doi: 10.15389/agrobiology.2022.3.555rus
УДК 633.9:631.461.52
Авторы благодарят Ю.В. Хомякова (ФГБНУ АФИ, г. Санкт-Петербург) за проведение агрохимического анализа почв и Е.Е. Андронова (ФГБНУ ВНИИСХМ, г. Санкт-Петербург) за помощь в проведении фотосъемки клубеньков. Работа выполнена при финансовой поддержке Российского научного фонда (проект № 21-16-00084).
ЭФФЕКТИВНОСТЬ АЗОТФИКСИРУЮЩЕГО СИМБИОЗА ГУАРА (Cyamopsis tetragonoloba) СО ШТАММАМИ Bradyrhizobium retamae RCAM05275 И Ensifer aridi RCAM05276 В ВЕГЕТАЦИОННОМ ОПЫТЕ
П.С. УЛЬЯНИЧ1, А.А. БЕЛИМОВ1 ✉, И.Г. КУЗНЕЦОВА1, А.Л. САЗАНОВА1, О.С. ЮЗИХИН1, Ю.В. ЛАКТИОНОВ1,
Д.С. КАРЛОВ1, М.А. ВИШНЯКОВА2, В.И. САФРОНОВА1
Бобовая культура гуар (Cyamopsis tetragonoloba (L.) Taub.) — источник гуаровой камеди (комплекс полисахаридов, который используется в различных отраслях промышленности). Эта культура широко возделывается в основном в Индии и Пакистане, но в последние годы возрастает интерес к промышленному выращиванию гуара в южных регионах России. Одна из проблем внедрения этой культуры в сельское хозяйство Российской Федерации — отсутствие в почвах бактерий, способных образовывать симбиотические клубеньки на корнях гуара в почвенно-климатических условиях России. В представляемой работе получены первые данные по эффективности инокуляции клубеньковыми бактериями гуара при выращивании в почвах России. Цель нашего исследования состояла в оценке эффективности симбиоза перспективных штаммов клубеньковых бактерий гуара Bradyrhizobium retamae RCAM05275 и Ensifer aridi RCAM05276 при выращивании растений в почвах, отобранных в разных районах Российской Федерации и не содержащих комплементарных клубеньковых бактерий. Для инокуляции семян гуара сорта Кубанский Юбилейный использовали инокулюмы в виде водных суспензий бактерий, полученные по стандартной методике приготовления жидких биопреператов клубеньковых бактерий. Эффективность симбиоза изучали в вегетационном опыте с выращиванием растений в дерново-подзолистой почве и чернозёме. Инокуляция обоими штаммами привела к активному образованию клубеньков (около 20-40 шт. на растение), в то время как на корнях контрольных растений клубеньков обнаружено не было. Количество клубеньков на растении было максимальным в вариантах инокуляции штаммом B. retamae RCAM05275. Значения общей массы клубеньков на одно растение были максимальными в вариантах инокуляции штаммом E. aridi RCAM05276 благодаря образованию более крупных клубеньков. Клубеньки формировались на боковых корнях, имели округлую неправильную форму, розоватый цвет (свидетельство наличия в них леггемоглобина) и существенно варьировали по размеру. Оба штамма повысили биомассу побегов и всего растения примерно на 70 % при выращивании на дерново-подзолистой почве и черноземе, но не повлияли на биомассу корней. Инокулированные растения обладали примерно одинаковой нитрогеназной активностью независимо от штамма ризобий и типа почвы. Удельная нитрогеназная активность (на единицу биомассы клубенька) была примерно в 2 раза выше по сравнению с другими вариантами при инокуляции растений штаммом B. retamae RCAM05275 в дерново-подзолистой почве. Во всех вариантах опыта в побегах инокулированных растений выявлено повышение содержания общего азота примерно в 1,4 раза и накопления азота — в 3-4 раза. Таким образом, оба изученных штамма оказались способны формировать азотфиксирующий симбиоз, что привело к значительному росту биомассы растений и накоплению азота в побегах. Результаты показали перспективность дальнейших исследований по испытанию штаммов в полевых экспериментах с целью создания биопрепаратов для улучшения азотного питания этой сельскохозяйственной культуры.
Ключевые слова: азотфиксация, гуар, клубеньки, симбиоз, Cyamopsis tetragonoloba.
Гуар Cyamopsis tetragonoloba (L.) Taub. — одна из важнейших тропических бобовых культур, в семенах которой содержится гуаровая камедь, используемой в угольной, нефтегазовой, пищевой, текстильной, бумажной и косметической промышленности. Гуар широко культивируется в Индии и Пакистане, а также в Афганистане, Кении, Австралии и полупустынных регионах США (1). Спрос на гуаровую камедь постоянно растет, и в 2016 году импорт гуаровой камеди в Россию превысил 15 тыс. т (2). В Россию гуар был завезен в середине 1920-х годов (3), но широкого распространения не нашел из-за недостаточных знаний о технологии его возделывания (4). В последние годы интерес к промышленному выращиванию гуара в южных регионах России возрастает (5). Одним из аспектов решения этой задачи — поиск штаммов клубеньковых бактерий (ризобий), способных образовывать эффективный азотфиксирующий симбиоз с гуаром в почвенно-климатических условиях Российской Федерации и создание биопрепаратов на основе таких микроорганизмов.
Инокуляция семян биопрепаратами клубеньковых бактерий обеспечивает интенсивную биологическую азотфиксацию, которая способствует усилению фотосинтеза и увеличению урожайности бобовых культур (6, 7). Использование ризобий для инокуляции особенно важно при возделывании бобовых на новых территориях, почвы которых не содержат необходимых микросимбионтов. Например, при попытке выращивания сои культурной Glycine max (L.) Merr. в нетипичных для вида географических зонах России клубеньки на корнях практически не формировались, поэтому потребовалась разработка биопрепаратов и инокуляция семян специфичными ризобиями (6). В настоящее время для решения этой проблемы производители семян сои обеспечивают их поставку совместно с биопрепаратами клубеньковых бактерий (https://kingsagriseeds.com/soybeans/).
