








УДК 636.32/.38:575.17
ДИНАМИКА ГЕНЕТИЧЕСКОЙ СТРУКТУРЫ ОВЕЦ МЯСОШЕРСТНОЙ ПОРОДЫ СИБИРСКОГО ТИПА, СОЗДАННОЙ С ИСПОЛЬЗОВАНИЕМ ВОСПРОИЗВОДИТЕЛЬНЫХ МНОГОПЛОДНЫХ СКРЕЩИВАНИЙ
С.В. ПАВЛЕНКО, Е.К. МИНИНА, А.В. КУШНИР, В.И. ГЛАЗКО
Выполняли анализ генетической структуры поколений овец мясошерстной породы с использованием различных молекулярно-генетических маркеров для оценки ее консолидированности. Обнаружены локус-специфические особенности генетических взаимоотношений между поколениями новой породы и исходными породами.
Ключевые слова: молекулярно-генетические маркеры, полиморфизм, генетические расстояния, многопородные скрещивания, породы овец.
Создание новой породы всегда связано с преобразованием исходного генетического материала под влиянием традиционных методов селекции — отбора, подбора животных для скрещиваний и формирования групп с желательным сочетанием хозяйственно ценных признаков (1-4). Применение молекулярно-генетических маркеров для описания генетической структуры популяции на ранних этапах породообразовательного процесса может способствовать выявлению сочетаний генотипов, ассоциированных с контролируемым комплексом фенотипических характеристик, позволяет оценивать изменения генетической структуры и их направленность при воспроизводстве новой породы в ряду поколений, контролировать степень консолидиро-ванности породы и особенности генетических взаимоотношений с той или иной исходной породой (1, 5).
В задачу настоящего исследования входила оценка внутрипородной генетической гетерогенности в поколениях для определения консолидированности сибирского типа советской мясошерстной породы овец, полученного при сложных воспроизводительных скрещиваниях между породами алтайская, линкольн и ромни-марш, и сравнение этих внутрипородных групп с исходными породами (2).
Методика. Объектом исследований служили четыре группы кроссбредных овец сибирской мясошерстной породы (по 27-30 гол. в каждой), воспроизводство которых осуществлялось в совхозе «Медведский» (Новосибирская обл.): I, II и III группы — матки соответственно 1997, 1998 и 2000 года рождения, IV — молодняк (баранчики и ярочки) 2002 года рождения; образцы крови у всех животных отбирали в 2002 году.
Анализ полиморфизма и распределения аллельных вариантов белков выполняли электрофоретическим разделением в крахмальном и полиакриламидном геле (ПААГ) c гистохимическим окрашиванием (1, 6-8). Изучали следующие генетико-биохимические системы: из группы транспортных белков — гемоглобин (НB) и Х-белок (ХР), трансферрин (TF), отвечающий за транспорт железа, посттрансферин (PTF-2), рецептор витамина D (GC); из группы ферментов внутриклеточного энергетического метаболизма — регулятор активности лактатдегидрогеназы (LDR) и малик-энзим (ME); фермент пуринового обмена — пуриннуклеозидфосфорилазу (PN); из группы ферментов, катализирующих восстановление метгемогло-бина, — диафоразу (DP); из группы ферментов метаболизма экзогенных субстратов — эстеразу (EST) и фермент, гидролизующий карбоновые эфиры нафтола, — карбоангидразу (СA).
Сравнивали группы I-IV c породными группами 1978, 1980 и 1982 годов рождения и исходными породами — алтайской, ромни-марш и линкольн по следующим локусам: TF, HB, LDR, DP, EST и СA (1).
Для статистической обработки полученных данных использовали стандартную компьютерную программу BIOSIS-1.
