doi: 10.15389/agrobiology.2022.3.566rus
УДК 579.6:632.9
ФУНГИСТАТИЧЕСКАЯ АКТИВНОСТЬ ШТАММОВ Serratia proteamaculans И S. liquefaciens, ВЫДЕЛЕННЫХ В ГРЕЦИИ ИЗ ОТЛОЖЕНИЙ ГУАНО ЛЕТУЧИХ МЫШЕЙ В ПОДЗЕМНОЙ ПЕЩЕРЕ
G. MICHAIL1 ✉, A. REIZOPOULOU2, I. VAGELAS3
Представители рода Serratia вызывают повышенный научный интерес, поскольку они распространены практически повсеместно и проявляют эмульгирующие, поверхностно-активные, противообрастающие, противоопухолевые и противомикробные свойства. Вода — естественная среда обитания для нескольких видов серраций. В представляемой публикации мы сообщаем о первом выделении S. proteamaculans из гуано летучих мышей. Цель нашей работы заключалась в изучении гуано летучих мышей из подземной водной экосистемы на наличие изолятов Serratia, активных против фитопатогенных грибов. Штаммы Serratia, которые выделили из отложений гуано в пещере в регионе Фессалия (Эолия, Греция), первоначально были обозначены как Sl2 и Sl4, оказались способны ферментировать D-глюкозу, другие углеводы (D-маннит, D-маннозу) и сахарозу в качестве источников углерода и сахаров. Для обоих штаммов оптимальная температура для роста — 28 °С, при этом штамм Sl4 способен расти и при 4 °С. Штаммы бактерий Sl2 и Sl4 были отнесены к группе Serratia liquefaciens с помощью автоматического бактериологического анализатора VITEK® 2 («bioMerieux SA», Франция) и точно идентифицированы до вида с использованием MALDI-TOF MS («bioMerieux SA», Франция). MALDI-TOF MS классифицировала штамм Sl2 как S. proteamaculans, а штамм Sl4 — как S. liquefaciens. Насколько нам известно, это первое в мировой научной литературе сообщение о выделении и классификации S. proteamaculans из гуано летучих мышей. При 28 °С оба изученных штамма Serratia продуцировали продигиозин, при этом его оптимальную продукцию отмечали через 72 ч инкубации. Определение активности штаммов S. liquefaciens и S. proteamaculans против фитопатогенных грибов (Fusarium oxysporum, Alternaria alternata, Botrytis cinerea, Sclerotinia sclerotiorum и Rhizoctonia solani) in vitro показало способность изученных серраций продуцировать свободно диффундирующие соединения с фунгистатической активностью in vitro. Это первое сообщение о том, что штаммы S. liquefaciens и S. proteamaculans, выделенные из гуано летучих мышей, потенциально могут рассматриваться как биоагенты против фитопатогенных грибов. Исследование взаимодействия между фитопатогенными грибами и бактериями S. liquefaciens и S. proteamaculans подтвердили их эффективность для биоконтроля этих грибных патогенов.
Ключевые слова: Serratia spp., гуано летучих мышей, подземная водная среда, вторичные метаболиты, биоконтроль.
Род Serratia включает до 18 видов, которые можно найти в различных средах (1). Штаммы рода Serratia были выделены из воды, почвы, из растений и животных (2). У млекопитающих штаммы Serratia связаны с маститом у крупного рогатого скота, конъюнктивитом лошадей, септицемией у жеребят, коз и свиней, но также серрации были обнаружены у нескольких клинически здоровых людей (3). Сообщалось, что S. liquefaciens и S. marcescens являются условно-патогенными микроорганизмами для видов рукокрылых (4).
Группа Serratia liquefaciens включает виды S. liquefaciens, S. proteamaculans и S. grimesii (3). Штаммы группы S. liquefaciens преимущественно вызывают сепсис и инфекции кровотока через контаминированное клиническое оборудование и компоненты крови (5). В 1980 году в «Утвержденном списке названий бактерий» S. liquefaciens и S. proteamaculans были перечислены как отдельные виды (6). Хотя S. proteamaculans редко регистрируют в клинических образцах (вероятно, из-за невозможности легко различить виды в группе), было показано, что S. proteamaculans вызывает заболевания человека (7).
Вода — естественная среда обитания для нескольких видов серраций, включая S. marcescens, S. fonticola, S. grimesii, S. liquefaciens, S. plymuthica, S. rubidaea и S. ureilytica (8). S. marcescens, S. liquefaciens, S. proteamaculans, S. grimesii и S. plymuthica обнаружены в речной воде в одном исследовании, при этом преобладающим оказался вид S. marcescens, за которым следовал S. liquefaciens (3). S. marcescens subsp. sakuensis первоначально был выделен из взвеси сточных вод в очистных резервуарах в Японии (9). Виды Serratia вызывают серьезные болезни животных. S. marcescens был описан в 1958 году как причина заболевания, когда у части молочного стада диагностировали мастит. Есть много других сообщений о колонизации или патологиях, связанных с видами Serratia, у животных, включая рептилий, грызунов, птиц, цыплят, коз, свиней, рыб и лошадей (8).