Медленнорастущие клубеньковые бактерии рода Bradyrhizobium (семейство Bradyrhizobiaceae) являются основной группой ризобий, вступающих в симбиоз с гуаром (8, 9). В большинстве случаев штаммы не были идентифицированы до вида, но многие были близки к виду B. japonicum (10, 11). К этому виду относится очень большая и генетически разнородная группа микросимбионтов сои, а также штаммы, нодулирующие разные виды вигны (Vigna), люпина (Lupinus), сераделлы (Ornithopus) и широкий спектр бобовыхрастений генистоидного комплекса (12-14).
Исследования показали, что инокуляция гуара штаммами Bradyrhizobium spр. оказывает положительное влияние на развитие растений, значительно увеличивает количество клубеньков (до 79 %), массу растений (до 71 %), массу корней (до 262 %), урожай семян (до 53 %), содержание белка (до 33 %), клетчатки (до 26 %), повышает общее содержание азота и минеральных веществ (8, 12, 15-17). Недавно описана способность гуара образовывать симбиоз с быстрорастущими ризобиями Ensiferaridi(семейство Rhizobiaceae), также обладающими широким спектром бобовых растений-хозяев из подсемейств Mimosoideaeи Papilionoideae (18). Наличие среди клубеньковых бактерий гуара представителей родов Bradyrhizobium и Ensifer— общая черта, объединяющая это растение с соей и вигной (19-21).
Для успешной интродукции гуара в России требуется (наравне с созданием новых сортов, разработкой технологий их возделывания и применения минеральных удобрений) селекция эффективных микросимбионтов этой культуры с целью создания биопрепаратов. В Индии применение минеральных азотных удобрений ограничено высокой стоимостью и низким уровнем механизации сельского хозяйства. Поэтому высокая продуктивность гуара достигается во многом благодаря наличию в почвах высокоэффективных штаммов клубеньковых бактерий. Следует подчеркнуть, что наблюдения российских селекционеров за корневой системой гуара в районах России, где его недавно начали возделывать (Краснодарский край, Крым, Астраханская и Волгоградская области и Дагестан), показали отсутствие симбиотических клубеньков.
Ранее мы выделили клубеньковые бактерии гуара, изучили их биоразнообразие и в результате охарактеризовали медленнорастущие штаммы рода Bradyrhizobium (22) и быстрорастущие штаммы рода Ensifer(неопубликованные данные).
Цель представляемой работы состояла в оценке способности образовывать эффективный симбиоз у двух перспективных штаммов ризобий родов Bradyrhizobium и Ensifer с гуаром в условиях вегетационного опыта при выращивании растений в почвах, отобранных в разных районах России и не содержащих комплементарных клубеньковых бактерий.
Методика. Штаммы клубеньковых бактерий Bradyrhizobium retamae RCAM05275 (23) и Ensifer aridi RCAM05276 (24), которые были выделены из клубеньков гуара, охарактеризованы в предварительных экспериментах и запатентованы авторами, депонированы в Ведомственной коллекции полезных микроорганизмов сельскохозяйственного назначения (ФГБНУ ВНИИСХМ, г. Санкт-Петербург). Для обработки семян бактериями использовали инокулюм, приготовленный в лабораторных условиях в виде образца жидкого биопреперата. Посевной материал чистых культур бактерий помещали в колбы с 250 мл полусинтетической питательной среды (г/л): маннит — 10; дрожжевой экстракт — 1; K2HPO4 — 0,5; MgSO4·7H2O — 0,2; NaCl — 0,1. Колбы инкубировали при 28 °С на качалке при 180 об/мин (в течение 3 сут — для штамма E. aridi RCAM05276, в течение 5 сут — для штамма B. retamae RCAM05275). По стандартной методике с соблюдением мер асептики содержимое колбы пересевали в лабораторный ферментер BIORUS 5L («BIORUS», Россия), содержащий 5 л такой же среды. Культивирование в ферментере проходило при 28 °С, интенсивности аэрации 1 л воздуха·1 л среды-1·мин-1 и перемешивании при 200 об·мин-1 в течение 3 сут (штамм E. aridi RCAM05276) или 4 сут (штамм B. retamae RCAM05275). Затем бактериальные суспензии (инокулюм) помещали в стерильную тару и хранили при комнатной температуре (22-24 °С) для последующего использования.
Семена гуара сорта Кубанский Юбилейный (получены во Всероссийском институте генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург) скарифицировали и поверхностно стерилизовали в 98 % H2SO4 в течение 30 мин, промывали стерильной водопроводной водой и проращивали на фильтровальной бумаге в чашках Петри при температуре 25 °С в темноте в течение 2 сут. Для вегетационного опыта использовали дерново-подзолистую почву (Albic Retisol, Abruptic, Ochric, Псковская обл.) и чернозем (Haplic Chernozem, Pachic, Воронежская обл.), отобранную летом 2017 года. Дерново-подзолистая почва была предоставлена Псковским НИИ сельского хозяйства и совхозом «Родина» (57°50′44.2′′ N, 28°12′03.7′′ E), чернозем получен в заповеднике «Каменная степь» (51°01′41.6′′ N, 40°43′39.3′′ E). Агрохимические показатели сухой почвы измеряли по стандартным методикам (25). Почву вносили в металлические сосуды по 400 г и удобряли K2HPO4 (600 мг/кг). Проросшие семена выдерживали 1 ч в инокулюме и высаживали в сосуды (2 сосуда по 4 проростка на каждый вариант опыта). Растения культивировали в течение 106 сут в фитокомнате с относительной влажностью 60 % при 2-уровневом режиме освещенности и температуры: ночь (темнота, 18 °С, 8 ч), день (400 мклкŁм-2·с-1, 23 °С, 16 ч). Для освещения использовались лампы ДНаТ (ООО «ССЗ Лисма», Россия) и L36W/77 FLUORA («OSRAM Licht AG», Германия). Влажность почвы поддерживали на уровне 60-70 % полной влагоемкости регулярным поливом при взвешива-нии сосудов. На 45-е сут культивирования почву дополнительно удобряли раствором K2HPO4 (600 мг/кг).