Результаты. Все группы овец различались по особенностям распре-деления аллельных вариантов большинства исследованных локусов. По локусам НB, ME и GC межгрупповые различия отсутствовали (табл. 1) и индекс идентичности равнялся 1. По локусу ХP молодняк IV группы (частота аллеля Х+ — 0,793) был наиболее близок к овцематкам II группы (частота аллеля Х+ — 0,739) (показатель индекса идентичности — 1), особи I (Х+ — 0,652) и III (Х+ — 0,632) групп различались незначительно (индекс идентичности соответственно 0,986 и 0,979).
По локусу TF незначитель-ные различия наблюдались между животными II и III групп (индекс идентичности 0,937). Следует отме-тить у молодняка 2002 года рождения (IV группа) наличие редкого аллельного варианта TfF с частотой 0,017. По локусу PTF2 овцематки I и II групп (частота pTf-2а — соответственно 0,348 и 0,500) различались незначительно (индекс идентичности — 0,977).
По локусу LDR наблюдалось следующее распределение аллельных вариантов: у овцематок I группы частота аллеля Ldr-1а составляла 0,524, II — 0,646 (индекс идентичности между ними — 0,993), III и IV — соответственно 0,556 и 0,633, что обусловило их сходство с особями II группы.
Наибольшие различия между группами животных выявлены по локусу DP. Частота аллеля Dp-1а в I группе была равна 0,413, во II и IV — соответственно 0,520 и 0,583 (индекс идентичности между ними — соответственно 0,998 и 0,961). Особи III группы, у которых образцы крови отбирали примерно в 1,5-летнем возрасте, существенно (χ2 = 7,7; Р < 0,01) отличались от овцематок I группы, которые были на 3 года старше, по распределению аллельных вариантов в локусе DP(частота Dp-1а— 0,711).
По локусу PN незначительные различия по распределению аллельных вариантов отмечались между молодняком IV группы (частота NpH — 0,833) и животными I (NpH — 0,957) и III (NpH — 0,947) групп (значения индекса идентичности соответственно 0,993 и 0,995). По локусу EST наблюдали небольшие различия между особями II (частота Est-1а— 0,429) и III (Est-1а— 0,579) групп, с одной стороны, и молодняком IV (Est-1а — 0,552) — c другой (индекс идентичности соответственно 0,980 и 0,990).
Локус СА у животных I-IV групп оказался мономорфным и со-держал только ген «медленного» аллельного варианта белка — Са-S.
Средняя гетерозиготность по локусам в I, II, III и IV группах составляла соответственно 0,345; 0,362; 0,385 и 0,376. Наиболее низкие показатели гетерозиготности выявили по локусам HB и ME, наиболее высокие — по локусам EST, LDR, TF и PTF.
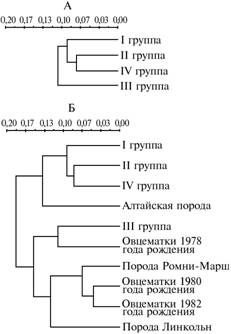
|
Генетические дистанции между представителями новой породной группы кроссбредных овец сибирской мясошерстной породы разных поколений и исходными породами: А — анализируемые помесные половозрастные группы животных текущих поколений; Б — эти же группы в сравнении с кроссбредными особями предыдущих поколений и исходными породами. Описание групп см. в разделе «Методика». |
На построенной дендрограмме особи II и IV групп образовали общий кластер (рис.). При сравнительном анализе полиморфизма по ряду молекулярно-генетических маркеров (5 локусов) (1) у овец II-IV групп и ранее исследованных исходных пород (алтайская, ромни-марш и линкольн), а также породных групп животных, рожденных в 1978, 1980 и 1982 годах, локус СА оказался мономорфным и фиксированным по аллельному варианту Ca-S во всех исследованных группах, кроме представителей алтайской породы, у которых выявлялся аллельный вариант Сa-F с частотой 0,010.