Представители рода Serratia вызывают повышенный научный интерес, поскольку они проявляют эмульгирующие, поверхностно-активные, противообрастающие, противоопухолевые и противомикробные свойства (10, 11). Публикации показывают, что способность этого рода к обитанию практически во всех средах обусловлена синтезом многочисленных внеклеточных продуктов, включая экзоферменты, нуклеазы и вторичные метаболиты, которые помогают в адаптации Serratia к суровым условиям окружающей среды (12). К таким соединениям относится пигмент продигиозин, продуцируемый некоторые штаммы S. marcescens, S. rubidaea и S. surfactantfaciens и проявляющий антибактериальную и антимикотическую активность (2, 10).
Фитопатогенные грибы вызывают серьезные потери в сельском хозяйстве во всем мире. Для борьбы с распространением грибных болезней растений эффективны химические фунгициды, которые применяются в отношении ряда плодовых и других сельскохозяйственных культур. Растущая угроза устойчивости к фунгицидам, например развитие резистентности у возбудителя серой гнили Botrytis cinerea, хорошо известна. Гриб B. cinerea печально известен как организм с высоким риском быстрого развития резистентности, и внедрение новых классов фунгицидов для борьбы с серой гнилью всегда сопровождалось появлением резистентности в полевых популяциях (13). В этих условиях возрастает потребность в альтернативах фунгицидам для профилактики и лечения инфекций.
Основная цель нашего исследования заключалась в изучении гуано летучих мышей из подземной водной экосистемы на наличие изолятов Serratia, активных против фитопатогенных грибов.
Методика. Пещера Малаки, в которой были отобраны образцы гуано, расположена в Фессалии (Эолия, Греция; 48°28′36″ с.ш., 20°29′09″ в.д., высота 339 м над уровнем моря).
Грамотрицательные бактерии выделяли из гуано летучих мышей. Для рутинного выделения применили метод квадрантного штриха. Использовали NA (питательный агар), картофельно-декстрозный агар (PDA) и агар MacConkey (MCA) («Oxoid Limited», Великобритания), чашки с агаром инкубировали при 22 °С в течение 2 сут (1).
Для идентификации бактериальных изолятов до уровня видов использовали автоматический бактериологический анализатор VITEK® 2 («bio-Merieux SA», Франция) и систему MALDI-TOF MS («bioMerieux SA», Франция) в соответствии с описаниями (14, 15). Отдельные колонии отбирали на скошенном агаре и суспендировали в 2,5 мл 0,45 % стерильного физиологического раствора, доводя бактериальную суспензию до стандарта мутности МакФарланда 0,5 с помощью денситометра («bioMerieux SA», Франция) в соответствии с инструкциями производителя. Бактериальные взвеси готовили в течение 30 мин после внесения в кассету для идентификации грамотрицательных (GN) бактерий с использованием автоматического анализатора VITEK® 2. Анализ проводили с помощью программы VITEK® 2 v. 07.01 («bioMerieux SA», Франция) (15), вероятность выше 80 % при соответствующем профиле считали достаточной при идентификации в пределах перечня микроорганизмов, определяемых до видов или родов (таксонов) с помощью VITEK® 2 (14, 15).
Перед измерением спектров MALDI-TOF MS бактериальные изоляты пересевали на РСА («Oxoid Limited», Великобритания) и культивировали в течение 24 ч при 28 °С. При получении масс-спектров следовали общему протоколу прямого переноса (обычно называемому измерением целых, или интактных, клеток). Для этого примерно 0,1 мг клеточного материала переносили непосредственно из бактериальной колонии или мазка колоний в целевое пятно MALDI. После высушивания при комнатной температуре на пятна образцов наносили 1 мкл матричного раствора (10 мг/мл α-циано-4-гидроксикоричной кислоты в 50 % ацетонитриле и 2,5 % трифторуксусной кислоте). Для определения воспроизводимости генерации масс-спектров все культуры выращивали независимо 4-кратно (биологические повторности), каждое измерение выполняли трижды (аналитические повторности). Анализ проводили на масс-спектрометре Autoflex MALDI-TOF («Bruker Daltonics», Германия) с применением Flex Frontiers (16), дерепликацию — в программе Recurrent Bacterial Isolates Control 3.4 («Bru-ker Daltonics», Германия), калибровку — с помощью Bacterial Test Standard («Bruker Daltonics», Германия) (http://www.bruker.com/jp/products/mass-spectrometry-andseparations/literature/literatureroom.html?eID=dam_frontend_ push&stream=1&docID=58883). Все масс-спектры были измерены автоматически с использованием программного обеспечения Flex Control («Bruker Corporation», США, Германия) в соответствии со стандартным методом измерения для микробной идентификации. В частности, наши установочные значения в линейном положительном режиме были следующими: напряжение 1-го ионного источника 20 кВ, 2-го — 19 кВ; напряжение на линзе 6,5 кВ; диапазон масс 2-20 кДа; окончательный спектр представлял собой сумму 10 отдельных спектров, каждый из которых был получен с помощью 200 лазерных выстрелов в случайных положениях пятна мишени. В MALDI-TOF MS, с помощью которой преимущественно генерируются и обнаруживаются ионы +1, значения Da получали на основании соотношения m/z. Для классификации бактерий использовали программное обеспечение BioTyper 3.1 («Bruker Daltonics», Германия), библиотеку спектров MBT 6903 MPS (выпущена в апреле 2016 года), стандартный метод предварительной обработки MALDI Biotyper и стандартный метод идентификации MALDI Biotyper MSP, скорректированные производителем («Bruker Dal-tonics», Германия) (14, 15).