По окончании эксперимента корни растений вместе с образовавшимися клубеньками отделяли от побегов, промывали водопроводной водой, помещали в герметичные пластиковые пробирки и добавляли ацетилен в количестве 5 % от объема пробирки для измерения биологической азотфиксации ацетиленовым методом. Для этого после инкубации пробирок при 25 °С в темноте в течение 1 ч определяли количество образовавшегося этилена на газовом хроматографе GC-2014 c ПИД-детектором (FID) и колонкой SUS 2.0 m½3.0 mm (L½ID) («Shimadzu Corporation», Япония). Анализ проводили при следующих параметрах: температура детектора — +250 °С; газ-носитель — азот; скорость потока газа-носителя — 70 мл/мин; температура инжектора — +72 °С; температура колонки — +70 °С; время анализа — 5,0 мин; время удерживания этилена — 1,20±0,01 мин; время удерживания ацетилена — 2,00±0,01 мин.
Фотографии корней делали с использованием фотокамеры PC1742 («Cannon», Япония), клубеньков — с помощью стереомикроскопа Stemi 508 («Carl Zeiss», Германия).
Клубеньки отделяли от корней и определяли их число на каждом растении. Побеги, корни и клубеньки высушивали при комнатной температуре и взвешивали. Содержание общего азота в побегах определяли на автоматическом анализаторе Kjeltec 8200 («FOSS Analytical», Дания) по стандартной методике производителя.
Статистический анализ данных проводили с использованием программного обеспечения STATISTICA v. 10 («TIBCO Software Inc.», США). Для оценки различий между средними значениями использовали однофакторный дисперсионный анализ и критерий НСР Фишера, равенство дисперсий выборок проверяли с помощью критерия Левене (Levene’s Test).
Результаты. Агрохимические показатели образцов сухой почвы представлены в таблице.
Инокуляция гуара обоими штаммами привела к активному образованию клубеньков (около 20-40 шт. на растение, рис. 1, А), в то время как на корнях контрольных растений клубеньков мы не обнаружили. При выращивании растений на черноземе число клубеньков, образованных штаммом B. retamae RCAM05275, было в 2,8 раза больше, чем у растений, выращенных на дерново-подзолистой почве и инокулированных штаммом E. aridi RCAM05276. Общая масса клубеньков на одном растении была максимальной при инокуляции E. aridi RCAM05276 на дерново-подзолистой почве и черноземе, однако различия между вариантами опыта не были достоверными из-за значительной вариации этого параметра (см. рис. 1, Б).
Ранее было показано, что образование клубеньков у гуара могут ингибировать высокие концентрации доступного почвенного азота (26). В использованном нами черноземе содержание разных форм азота было в 1,5-2 раза выше, чем в дерново-подзолистой почве (см. табл.). Но это не привело к ингибированию клубенькообразования, что свидетельствует о способности обоих партнеров образовывать симбиоз при различном содержании почвенного азота.
По сравнению со штаммомB. retamae RCAM05275 штамм E. aridi RCAM05276 образовывал более крупные клубеньки на растениях (см. рис. 1, В). Максимально крупные клубеньки у штамма E. aridi RCAM05276 формировались на дерново-подзолистой почве. Информация о влиянии типа и состава почвы на размер симбиотических клубеньков и роль штамма ризобий в этой зависимости очень ограничена.
Известно, что растение контролирует образование (число и биомассу) и функционирование (приток фотосинтатов и транспорт азотных соединений) клубеньков (27). Число и биомасса образуемых клубеньков также существенно варьируют в зависимости от штамма микросимбионта (6, 28, 29). Процессы формирования симбиоза зависят от физико-химических свойств почвы, таких как содержание органического вещества, доступного растениям азота, кислотности и других факторов (29, 30). Большое влияние на число образуемых клубеньков оказывают аборигенные ризосферные микроорганизмы, которые модулируют гормональный статус и обеспеченность растения питательными веществами и взаимодействуют с интродуцентами (31). Наблюдаемый нами феномен влияния почвы на биомассу клубенька может быть связан с несколькими перечисленными факторами и его объяснение требует более детального изучения.
Оба штамма повысили биомассу побегов примерно на 70 % при выращивании гуара на дерново-подзолистой почве и черноземе (рис. 2, А), но не повлияли на биомассу корней (см. рис. 2, Б). В результате биомасса всего инокулированного растения во всех вариантах опыта также была больше, чем в контроле, примерно на 60-80 % (см. рис. 2, В). Это согласуется с данными литературы о высокой отзывчивости гуара на инокуляцию клубеньковыми бактериями, что проявлялось в увеличении биомассы надземной части растений и урожая семян (8, 11, 13).
На рисунке 3 в качестве примера представлен внешний вид надземной части, корневая система и клубеньки растений, выращенных на дерново-подзолистой почве, в день окончания эксперимента. Контрольные растения существенно уступали инокулированным в высоте и площади листьев, проявляли признаки хлороза, вероятно обусловленного дефицитом азота (рис. 3, А). Корни были хорошо развиты, разветвлены, но не имели клубеньков (см. рис. 3, Б). Оба штамма формировали клубеньки на боковых корнях, при этом встречались единичные клубеньки и группы тесно расположенных клубеньков (см. рис. 3, В, Д). Клубеньки обоих штаммов имели неправильную округлую форму и существенно варьировали по размеру (см. рис. 3, Г, Е). Розоватый цвет клубеньков свидетельствовал о наличии в них леггемоглобина, который необходим для фиксации атмосферного азота.
