10 |
11,07 |
25 |
– |
– |
– |
– |
40 |
7 |
10 |
15,12 |
26 |
– |
– |
– |
– |
40 |
27 |
24 |
10,63 |
27 |
– |
– |
– |
– |
10 |
17 |
24 |
3,509 |
28 |
– |
– |
– |
– |
25 |
7 |
24 |
10,63 |
29 |
– |
– |
– |
– |
25 |
17 |
17 |
11,46 |
30 |
– |
– |
– |
– |
10 |
7 |
17 |
6,197 |
31 |
– |
– |
– |
– |
10 |
17 |
10 |
5,583 |
П р и м е ч а н и е. PN — интенсивность нетто-фотосинтеза, Е — освещенность, Т 1 — температура воздуха, Т 2 — температура почвы. Прочерк означает, что эксперимент проведен по 21-точечному плану 2-го порядка. |
||||||||
По результатам регрессионного анализа, исследуемые сорта светолюбивы, холодоустойчивы и сходны по возможному потенциальному максимуму нетто-фотосинтеза в пересчете на единицу сухой массы интактных
| 2. Условия потенциально возможного максимума и оптимум нетто-фотосинтеза у растений двух сортов люпина узколистного в фазу 6-7 листьев | ||||||||
Сорт |
Максимум |
Оптимум |
||||||
нетто-фотоси-нтез, мг/(г·ч) |
освеще-нность, клк |
температура,°С |
нетто-фотоси-нтез, мг/(г·ч) |
освещен-ность, клк |
температура, °С |
|||
воздуха |
почвы |
воздуха |
почвы |
|||||
Кристалл |
18,3 |
49,0 |
12,6 |
6,0 |
17,4 |
41,0-55,0 |
1,0-24,5 |
5,0-19,0 |
Сидерат 38 |
17,0 |
55,0 |
9,5 |
10,8 |
15,4 |
37,0-56,0 |
1,0-19,0 |
3,5-20,4 |
растений при естественном содержании СО2 в воздухе, но значительно различаются по зависимости от свето-температурных условий, обеспечивающих его достижение (табл. 2). По этим показателям растения сорта Кристалл менее светолюбивы, чем растения сорта Сидерат 38, и оптимум нетто-фотосинтеза у них возможен при более узком диапазоне температуры почвы и более широком диапазоне температуры воздуха.
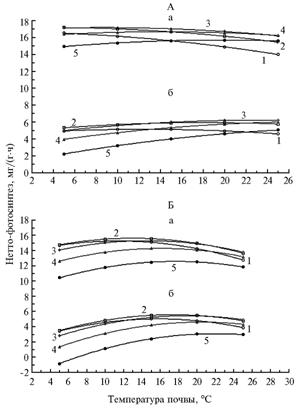 |
Влияние температуры почвы на нетто-фотосинтез интактных растений люпина узколистного сорта Кристалл (А) и Сидерат 38 (Б) в фазу 6-7 листьев в зависимости от освещенности (а — 40 клк, б — 10 клк) и температуры воздуха (1-5 — соответственно 5, 10, 15, 20 и 25 °С). |
В естественных условиях сочетание интенсивности факторов, обеспечивающих достижение потенциального максимума видимого фотосинтеза, наблюдается весьма редко, поэтому успешное вегетирование определяется сочетанием интенсивности факторов, обеспечивающих оптимальные условия произрастания, то есть фоновой зоной, в которой, по данным литературы, интенсивность нетто-фотосинтеза составляет не менее 90 % от потенциального максимума (10). При колебаниях условий среды в пределах этого диапазона терморезистентность растений не изменяется. За пределами фоновой зоны расположены зоны закаливания и повреждения (11). Границы зон определяются геномом и зависят от вида, сорта, фазы развития растений и условий среды.