Для выявления принадлежности штаммов Serratia proteamaculans (Sl2) и Serratia liquefaciens (Sl4) к группе Serratia liquefaciens с помощью системы VITEK® 2 («bioMerieux SA», Франция) использовали идентификационную карту VITEK® 2 GN (17).
Штаммы Serratia (Sl2, Sl4), выделенные из гуано летучих мышей и идентифицированные методом MALDI-TOF MS как S. proteamaculans (штамм Sl2) и S. liquefaciens (штамм Sl4), подвергали скринингу на продукцию продигиозина на среде MacConkey. Чашки инкубировали при 28 °С в течение 24, 48 и 72 ч и проверяли на наличие колоний Serratia с гиперпигментацией. Штаммы, продуцирующие пигмент (гиперпигментация), обнаруживали по появлению розово-красного окрашивания.
Для оценки противогрибных свойств штаммов серраций использовали пять штаммов грибов, вызывающих болезни растений. Штаммы Fusarium oxysporum, Alternaria alternata, Botrytis cinerea, Sclerotinia sclerotiorum и Rhizoctonia solani были ранее выделены из пораженных растений томата в Центральной Греции (18, 19) согласно описанию (20, 21).
Для скрининга in vitro противогрибной активности выделенных штаммов Serratia (S. liquefaciens и S. proteamaculans) использовали штаммы F. oxysporum (BFI 2550), A. alternata (BFI 2596), B. cinerea (BFI 1952), S. scle-rotiorum (BFI 2529) и R. solani (BFI 2531), полученные из Benaki Phytopathological Institute (BFI). Применяли метод двойного культивирования на среде PDA (22). В чашке Петри с PDA в течение 24 ч выращивали культуры бактерий в виде 40-миллиметрового штриха с началом на расстоянии 30 мм от центра чашки. В центр чашки с тестируемым бактериальным штаммом помещали агаровый блок диаметром 5 мм с 5-суточной культурой гриба. Чашки инкубировали при 25 °С в течение 5 сут, ежедневно контролировали зону ингибирования. Измеряли мицелиальный радиальный рост фитопатогенов (диаметр), анализировали особенности проявления антифунгальной активности.
Данные анализировали с использованием статистического пакета Minitab (https://www.minitab.com/en-us/support/downloads/). Для оценки антагонистического эффекта использовали дисперсионный анализ, результаты представлены в виде графика Excel. Приведены средние (M) и стандартные ошибки различий между средними (±SEM).
Результаты. Место отбора образов гуано летучих мышей представляет собой небольшую известняковую подземную пещеру (рис. 1). Пещера Малаки делится на три зоны: 1) сумеречная зона у входа, 2) средняя зона с недостаточным освещением и переменной температурой и 3) глубокая зона полной темноты и постоянной температуры (17 °С) на протяжении все-го года. В глубокой зоне пещеры есть небольшое подземное озеро и несколько крупных отложений гуано летучих мышей. В фауне глубокой зоны в основном были обнаружены виды родов Minio-pterus, Myotis и Rhinolophus (23).
Штаммы Serratia, первоначально обозначенные как Sl2 и Sl4, были способны ферментировать D-глюкозу, использовать мальтозу, D-маннит, D-маннозу и трегалозу, другие углеводы, сахарозу как источники углерода и сахара (табл. 1). У обоих штаммов оптимальная температура для роста составляла 28 °С, при этом штамм Sl4 оказался способен расти и при 4 °С.
Все бактериальные штаммы с палочковидной формой клеток, способные к росту на агаре MacConkey, были идентифицированы до вида (с вероятностью 99,9 %) с помощью системы VITEK® 2 и карты VITEK 2 GN ID (см. табл. 1). В соответствии с идентификационной картой VITEK 2 GN оба штамма (Sl2 и Sl4) классифицированы как группа Serratia liquefaciens. Время идентификации на автоматическом анализаторе составляло от 4,17 до 4,65 ч для всех изолятов.
С использованием MALDI-TOF MS все штаммы, идентифицированные с помощью системы VITEK® 2 как группа Serratia liquefaciens, были классифицированы до вида (с доверительным интервалом 99,9 %) как S. proteamaculans (Sl2) и S. liquefaciens (Sl4). Величина доверительного интервала 99,9 % свидетельствует о четком различии и присутствии указанных штаммов — S. proteamaculans и S. liquefaciens.
Идентификация микроорганизмов с помощью MADLI-TOF основана на применении четырех коммерческих систем и их баз данных: 1) MALDI Biotyper («Bruker Daltonics», Германия); 2) система Spectral ARchive and Microbial Identification System (SARAMIS™) («AnagnosTec», Германия); 3) Andromas («Andromas», Франция); 4) Vitek MS («bioMerieux SA», Франция). Каждая из систем включает в себя прибор MALDI-TOF от «Bru-ker Daltonics» или «Shimadzu Corporatioт» (Япония), а наиболее часто устанавливаемыми в обычных лабораториях системами являются MALDI Biotyper и Vitek MS, которые служат платформами, одобренными FDA (24). Системы различаются базами данных, алгоритмами идентификации и инструментарию (25). В рутинной лабораторной практике MALDI-TOF используется для идентификации бактерий или грибов из колоний, выращенных на твердых средах. До сих пор эта процедура использовалась для идентификации грамотрицательных и положительных палочек, грамположительных кокков, требовательных организмов, таких как микобактерии, нокардии и другие актиномицеты, анаэробных бактерий, дрожжей и мицелиальных грибов (26-29). По имеющимся данным, показатели идентификации рода чрезвычайно высоки (97-99 %) и колеблются от 85 % до 97 % на уровне видов (30-35).