Инокулированные растения обладали примерно одинаковой нитрогеназной активностью независимо от штамма ризобий и типа почвы (рис. 4, А). Но удельная нитрогеназная активность, выраженная на единицу биомассы клубенька, была в 1,5-2,4 раза выше в дерново-подзолистой почве при инокуляции растений штаммом B. etamae RCAM05275, чем в других вариантах (см. рис. 4, Б). Вероятно, что относительно невысокая биомасса клубеньков (см. рис. 1, Б) компенсировалась повышением эффективности функционирования азотфиксирующей системы с участием штамма B. retamae RCAM05275. Сравнение штаммов и измерение активности азотфиксации в этих исследованиях не проводилось, однако ранее было показано, что масса клубеньков коррелировала с накоплением азота и урожаем при сравнении 50 генотипов гуара (32). Об уровне нитрогеназной активности клубеньков гуара известно мало, но полученные нами значения были сравнимы с данными по измерению нитрогеназной активности у сои при инокуляции различными штаммами родов Bradyrhizobium и Ensifer (33, 34).
Об эффективном функционировании симбиоза свидетельствовало также повышение содержания общего азота примерно в 1,4 раза (см. рис. 4, В) и накопления азота — примерно в 3-4 раза (см. рис. 4, Г) в побегах инокулированных растений гуара. Эти эффекты проявились во всех вариантах опыта с минимальным значением при инокуляции выращенных на дерново-подзолистой почве растений штаммом E. aridi RCAM05276. Существенное повышение содержания и накопления азота в растениях гуара в результате инокуляции клубеньковыми бактериями ранее было неоднократно описано (8, 11, 12, 35). Было также показано, что в симбиозе с соей штаммы рода Bradyrhizobium более активно фиксировали азот, чем штаммы рода Ensifer (34). В наших экспериментах B. retamae RCAM05275 не уступал штамму E. aridi RCAM05276 по измеряемым параметрам симбиоза, а на дерново-подзолистой почве повысил содержание и накопление азота в растениях на 26 % по сравнению с увеличением этих показателей под влиянием штамма E. aridi RCAM05276 (см. рис. 4, В, Г). В среднем для всех вариантов опыта биомасса отдельного клубенька отрицательно коррелировала с содержанием азота в побегах (r = -0,98; p = 0,019; n = 4), что свидетельствует о более высокой эффективности мелких клубеньков, которые образовывал штамм B. retamae RCAM05275.
Таким образом, оба штамма — B. retamae RCAM05275 и E. aridi RCAM05276 способны формировать эффективный симбиоз с гуаром при выращивании растений в дерново-подзолистой почве и черноземе, отобранных в разных районах Российской Федерации и не содержащих комплементарных клубеньковых бактерий. В основном штаммы имели сходство по параметрам образования клубеньков и эффективности симбиоза. Однако также выявлены характерные особенности штаммов во взаимодействии с растениями гуара, что выражалось в различиях по числу клубеньков, удельной нитрогеназной активности и интенсивности обеспечения растений азотом. Полученные нами результаты показали перспективность дальнейших испытаний изучаемых штаммов в полевых экспериментах с целью создания биопрепаратов для улучшения азотного питания растений гуара.