Частоты встречаемости аллельных вариантов по другим локусам для 10 исследованных групп представлены в таблице 2. По локусу TF у особей пород алтайская, ромни-марш и овцематок 1980 года рождения частота встречаемости аллеля Tf-A составляла соответственно 0,506; 0,496 и 0,517, у овец породы линкольн в этом локусе значительно чаще встречался аллель Tf-C (0,408). В то же время по указанному локусу не отмечали значительных различий между исходными породами и полученными кроссбредными овцами всех поколений, за исключением IV группы, в которой проявилась тенденция к увеличению частоты встречаемости аллельного варианта Tf-D (0,342). Статистически достоверные различия по распределению аллельного варианта Tf-A наблюдались между особями IV группы и рожденными в 1980 году (χ2 = 11,7; Р < 0,05). Также достоверные различия были выявлены между животными пород алтайская и линкольн, алтайская и овцематками 1978 года рождения (χ2 = 14,5 при Р < 0,01 и χ2 = 36,1 при Р < 0,001), ромни-марш и линкольн, ромни-марш и овцематками 1978 года рождения (χ2 = 11,8 при Р < 0,01 и χ2 = 28,3 при Р < 0,001), линкольн и овцематками 1980 года рождения (χ2 = 15,6 при Р < 0,01), животными 1978 и 1980, а также 1978 и 1982 годов рождения (соответствен но χ2 = 37,4 χ2= 19,4 при Р < 0,001). Интересно отметить у представителей исходных пород (кроме овец породы линкольн и поколений 1978-1982 годов рождения) присутствие редко встречающихся аллельных вариантов Tf-E и Tf-F, которые практически отсутствовали у овец последующих поколений (I-IV группы).
По локусу НВ у овец всех групп отмечали низкую частоту встречаемости аллельного варианта Нb-A, причем у животных породы линкольн он оказался мономорфным с наличием только аллельного варианта Hb-B (1). Особи I группы достоверно отличались от овцематок породы линкольн и рожденных в 1978 году (соответственно χ2 = 9,0 и χ2 = 9,2 при Р < 0,01). Необходимо подчеркнуть, что овцы породы линкольн достоверно различались с животными II (χ2 = 6,2 при Р < 0,05) и III (χ2 = 9,6 при Р < 0,01) групп, овцами пород алтайская (χ2 = 54,4 при Р < 0,001), ромни-марш (χ2 = 17,8 при Р < 0,001), овцематками 1978 (χ2 = 4,0 при Р < 0,05), а также 1980 и 1982 (соответственно χ2 = 16,5 χ2 = 21,3 при Р < 0,001) годов рождения. В свою очередь, овцы алтайской породы по этому локусу имели достоверные различия с животными породы ромни-марш (χ2 = 13,4 при Р < 0,001), рожденными в 1978 (χ2 = 44,2 при Р < 0,001), а также в 1980 и 1982 (соответственно χ2 = 8,3 и χ2 = 6,8 при Р < 0,01) годах.
По локусу LDR значительные отличия от остальных групп выявили у представителей породы линкольн и овец, рожденных в 1978 году (Ldr-1а— соответственно 0,415 и 0,425, индекс идентичности 1). В то же время особи II группы и молодняк IV (частота Ldr-1а— соответственно 0,646 и 0,633) достоверно отличались от животных породы ромни-марш (частота Ldr-1а— 0,471; соответственно χ2 = 4,9 и χ2 = 5,0 при Р < 0,05), породы линкольн (частота Ldr-1а— 0,415; χ2 = 7,2 и c2 = 7,4 при Р < 0,01) и новой породной группы 1978 года рождения (частота Ldr-1а— 0,425; c2 = 6,8 и c2 = 7,1 при Р < 0,01). Овцы алтайской породы (частота Ldr-1а— 0,564) по распределению аллельных вариантов по этому локусу были близки к особям I, III и IV групп (частота Ldr-1а— соответственно 0,524; 0,646 и 0,633, индекс идентичности — 1; 0,999 и 1) и достоверно отличались от животных породы линкольн и рожденных в 1978 году (соответственно χ2= 6,4 и χ2 = 6,0 при Р < 0,05).