Оба микроорганизма (S. liquefaciens и S. proteamaculans) ферментировали лактозу на среде MacConkey (рис. 2, А). S. liquefaciens способна ферментировать лактозу быстрее, чем S. proteamaculans. Колонии S. liquefaciens круглые, гладкие, с ровными краями, выпуклые, среднего размера, блестяще-красные и непрозрачные, тогда как колонии S. proteamaculans круглые, нитевидные, выпуклые, мелкие, оранжево-красные и непрозрачные (см. рис. 2, Б). Красновато-оранжевая окраска колоний (оранжевый ореол) свидетельствует о синтезе пигмента продигиозина (см. рис. 2, Б). Его оптимальную продукцию отмечали при температуре 28 °С и длительности инкубации 72 ч (табл. 2).
Изоляты S. proteamaculans и S. liquefaciens продемонстрировали значительный антагонизм по отношению к F. oxysporum, A. alternata, B. cinerea, S. sclerotiorum и R. solani и в значительной степени ограничивают рост колоний этих фитопатогенов (рис. 3).
Следует отметить высокую фунгистатическую активность в отношении мицелия A. alternate, выявленную нами у выделенных из гуано летучих мышей штаммов S. liquefaciens и S. proteamaculans.
В этом исследовании мы взяли образцы гуано летучих мышей из разных мест подземной карстовой водной экосистемы, которая служит местом гнездования нескольких видов летучих мышей. Выделенные бактерии выращивали на питательном агаре, PDA и агаре MacConkey. В образцах с помощью системы VITEK® 2 мы идентифицировали бактерии группы Serratia liquefaciens. Применив типирование методом времяпролетной масс-спектрометрии с лазерной десорбцией/ионизацией с использованием матрицы (MALDI-TOF MS) мы смогли классифицировать изоляты группы S. liquefaciens как S. liquefaciens и S. proteamaculans. MALDI-TOF MS — простая в использовании, быстрая, надежная, экономичная и безвредная для окружающей среды методология, которая произвела революцию в идентификации патогенов и обнаружении чувствительности/резистентности к противомикробным препаратам в клинических условиях (36). На агаре Mac-Conkey все штаммы после 48 ч инкубации продуцировали пигмент продигиозин, как описано и в других исследованиях (37).
Между штаммами, классифицированными как группа Serratia liquefaciens, имелись четыре различия в фенотипических характеристиках (см. табл. 1). Штамм S. proteamacula характеризовалсяналичием альфа-глюкозидазы, палатинозы, гамма-глутамила и фосфатазы, тогда как у S. liquefaciens эти свойства отсутствовали. Наши данные показывают, что MALDI-TOF MS, если необходимо, может использоваться для идентификации видов в группе S. liquefaciens, как это применялось ранее при вспышках внутрибольничных инфекций с распространением других видов Serratia (14, 38). В нашем исследовании мы изолировали из гуано летучих мышей сходные виды бактерий, обозначенные как группа Serratia liquefaciens. S. liquefaciens и S. marcescens были описаны как условно-патогенные микроорганизмы для многих видов европейских летучих мышей (4), но до сих пор не было получено убедительных доказательств присутствия S. proteamaculans ни в гуано летучих мышей, ни в составе их бактериальной микрофлоры. Насколько нам известно, в своих работах мы первые сообщаем о выявлении S. proteamaculans в гуано летучих мышей.
С момента первого применения в целях идентификации микроорганизмов (39, 40) MALDI-TOF MS предлагали в качестве многообещающей альтернативы для дерепликации бактериальных изолятов (41-43). MALDI-TOF MS использовалась как эффективная альтернатива секвенированию гена 16S рРНК (44-48). Идентификация микроорганизмов методом MALDI-TOF MS включает генерацию масс-спектров (из интактных клеток или внутриклеточного содержимого) с последующим сопоставлением с известными ссылками в базе данных (49, 50).
M.M. Newman с соавт. (51) в своих недавних исследованиях в пещерах Нью-Мексико (США) не представили доказательств выделения или идентификации каких-либо видов Serratia. Их исследование было основано на выделении бактерий на среде TSA, кровяном агаре и гуано летучей мыши (BGM) и идентификации с использованием ПЦР-амплификации гена 16S рРНК. Эти авторы исследовали как свежее, так и разлагающееся гуано летучих мышей и обнаружили разные бактериальные таксоны, но не виды Serratia (51). Наши данные указывают на то, что у европейских (4, 52, 53), индийских (54, 55) и американских (51) континентальных летучих мышей микробиомы и/или гуано могут различаться по составу бактериальных штаммов. Как европейские, так и индийские исследования гуано местных летучих мышей выявили присутствие Serratia spp., но в микробиоме летучих мышей из Америки серрации отсутствовали.