Агрохимические показатели почв, использованных в эксперименте |
||
Показатель |
Дерново-подзолистая |
Чернозем |
Массовая доля органического вещества, % |
2,4 |
8,8 |
Азот общий, % |
0,22 |
0,38 |
Азот аммиачный, мг N/кг |
25 |
37 |
Азот нитратный, мг N/кг |
9,6 |
26,3 |
Подвижный фосфор, мг P2O5/кг |
85 |
121 |
Подвижный калий, мг K2O/кг |
60 |
155 |
Гидролитическая кислотность, ммоль/кг |
29 |
18 |
Сумма поглощенных оснований (Са2+, Mg2+, Na+, К+, NH4+), ммоль/кг |
60 |
372 |
рНН2О |
6,1 |
7,3 |
рНKCl |
4,9 |
6,2 |
Примечание. Дерново-подзолистая почва и чернозем отобраны в 2017 году соответственно в Псковской (57°50′44.2′′ N, 28°12′03.7′′ E) и Воронежской (51°01′41.6′′ N, 40°43′39.3′′ E) областях. Массовую долю органического вещества определяли по Тюрину; общий азот — по Кьельдалю; аммиачный азот — с реактивом Несслера (экстракция 2 % раствором KCl); нитратный азот — дисульфофеноловым методом (экстракция 0,05 % раствором K2SO4); подвижный фосфор — по образованию фосфомолибденовой сини, подвижный калий — пламенно-фотометрическим методом, экстракцию из дерново-подзолистой почвы проводили 0,2 моль/л HCl, из чернозема — 10 г/л (NH4)2CO3; гидролитическую кислотность и сумму поглощенных оснований — методом Каппена, экстрагируя соответственно 1 н. раствором CH3COONa·3H2O и 0,1 н. раствором HCl. |
||
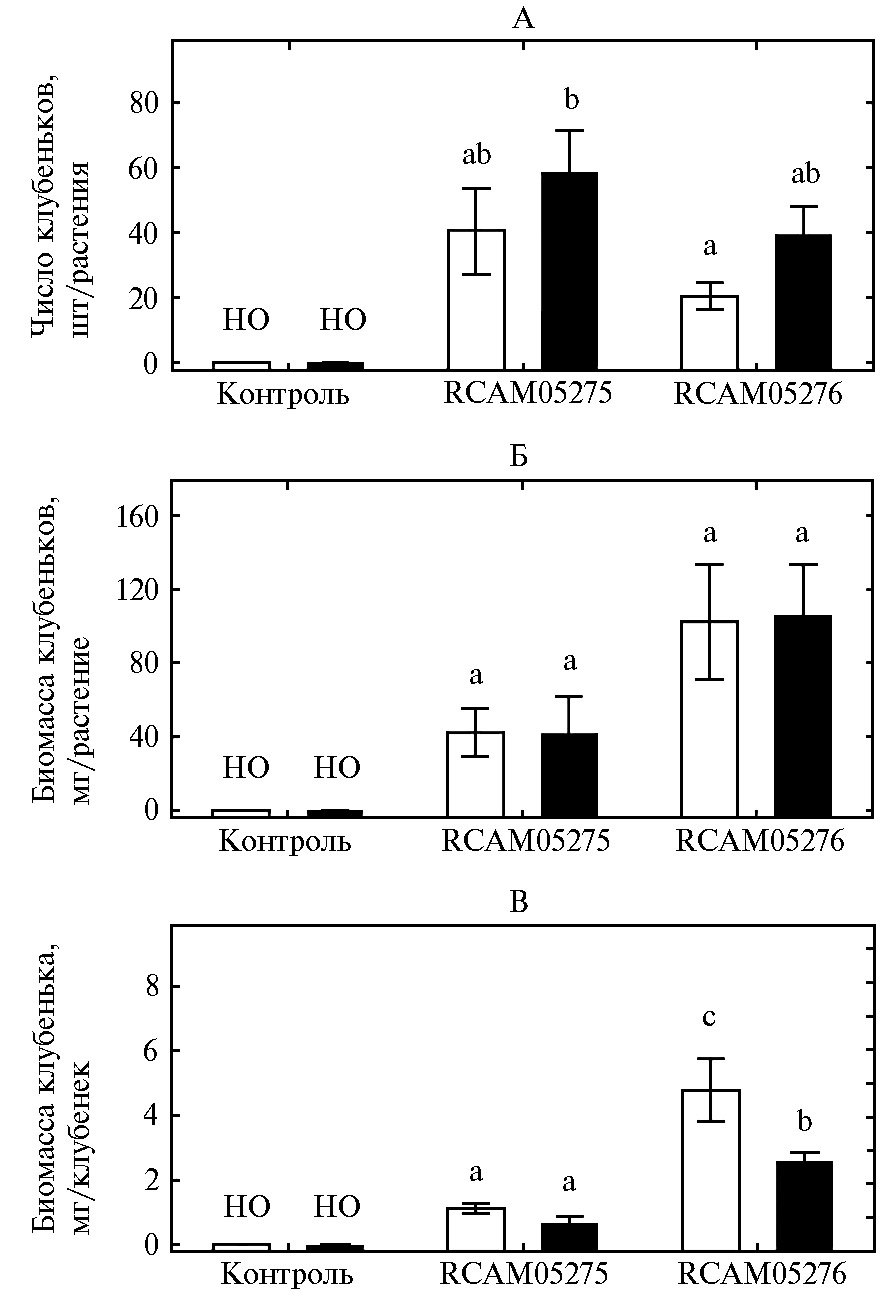
Рис. 1. Число клубеньков (А) и сухая биомасса клубеньков (Б) на одном растении и биомасса одного клубенька (В) на корнях гуара сорта Кубанский Юбилейный: контроль — без инокуляции, RCAM05275 — инокуляция штаммом Bradyrhizobium retamae RCAM05275, RCAM05276 — инокуляция штаммом Ensifer aridi RCAM05276; белые столбики — дерново-подзолистая почва, черные — чернозем. Вертикальные отрезки означают ошибки средних значений, но — не обнаружено. Разными латинскими буквами отмечены варианты, различия между которыми статистически значимы (n = 8, критерий НСР Фишера, p < 0,05; лабораторный опыт).
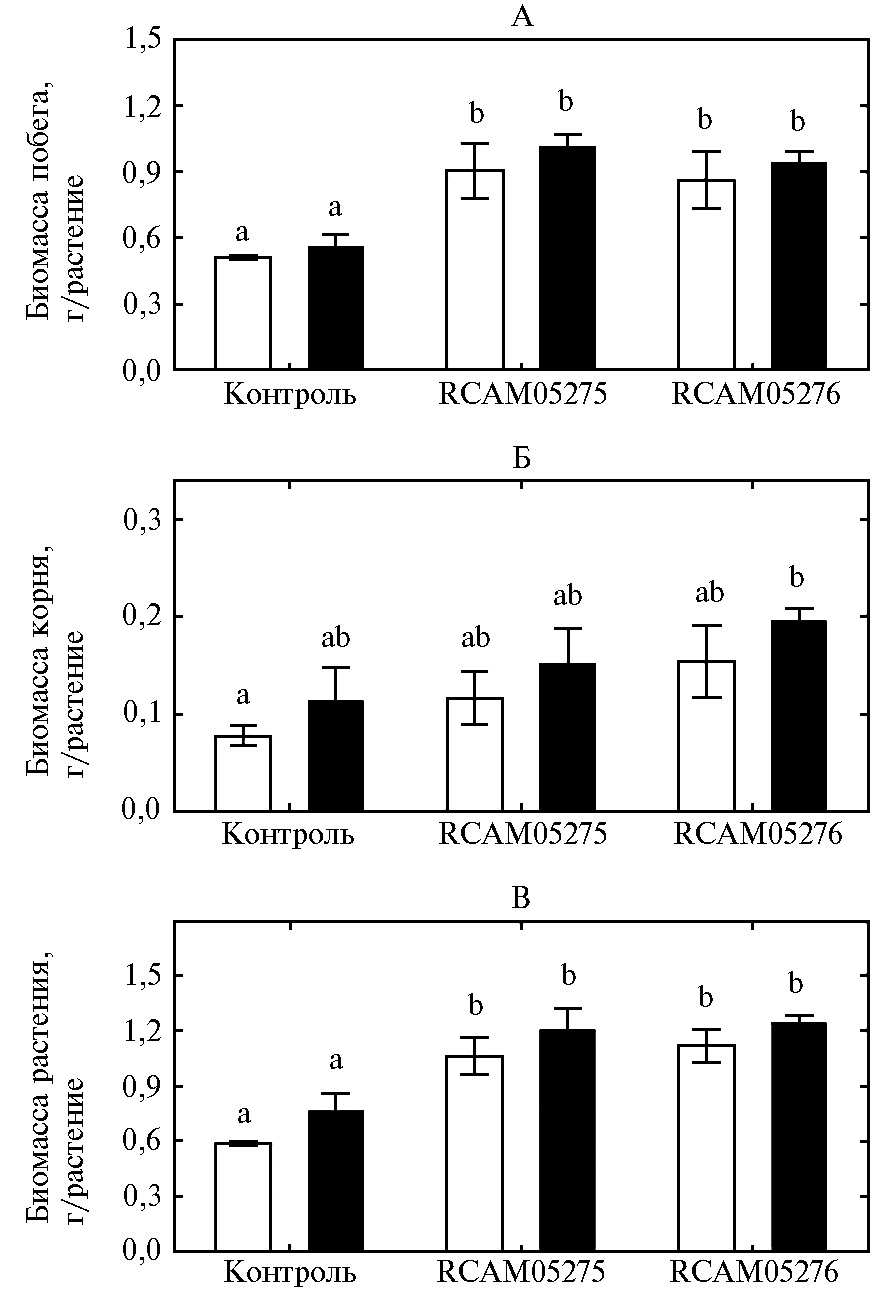
Рис. 2. Сухая биомасса побегов (А), корней (Б) и общая биомасса растения (В) гуара сорта Кубанский Юбилейный: контроль — без инокуляции, RCAM05275 — инокуляция штаммом Bradyrhizobium retamae RCAM05275, RCAM05276 — инокуляция штаммом Ensifer aridi RCAM05276; белые столбики — дерново-подзолистая почва, черные — чернозем. Вертикальные отрезки означают ошибки средних значений. Разными латинскими буквами отмечены варианты, различия между которыми статистически значимы (n = 8, критерий НСР Фишера, p < 0,05; лабораторный опыт).