По локусу DP наиболее близкими друг другу оказались овцематки IV группы и рожденные в 1978 году (частота Dp-1а— соответственно 0,711 и 0,755, индекс идентичности 1). При этом животные IV группы имели достоверные различия с овцами алтайской породы (χ2 = 12,7 при Р < 0,001) и особями I группы (χ2 = 7,7 при Р < 0,01). В тоже время по частоте Dp-1а животные I группы достоверно отличались от овец породы ромни-марш (0,613, χ2 = 6,1 при Р < 0,05), линкольн (0,644, χ2 = 7,0 при Р < 0,01), рожденных в 1978 (0,755, χ2 = 19,1 при P < 0,001) и 1982 (0,654, χ2 = 8,8 при P < 0,01) годах. Молодняк IV группы достоверно отличался от овец алтайской породы и овцематок 1978 года рождения (соответственно χ2 = 6,1 и χ2 = 6,3 при Р < 0,05). В свою очередь, овцы алтайской породы имели достоверные отличия от овец пород ромни-марш (χ2 = 18,3 при Р < 0,001), линкольн (χ2 = 16,2 при Р < 0,001), а также рожденных в 1978 (χ2 = 55,1 при Р < 0,001), 1980 (χ2 = 7,7 при Р < 0,01) и 1982 (χ2 = 24,9 при Р < 0,001) годах. У особей алтайской породы (частота Dp-1а— 0,402) индекс идентичности с животными I (Dp-1а— 0,413) и II (Dp-1а— 0,520) групп составлял соответственно 1 и 0,985.
По локусу EST следует отметить высокую частоту встречаемости аллеля Est-1а у овец практически всех ранее исследованных групп (за исключением животных алтайской породы) в отличие от представителей поколений 1997-2002 годов рождения (I-IV группы): у овец породы линкольн — 0,944, ромни-марш — 0,803, рожденных в 1980 году — 0,818, в 1982 — 0,825, тогда как у овец алтайской породы, овцематок I и II групп отмеченный показатель составлял соответственно 0,674; 0,429 и 0,477. При этом особи I и II групп достоверно отличались от овец алтайской породы (соответственно χ2 = 6,3 при Р < 0,05 и χ2 = 9,4 при Р < 0,01), ромни-марш (χ2 = 18,1 и χ2 = 22,7 при Р < 0,001), линкольн (χ2 = 42,0 и χ2 = 47,8 при Р < 0,001), рожденных в 1978 (соответственно χ2 = 9,4 при Р < 0,01 и χ2 = 12,9 при Р < 0,001), 1980 (соответственно χ2 = 20,6 и χ2 = 25,4 при Р < 0,001) и 1982 (χ2 = 22,1 и χ2 = 27,1 при Р < 0,001) годах. Овцематки II группы и молодняк IV, в свою очередь, также имели достоверные отличия от овец породы ромни-марш (соответственно χ2= 8,0 при Р < 0,01 и χ2 = 14,1 при Р < 0,001), линкольн (χ2= 25,3 и χ2= 38,5 при Р < 0,001), рожденных в 1980 (χ2 = 9,5 при Р < 0,01 и χ2= 16,6 при Р < 0,001) и 1982 (χ2 = 10,3 при P < 0,01 и χ2 = 18,1 при P < 0,001) годах. Животные алтайской породы достоверно отличались от представителей других исходных пород — ромни-марш (χ2 = 12,5 при Р < 0,001) и линкольн (χ2 = 49,2 при Р < 0,001).
На дендрограмме (см. рис.) овцематки I, II и IV групп, а также особи исходной алтайской породы образовали кластер. При этом животные II и IV групп выделились в подкластер, к которому поочередно присоединились овцематки I группы и исходной алтайской породы. В основу другого кластера вошли два подкластера: первый — овцематки III группы и рожденные в 1978 году, второй — овцематки 1980 и 1982 годов рождения и примкнувшие к ним представители исходных пород ромни-марш и линкольн.