Новый штамм Serratia — Serratia marcescens B4A продуцирует сильнодействующие антимикотические соединения и подавляет развитие насекомых и патогенов растений, например Rhizoctonia solani и Alternaria raphanin (56). Serratia spp. также отвечают за продукцию вторичных метаболитов — сидерофоров и фитогормонов и защищают растения от инфекций, вызванных патогенами (56). Некоторые штаммы Serratia синтезируют галогенированный вторичный метаболит пирролнитрин, который рассматривают как перспективный сельскохозяйственный фунгицид (56, 57). Виды Serratia также продуцируют биологически активные соединения и ферменты — нуклеазы, хитиназы, липазы, протеазы, амилазы, серрализин и гемолизин (56). Существует довольно много исследований, посвященных важной роли видов Serratia в качестве агентов биоконтроля на клубнике, цветной капусте и оливках. S. plymuthica A30 обладает активностью в отношении патогенных бактерий Dickeya solani, вызывающих черную ножку и мягкую гниль картофеля (58-60).
В нашем исследовании оба штамма Serratia — S. liquefaciens и S. proteamaculans были способны продуцировать красный пигмент продигиозин и проявлять значительную активность против F. oxysporum, A. alternata, B. cin-erea, S. sclerotiorum и R. solani. Сообщается, что соединение продигиозина, биологическая роль которого четко не выяснена, продуцируют многие виды Serratia. Продигиозин (2-метил-3-пентил-6-метоксипродигинин) представляет собой гетероциклический вторичный метаболит красного цвета, принадлежащий к классу трипиррольных соединений (61). Продигиозин появляется на более поздних стадиях роста бактерий, действуя как избыточная продукция вторичных метаболитов с антимикробной активностью широкого спектра действия (11). Биосинтез продигиозина контролируется многими факторами окружающей среды и физико-химическими факторами, включая температуру, содержание кислорода и рН, при этом максимальные выходы продукции достигаются в отсутствие света (61). Другие виды рода Serratia, например S. plymuthica, S. rubidaea и S. nematodiphila, также способны продуцировать недиффундирующий красный пигмент продигиозин как вторичный метаболит (10). Сообщалось, что продигиозин проявляет противомалярийную, антибактериальную, антимикотическую, противопротозойную, противоопухолевую и иммунодепрессивную активность (62). Наши данные — это первое сообщение о том, что S. liquefaciens и S. pro-teamaculans, выделенные из гуано летучих мышей, демонстрируют явную и отчетливую противогрибную (фунгистатическую) активность.
Таким образом, наши исследования предоставили новые данные о наличии изолятов Serratia proteamaculans и S. liquefaciens в гуано летучих мышей. Нами также получена новая информация о присутствии S. proteamaculans в гуано летучих мышей из подземной водной экосистемы. Показано, что изоляты S. proteamaculans и S. liquefaciens способны продуцировать продигиозин. Небольшое количество бактериальных клеток может обладать высокой конкурентоспособностью благодаря соединениям, которые диффундируют в агар и, в частности, проявляют фунгистатические свойства в отношении патогенов растений. Перспективы применения продигиозина и других вторичных метаболитов серраций в качестве сельскохозяйственных фунгицидов требует дальнейшего изучения.
1. VITEK® 2 GN ID карта быстрой идентификации грамотрицательных бактерий, выделенных из отложений гуано летучих мышей (пещера Малаки, Фессалия, Эолия, Греция; 48°28′36″ с.ш., 20°29′09″ в.д.) |