Рис. 3. Внешний вид побегов (А), корневой системы (Б, В, Д) и клубеньков (Г, Е) растений гуара сорта Кубанский Юбилейный, выращенных на дерново-подзолистой почве: Б — контроль без инокуляции, В, Г — инокуляция штаммом Bradyrhizobium retamae RCAM05275, Д, Е — инокуляция штаммом Ensifer aridi RCAM05276 (лабораторный опыт).
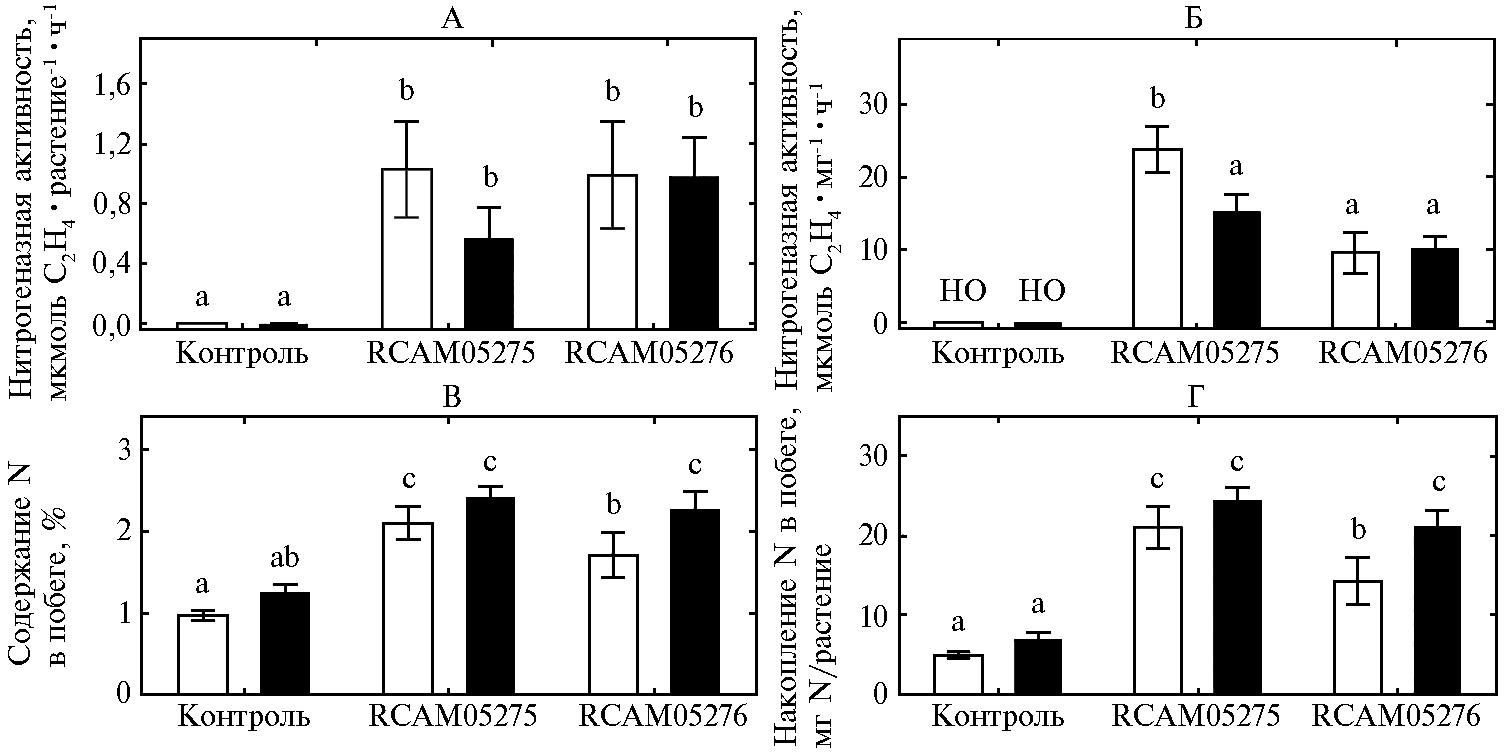
Рис. 4. Нитрогеназная (ацетилен-редуктазная) активность растения (А), удельная нитрогеназная активность на единицу биомассы клубенька (Б), содержание (В) и накопление (Г) общего азота в побегах гуара сорта Кубанский Юбилейный: контроль — без инокуляции, RCAM05275 — инокуляция штаммом Bradyrhizobium retamae RCAM05275, RCAM05276 — инокуляция штаммом Ensifer aridi RCAM05276; белые столбики — дерново-подзолистая почва, черные — чернозем. Вертикальные отрезки означают ошибки средних значений. Разными латинскими буквами отмечены варианты, различия между которыми статистически значимы (n = 8, критерий НСР Фишера, p < 0,05; лабораторный опыт).