Обращает на себя внимание тот факт, что поколения овец новой породной группы 1997, 1998 и 2002 годов рождения сближаются с родительской алтайской породой по распределению аллельных вариантов локусов LDR, DP и EST, а поколение 2000 года рождения — с исходными родительскими породами линкольн и ромни-марш по структуре локусов DPи TF.
Таким образом, полученные данные свидетельствуют о наличии определенной стабильности (консолидированности) групп овцематок, рожденных в 1997 и 1998 годах, а также близости к ним молодняка 2002 года рождения. Образование этими группами животных одного кластера с исходной алтайской полутонкорунной породой свидетельствует об их сближении с высокоадаптированной к местным условиям отечественной породой. В то же время уклонение генетической структуры овцематок 2000 года рождения в сторону пород линкольн и ромни-марш может отражать увеличение интенсивности искусственного отбора именно в этой группе животных. Полученные данные подтверждают, что в поколениях новой сибирской мясошерстной породы продолжается эволюция генетической структуры, связанная, по-видимому, с популяционно-генетическим ответом на изменяющееся соотношение между искусственным и естественным отбором. Обнаруживаются выраженные локус-специфические особенности отличий разных поколений этой породы как друг от друга, так и от исходных пород. Так, в поколениях 1997-2002 годов рождения выявлено сближение с генетической структурой родительской алтайской породы по распределению частот аллелей локуса эстеразы, 1978-1982 и 2000 годов ? наблюдается относительно бoльшая близость с улучшающими родительскими породами линкольн и ромни-марш по локусам трансферрина, гемоглобина и диафоразы.
Л И Т Е Р А Т У Р А
1. Г л а з к о В.И. Биохимическая генетика овец. Новосибирск, 1985.
2. М и н и н а Е.К. Фенотипическая и генотипическая изменчивость признаков продуктивности нового сибирского типа советской мясошерстной породы овец. Генетика, 2000, 36(7): 947-951.
3. В а с и н Б.Н. Генетика овец. В 4-х кн. М., 1928-1932.
4. Э р н с т Л.К., Д м и т р и е в Н.Г., П а р о н я н И.А. Генетические ресурсы сельскохозяйственных животных в России и сопредельных странах. СПб, 1994.
5. Д у б и н и н Н.П., Г л е м б о ц к и й Я.Л. Генетика популяций и селекция. М., 1967.
6. G a h n e B., J u n e j a R.K., G r o l m u s J. Horizontal polyacrylamide gradient gel electrophoresis for the simultaneous phenotyping of transferrin, post-transferrin, albumin and post-albumin in the blood plasma of cattle. Anim. Blood Groups Biochem. Genet., 1977, 8(3): 127-37.
7. L i v s h i t s G., K o b y l a n s k y E. Lerner’s concept of development homeostasis and the problem of heterozygosity level in natural populations. Heredity, 1985, 55: 341-353.
8. L e a r y R.F., A l l e n d o r f F.W., K n u d s e n K.L. Differences in inbreeding coefficients do not explain the association between heterozygosity at allozyme loci and developmental stability in rainbow trout. Evolution, 1987, 41(6): 1413-1415.
DYNAMICS OF GENETIC STRUCTURE OF MEAT-WOOLY SHEEPS OF THE SIBERIAN TYPE CREATED WITH USE OF REPRODUCTIVE POLYCARPIC BREEDING
S.V. Pavlenko, E.K. Minina, A.V. Kushnir, V.I. Glazko
The analysis of genetic structure was made in few generations of meat-wooly sheeps with the use of different molecular-genetic markers for estimation its consolidation. The locus-specific features of genetic correlations were revealed between the generations of new breed and initial breeds.
Key words: molecular-genetic markers, polymorphism, genetic distances, multibreed crossings, sheep breeds.
ФГОУ ВПО Российский государственный аграрный университет РГАУ?МСХА им. К.А. Тимирязева, |
Поступила в редакцию |
Оформление электронного оттиска
![]()
![]()