||||
Признак |
Обозначение |
Количество |
Штамм Serratia |
|
Sl2 |
Sl4 |
|||
Ala-Phe-Pro-arylamidase |
APPA |
0,0384 мг |
- |
- |
Adonitol |
ADO |
0,1875 мг |
- |
- |
L-pyrrolydonyl arylamidase |
PyrA |
0,018 мг |
+ |
+ |
L-Arabitol |
1ARL |
0,3 мг |
- |
- |
D-Cellobiose |
dCEL |
0,3 мг |
- |
- |
Beta-galactosidase |
BGAL |
0,036 мг |
+ |
+ |
H2S |
H2S |
0,0024 мг |
- |
- |
Beta-N-acetyl-glucosaminidase |
BNAG |
0,0408 мг |
+ |
+ |
Glutamyl arylamidase pNA |
AG LTp |
0,0324 мг |
- |
- |
D-glucose |
dGLU |
0,3 мг |
+ |
+ |
Gamma-glutamyl-transferase |
GGT |
0,0228 мг |
+ |
- |
Fermentation/glucose |
OFF |
0,45 мг |
+ |
+ |
Beta-glucosidase |
BGLU |
0,036 мг |
+ |
+ |
D-maltose |
dMAL |
0,3 мг |
- |
- |
D-mannitol |
dMAN |
0,1875 мг |
+ |
+ |
D-mannose |
dMNE |
0,3 мг |
+ |
+ |
Beta-xylosidase |
BXYL |
0,0324 мг |
- |
- |
Beta-alanine arylamidase pNA |
BAlap |
0,0174 мг |
- |
- |
L-proline arylamidase |
ProA |
0,0234 мг |
+ |
+ |
Lipase |
LIP |
0,0192 мг |
- |
- |
Palatinose |
PLE |
0,3 мг |
+ |
- |
Tyrosine arylamidase |
TyrA |
0,0276 мг |
+ |
+ |
Urease |
URE |
0,15 мг |
- |
- |
D-sorbitol |
dSOR |
0,1875 мг |
+ |
+ |
Saccharose/sucrose |
SAC |
0,3 мг |
+ |
+ |
D-tagatose |
dTAG |
0,3 мг |
- |
- |
D-trehalose |
dTRE |
0,3 мг |
+ |
+ |
Citrate (sodium) |
CIT |
0,054 мг |
+ |
+ |
Malonate |
MNT |
0,15 мг |
- |
- |
5-keto-D-gluconate |
5KG |
0,3 мг |
+ |
+ |
[-Lactate alkalinisation |
1LATk |
0,15 мг |
+ |
+ |
Alpha-glucosidase |
AGLU |
0,036 мг |
+ |
- |
Succinate alkalinisation |
SUCT |
0,15 мг |
+ |
+ |
Beta-N-acetyl-galactosaminidase |
NAGA |
0,0306 мг |
+ |
+ |
Alpha-galactosidase |
AGAL |
0,036 мг |
- |
+ |
Phosphatase |
PHOS |
0,0504 мг |
+ |
- |
Glycine arylamidase |
GIyA |
0,012 мг |
- |
N/A |
Ornithine decarboxylase |
ODC |
0,3 мг |
+ |
+ |
Lysine decarboxylase |
LDC |
0,15 мг |
+ |
+ |
Decarboxylase Base |
ODEC |
N/A |
|
|
L-histidine assimilation |
1H1Sa |
0,087 мг |
- |
- |
Coumarate |
CMT |
0,126 мг |
+ |
+ |
Beta-glucoronidase |
BGUR |
0,0378 мг |
- |
- |
O/129 resistance (comp. vibrio) |
0129R |
0,0105 мг |
+ |
+ |
Glu-Gly-Arg-arylamidase |
GGAA |
0,0576 мг |
+ |
+ |
L-malate assimilation |
1MLTa |
0,042 мг |
- |
- |
ELLMAN |
ELLM |
0,03 мг |
- |
- |
[-Lactate assimilation |
1LATa |
0,186 мг |
- |
- |
Примечание. «-» — отсутствие признака, «+» — наличие признака, N/A (not applicable) — данные отсутствуют. Для Sl2 приведены данные, представленные в работе Michail G., Reizopoulou A., Vagelas I. First report of Serratia species isolated from subterranean cave aquatic environment. InternationalResearchJournalofEngineeringandTechnology, 2020, 07(12): 1776-1780. |
||||
2. Влияние времени инкубации на продукцию продигиозина штаммами Serratia, выделенными из отложений гуано летучих мышей в Греции |
|||
Штамм |
24 ч |
48 ч |
72 ч |
S. proteamaculans |
+/- |
++ |
+++ |
S. liquefaciens |
+/- |
+ |
++ |
Примечание. Штаммы культивировали на агаре MacConkey при 28 °С; +/- — очень низкая продукция пигмента, + — низкая продукция пигмента, ++ — средняя интенсивность пигментации, +++ — высокий уровень продукции пигмента. |
|||
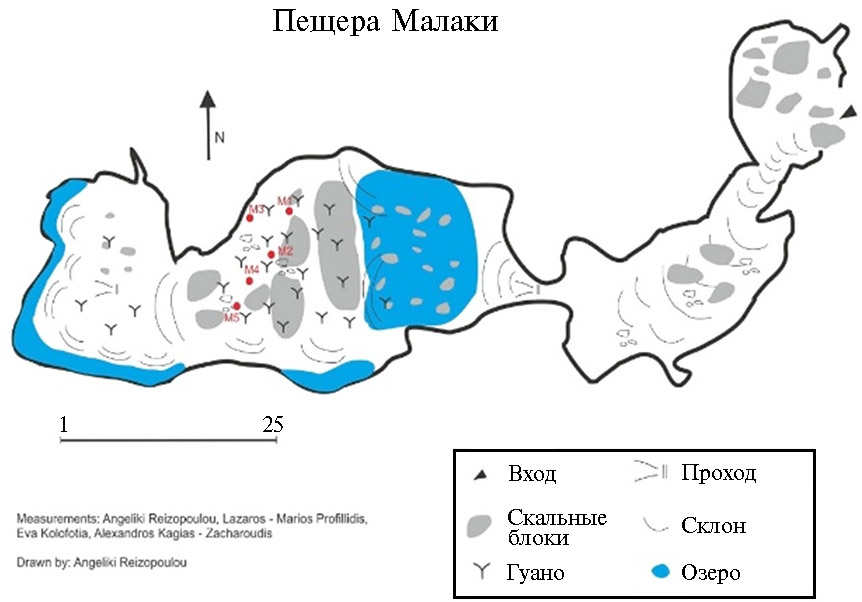
Рис. 1. Карта известняковой пещеры Малаки в Фессалии (Эолия, Греция; 48°28′36″ с.ш., 20°29′09″ в.д., высота 339 м над уровнем моря). Красными точками обозначены места отбора проб, расположенные рядом с гнездами местных летучих мышей. Рисунок выполнен Angeliki Reizopoulou.