ЛИТЕРАТУРА
- Лебедь Д.В., Костенкова Е.В., Волошин М.И. Агрономическое обоснование размещения посевов Cyamopsis tetragonoloba L. на юге европейской части России. Таврический вестник аграрной науки, 2017, 1(9): 53-63.
- Старцев В.И., Ливанская Г.А, Куликов М.А. Перспективы возделывания гуара Cyamopsis tetragonoloba L. в России. Вестник Российского государственного аграрного заочного университета, 2017, 24(29): 11-15.
- Вавилов Н.И. Интродукция растений в советское время и ее результаты. Избранные труды. М.-Л., 1965, т. V: 674-689.
- Волошин М.И., Лебедь Д.В., Брусенцов А.С. Результаты интродукции нового бобового растения — гуара (Cyamopsis tetragonoloba (L.) Taub.). Труды Кубанского государственного аграрного университета, 2016,1(58): 84-91.
- Булынцев С.В., Вальяникова Т.И., Силаева О.И., Копоть Е.И., Пимонов К.И. Гуар — новая бобовая культура для России. Мат. Всероссийской науч.-практ. конф. «Инновации в технологиях возделывания сельскохозяйственных культур». Пос. Персиановский, 2017: 167-172.
- Берестецкий О.А., Доросинский Л.М., Кожемяков А.П. Эффективность препаратов клубеньковых бактерий в географической сети опытов. Известия АН СССР, Серия биологическая, 1987, 5: 670-679.
- Кожемяков А.П., Лактионов Ю.В., Попова Т.А., Орлова А.Г., Кокорина А.Л., Вайшля О.Б., Агафонов Е.В., Гужвин С.А., Чураков А.А., Яковлева М.Т. Агротехнологические основы создания усовершенствованных форм микробных биопрепаратов для земледелия. Сельскохозяйственная биология, 2015, 50(3): 369-376 (doi: 10.15389/agrobiology.2015.3.369rus).
- Elsheikh El.A.E., Ibrahim K.A. The effect of Bradyrhizobium inoculation on yield and seed quality of guar (Cyamopsis tetragonoloba L.). Food Chemistry, 1999, 65(2): 183-187 (doi: 10.1016/S0308-8146(98)00192-7).
- Ibrahim K., Suliman K.H., Abdalla A.A., Mohamed E.A., Ahmed A.I., Mukhtar S. Response of growth, yield and seed quality of guar (Cyamopsis teteragonolopa L.) to Bradyrhizobium inoculations. Pakistan Journal of Nutrition, 2011, 10(9): 805-813 (doi: 10.3923/pjn.2011.805.813).
- Hassen A.I., Bopape F.L., Trytsman M. Nodulation study and characterization of rhizobial microsymbionts of forage and pasture legumes in South Africa. World Journal of Agricultural Research, 2014, 2(3): 93-100 (doi: 10.12691/wjar-2-3-2).
- Weaver R.W., Arayangkoon T., Schomber H.H. Nodulation and N2 fixation of guar at high root temperature. Plant and Soil, 1990, 126(2): 209-213 (doi: 10.1007/BF00012824).
- Stępkowski T., Zak M., Moulin L., Króliczak J., Golińska, B., Narożna D., Safronova V.I., Mądrzak C.J. Bradyrhizobium canariense and Bradyrhizobium japonicum are the two dominant rhizobium species in root nodules of lupin and serradella plants growing in Europe. Systematic and American Deserts. BMC Genomics, 2017, 18(1): 1-24 (doi: 10.1186/s12864-016-3447-y).
- Vinuesa P., León-Barrios M., Silva C., Willems A., Jarabo-Lorenzo A., Pérez-Galdona R., Werner D., Martínez-Romero E. Bradyrhizobium canariense sp. nov., an acid-tolerant endosymbiont that nodulates endemic genistoid legumes (Papilionoideae: Genisteae) from the Canary Islands, along with Bradyrhizobium japonicum bv. genistearum, Bradyrhizobium genospecies alpha and Bradyrhizobium genospecies beta. International Journal of Systematic and Evolutionary Microbiology, 2005, 55(2): 569-575 (doi: 10.1099/ijs.0.63292-0).
- Zhang Y.F., Chang E.T., Tian C.F., Wang F.Q., Han L.L., Chen W.F., Chen W.X. Bradyrhizobium elkanii, Bradyrhizobium yuanmingense and Bradyrhizobium japonicum are the main rhizobia associated with Vigna unguiculata and Vigna radiate in the subtropical region of China. FEMS Microbiology Letters, 2008, 285(2): 146-154 (doi: 10.1111/j.1574-6968.2008.01169.x).
- Elnesairy N.N., Abubaker J.A., Mahmod H., Mukhtar N. The impact of Bradyrhizobium, farmyard manure and inorganic nitrogen on growth and yield of guar. World Journal of Agricultural Research,2016, 4(2): 56-63 (doi: 10.12691/wjar-4-2-4).
- Ibrahim K.A., Suliman K.H., Abdalla A.A. Influence of inoculation with some Bradyrhizobium strains on yield attributes, seed proximate composition and minerals content of guar (Cyamopsis teteragonolopa L.) grown in Sudan. Australian Journal of Basic and Applied Sciences, 2010, 4(5): 808-816.
- Ibrahim K.A., Naeim E.A.M., Naim A.M.E., Elsheikh M.A. Response of Guar (Cyamopsis teteragonolopa L.) to Bradyrhizobium inoculations in semi-arid environment. International Journal of Agriculture and Forestry, 2016, 6(4):137-141 (doi: 10.5923/j.ijaf.20160604.01).