Изучение влияния температуры почвы на нетто-фотосинтез интактных растений люпина показало, что оно зависит от сорта, освещенности и температуры воздуха (рис.). Потенциальный максимум видимого фотосинтеза при наиболее благоприятном сочетании освещенности и температуры воздуха у растений сорта Кристалл наступал при температуре почвы 6,0 °С и воздуха 12,6 °С (табл. 2). В то же время при более низкой освещенности (около 10 клк) максимум интенсивности нетто-фотосинтеза (см. рис. А, б) достигался при более высокой температуре почвы (примерно 25 °С) и воздуха (15 °С). Максимум видимого фотосинтеза по мере снижения температуры воздуха при высокой освещенности смещался в сторону пониженных, при низкой — более высоких почвенных температур. У растений сорта Сидерат 38 потенциальный максимум нетто-фото-синтеза при высокой освещенности регистрировали при температуре почвы около 13 °С (см. рис. Б, а), при низкой — около 18 °С (см. рис. Б, б).
При дальнейшем повышении температуры воздуха снижение интенсивности нетто-фотосинтеза после достижения максимума связано, в основном, с возрастанием величины составляющей поддержания в дыхании, особенно при переходе в зону теплового закаливания.
Учитывая, что фотосинтез — один из процессов, наиболее отзывчивых на влияние внешних факторов, правомерно рассматривать условия, обеспечивающие достижение оптимума нетто-фотосинтеза, как условия оптимума жизнедеятельности в исследуемую фазу развития конкретного генотипа.
Таким образом, влияние температуры почвы на нетто-фотосинтез у люпина узколистного зависит от сорта и сопутствующих условий окружающей среды. Исследованные сорта светолюбивы и холодоустойчивы: потенциальный максимум нетто-фотосинтеза у растений достигается при высокой освещенности и относительно низких температурах воздуха и почвы, а оптимум — в широком диапазоне значений этих факторов среды.
Л И Т Е Р А Т У Р А
1. Д а д ы к и н В.П. Особенности поведения растений на холодных почвах. М., 1952.
2. К о р о в и н А.И. Температура почвы и растения на Севере. Петрозаводск, 1961.
3. Р о м а н е н к о Г.А. Сельскохозяйственная наука в России в 21-м веке. Вест. РАСХН, 1994, 2: 3-7.
4. Ж у ч е н к о А.А. Научные приоритеты развития растениеводства в XXI веке. Вест. РАСХН, 2000, 2: 9-13.
5. А л е к с е е в Е.К. Слабоалкалоидные люпины как кормовая культура. В сб.: Кормовые люпины. М., 1953: 5-48.
6. Х о л о п ц е в а Е.С., Д р о з д о в С.Н. Нетто-фотосинтез растений как показатель экологической характеристики биоразнообразия. С.-х. биол., 2008, 3: 106-109.
7. К у р е ц В.К., Д р о з д о в С.Н., П о п о в Э.Г., Т а л а н о в А.В., Д е м б о Е.Д. Температурный градиент воздух—почва как фактор оптимизации нетто-фотосинтеза интактных растений. Физиол. раст., 2003, 50(1): 81-87.
8. К у р е ц В.К., П о п о в Э.Г. Статистическое моделирование системы связей растение—среда. Л., 2001.
9. Г о л и к о в а Т.И., П а н ч е н к о Л.А., Ф р и д м а н А.Ф. Каталог планов второго порядка. Ч. 1, вып. 47. М., 1974.
10. Л а р х е р В. Экология растений. М., 1978.
11. Д р о з д о в С.Н., К у р е ц В.К. Некоторые аспекты экологической физиологии растений. Петрозаводск, 2003.
SOIL TEMPERATURE AND СО2-METABOLISM IN LUPINE PLANTS
S.N. Drozdov, E.S. Kholoptseva, E.G. Popov
The authors studied the influence of soil temperature on СО2-metabolism in lupine varieties (Lupinus angustifolius L.), which are contrasting on alkaloids content, in connection with air temperature and illumination. The optimal net photosynthesis was achieved in wide rang of these parameters.
Key words: Lupinus angustifolius (var. Siderat 38, var. Kristall), air and soil temperatures, light intensity, CO2-exchang, preplanned multifactor experiment.
Учреждение Российской академии наук Институт биологии Карельского научного центра РАН, |
Поступила в редакцию |