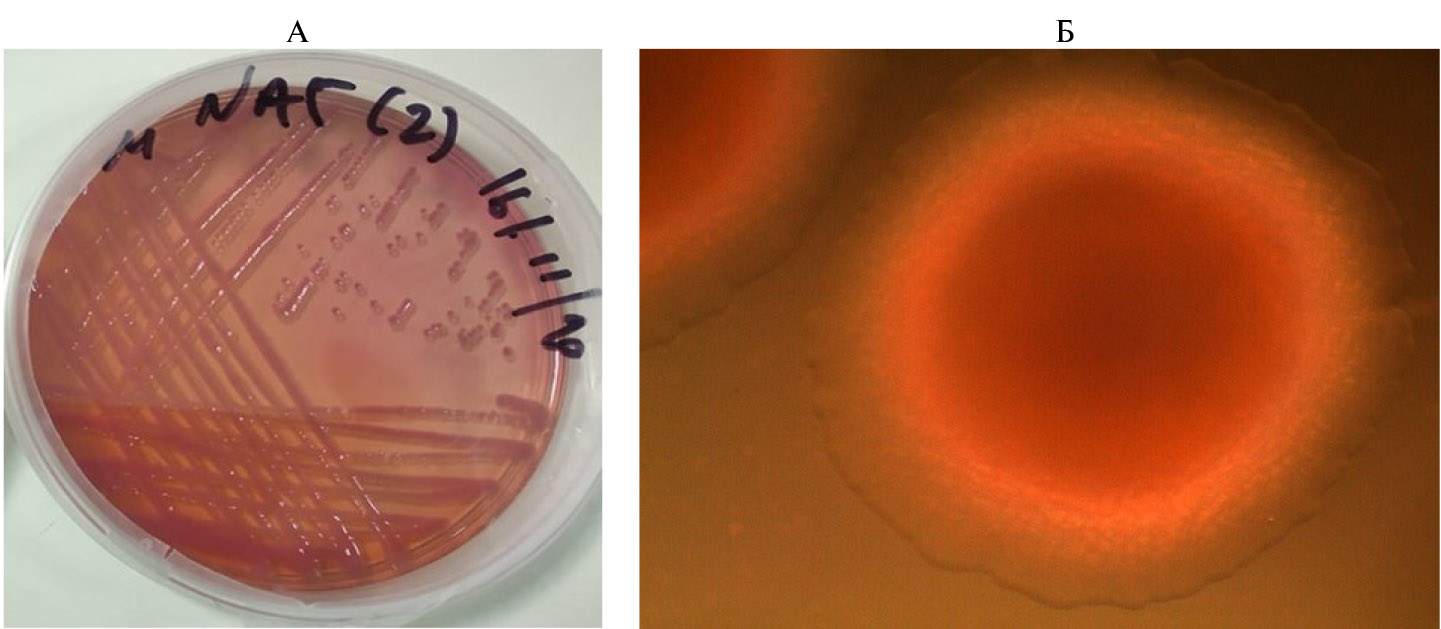
Рис. 2. Рост бактерий группы Serratia liquefaciens, выделенных из отложений гуано летучих мышей (пещера Малаки, Фессалия, Эолия, Греция; 48°28′36″ с.ш., 20°29′09″ в.д.), при инкубации при 28 °С в течение 72 ч на агаре MacConkey: А — ферментация лактозы, Б — пигментация (продукция продигиозина).
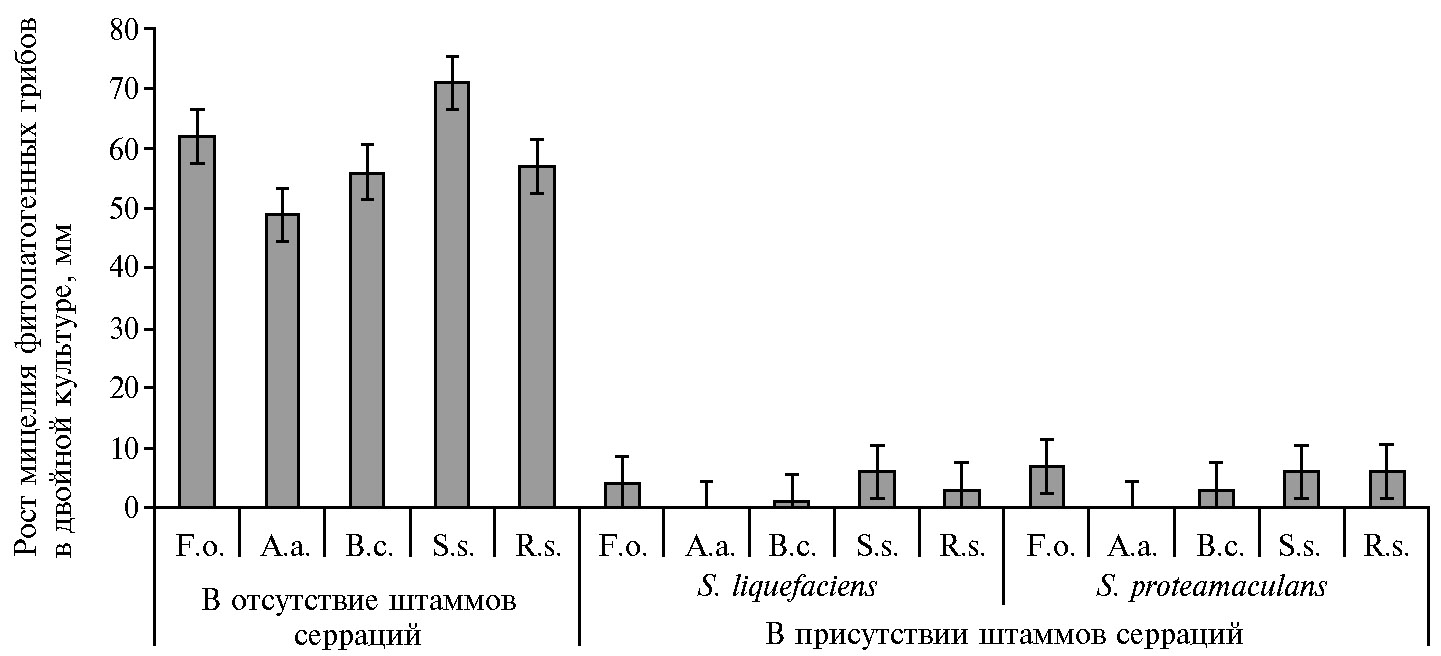
Рис. 3. Рост мицелия (диаметр, мм) фитопатогенных грибов Fusarium oxysporum (F.o.), Alternaria alternata (A.a), Botrytis cinerea(B.c.), Sclerotinia sclerotiorum(S.s.) и Rhizoctonia solani (R.s.) в двойных культурах in vitro, под влиянием S. liquefaciens и S. proteamaculans, выделенных из отложений гуано летучих мышей в Греции (n = 17, M±SEM; агар PDA, 25 °С, 5 сут).