- Le Quéré A., Tak N., Gehlot H.S., Lavire C., Meyer T., Chapulliot D., Rathi S., Sakrouhi I., Rocha G., Rohmer M., Severac D., Filali-Maltouf A., Munive J. A. Genomic characterization of Ensifer aridi, a proposed new species of nitrogen-fixing rhizobium recovered from Asian, African and American deserts. BMC Genomics, 2017, 18(1): 1-24 (doi: 10.1186/s12864-016-3447-y).
- Chen W.X., Yan G.H., Li J.L. Numerical taxonomic study of fast-growing soybean rhizobia and a proposal that Rhizobium fredii be assigned to Sinorhizobium gen. nov. International Journal of Systematic and Evolutionary Microbiology, 1988, 38(4): 392-397 (doi: 10.1099/00207713-38-4-392).
- Ondieki D.K., Nyaboga E.N., Wagacha J.M., Mwaura F.B. Morphological and genetic diversity of rhizobia nodulating cowpea (Vigna unguiculata L.) from agricultural soils of Lower Eastern Kenya. International Journal of Microbiology, 2017, 2017: 8684921 (doi: 10.1155/2017/8684921).
- Yan H., Yan J., Sui X.H., Wang E.T., Chen W.X., Zhang X.X., Chen W.F. Ensifer glycinis sp. nov., a rhizobial species associated with species of the genus Glycine. International Journal of Systematic and Evolutionary microbiology, 2016, 66(8): 2910-2916 (doi: 10.1099/ijsem.0.001120).
- Кузнецова И.Г., Сазанова А.Л., Сафронова В.И., Попова Ж.П., Соколова Д.В., Тихомирова Н.Ю., Оследкин Ю.С., Карлов Д.С., Белимов А.А. Выделение и идентификация клубеньковых бактерий гуара Cyamopsis tetragonoloba (L.) Taub. Сельскохозяйственнаябиология, 2018, 53(6): 1285-1293 (doi: 10.15389/agrobiology.2018.6.1285rus).
- Белимов А.А., Кузнецова И.Г., Потокина Е.К., Сазанова А.Л., Сафронова В.И. Штамм клубеньковых бактерий гуара Bradyrhizobium retamae — стимулятор азотфиксирующей способности гуара. МПК: C12N 1/20, A01N 63/20, C05F 11/08, C12R 1/01. Федеральное бюджетное научное учреждение «Всероссийский научно-исследовательский институт сельскохозяйственной микробиологии» (RU). № 2734836. Заявка: 2020100530 от 09.01.2020. Опубл. 23.10.2020. Бюл. № 30.
- Белимов А.А., Кузнецова И.Г., Потокина Е.К., Сазанова А.Л., Сафронова В.И. Штамм клубеньковых бактерий гуара Ensifer aridi RCAM05276 — азотфиксирующий симбионт гуара. МПК: C12N 1/20, A01N 63/20, C05F 11/08, C12R 1/01. Федеральное бюджетное научное учреждение «Всероссийский научно-исследовательский институт сельскохозяйственной микробиологии» (РФ). № 2734836. Заявка: 2020100529 от 09.01.2020. Опубл. 20.11.2020. Бюл. № 32.
- Аринушкина Е.В. Руководство по химическому анализу почв. М., 1970.
- Hinson P.O., Adams C.B. Quantifying tradeoffs in nodulation and plant productivity with nitrogen in guar. Industrial Crops and Products, 2020, 153: 112617 (doi: 10.1016/j.indcrop.2020.112617).
- Ferguson B.J., Mens C., Hastwell A.H., Zhang M., Su H., Jones C.H., Chu X., Gresshoff P.M. Legume nodulation: The host controls the party. Plant, Cell & Environment, 2019, 42(1): 41-51 (doi: 10.1111/pce.13348)
- de Almeida Ribeiro P.R., dos Santos J.V., da Costa E.M., Lebbe L., Assis E.S., Louzada M.O., Guimaraes A.A., Willems A., de Souza Moreira F.M. Symbiotic efficiency and genetic diversity of soybean Bradyrhizobia in Brazilian soils. Agriculture, Ecosystems & Environment,2015, 212: 85-93 (doi: 10.1016/j.agee.2015.06.017).
- Thilakarathna M.S., Raizada M.N. A meta-analysis of the effectiveness of diverse rhizobia inoculants on soybean traits under field conditions. Soil Biology and Biochemistry, 2017, 105: 177-196 (doi: 10.1016/j.soilbio.2016.11.022).
- Zahran H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiology and Molecular Biology Reviews, 1999, 63(4): 968-989 (doi: 10.1128/MMBR.63.4.968-989.1999).
- Zeffa D.M., Fantin L.H., Koltun A., de Oliveira A., Nunes M., Canteri M.G., Gonçalves L. Effects of plant growth-promoting rhizobacteria on co-inoculation with Bradyrhizobium in soybean crop: a meta-analysis of studies from 1987 to 2018. PeerJ, 2020, 8: e7905 (doi: 10.7717/peerj.7905).
- Shrestha R., Adams C.B., Ravelombola W., MacMillan J., Trostle C., Ale S., Hinson P. Exploring phenotypic variation and associations in root nodulation, morphological, and growth character traits among 50 guar genotypes. Industrial Crops and Products, 2021, 171: 113831 (doi: 10.1016/j.indcrop.2021.113831).
- Salvagiotti F., Cassman K.G., Specht J.E., Walters D.T., Weiss A., Dobermann A. Nitrogen uptake, fixation and response to fertilizer N in soybeans: a review, Field Crops Research, 2008, 108(1): 1-13 (doi: 10.1016/j.fcr.2008.03.001).
- Habibi S., Ayubi A.G., Ohkama-Ohtsu N., Sekimoto H., Yokoyama T. Genetic characterization of soybean rhizobia isolated from different ecological zones in North-Eastern Afghanistan. Microbes and Environments, 2017, 32(1): 71-79 (doi: 10.1264/jsme2.ME16119).
- MacMillan J., Adams C.B., Trostle C., Rajan N. Testing the efficacy of existing USDA Rhizobium germplasm collection accessions as inoculants for guar. Industrial Crops and Products, 2021, 161: 113205 (doi: 10.1016/j.indcrop.2020.113205).