ЛИТЕРАТУРА
- García-Fraile P., Chudíčková M., Benada O., Pikula J., Kolařík M. Serratia myotis sp. nov. and Serratia vespertilionis sp. nov., isolated from bats hibernating in caves. International Journal of Systematic and Evolutionary Microbiology, 2015, 65(Pt_1): 90-94 (doi: 10.1099/ijs.0.066407-0).
- Grimont F., Grimont P.A.D. The genus Serratia. In: The prokaryotes: a handbook on the biology of bacteria, 3rd edn. /M. Dworkin, S. Falkow, E. Rosenberg, K.H. Schleifer, E. Stackebrandt (eds.). New York, Springer, 2006: 219-244 (doi: 10.1007/0-387-30746-X_11).
- Grimont F., Grimont P.A.D. Genus XXXIV. Serratia Bizio 1823, 288AL. In: Bergey's manual of systematic bacteriology, 2nd edn., vol. 2, part B/D.J. Brenner, N.R. Krieg, J.T. Staley (eds.), Springer Science and Business Media, New York, NY, 2005: 799-811.
- Mühldorfer K., Speck S., Kurth A., Lesnik R., Freuling C., Müller T., Kramer‐Schadt S., Wibbelt G. Diseases and causes of death in European bats: dynamics in disease susceptibility and infection rates. PLoS ONE, 2011, 6: e29773 (doi: 10.1371/journal.pone.0029773).
- Stock I., Grueger T., Wiedemann B. Natural antibiotic susceptibility of strains of Serratia marcescens and the S. liquefaciens complex: S. liquefaciens sensu stricto, S. proteamaculans and S. grimesii. International Journal of Antimicrobial Agents, 2003, 22(1): 35-47 (doi: 10.1016/s0924-8579(02)00163-2).
- Skerman V.B.D., McGowan V., Sneath, P.H.A. Approved lists of bacterial names. International Journal of Systematic and Evolutionary Microbiology, 1980, 30(1): 225-420 (doi: 10.1099/00207713-30-1-225).
- Bollet C., Grimont P., Gainnier M., Geissler A., Sainty J.M., De Micco P. Fatal pneumonia due to Serratia proteamaculans subsp. quinovora. Journal of Clinical Microbiology, 1993, 31(2): 444-445 (doi: 10.1128/jcm.31.2.444-445.1993).
- Mahlen S.D. Serratia infections: from military experiments to current practice. Clinical Microbiology Reviews, 2011, 24(4): 755-791 (doi: 10.1128/CMR.00017-11).
- Ajithkumar B., Ajithkumar V.P., Iriye R., Doi Y., Sakai T. Spore-forming Serratia marcescens subsp. sakuensis subsp. nov., isolated from a domestic wastewater treatment tank. International Journal of Systematic and Evolutionary Microbiology, 2003, 53(1): 253-258 (doi: 10.1099/ijs.0.02158-0).
- Su C., Xiang Z., Liu Y., Zhao X., Sun Y., Li Z., Li L., Chang F., Chen T., Wen X., Zhou Y., Zhao F. Analysis of the genomic sequences and metabolites of Serratia surfactantfaciens sp. nov. YD25T that simultaneously produces prodigiosin and serrawettin W2. BMC Genomics, 2016, 17: 865 (doi: 10.1186/s12864-016-3171-7).
- Clements T., Ndlovu T., Khan W. Broad-spectrum antimicrobial activity of secondary metabolites produced by Serratia marcescens strains. Microbiological Research, 2019, 229: 126329 (doi: 10.1016/j.micres.2019.126329).
- Harris A.K.P., Williamson N.R., Slater H., Cox A., Abbasi S., Foulds I., Simonsen H.T., Leeper F.J., Salmond G.P.C. The Serratia gene cluster encoding biosynthesis of the red antibiotic, prodigiosin, shows species- and strain-dependent genome context variation. Microbiology, 2004, 150(11): 3547-3560 (doi: 10.1099/mic.0.27222-0).
- Hahn M. The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study. Journal of Chemical Biology, 2014, 7(4): 133-141 (doi: 10.1007/s12154-014-0113-1).
- Rodel J., Mellmann A., Stein C., Alexi M., Kipp F., Edel B., Dawczynski K., Brandt C., Seidel L., Pfister W., Löffler B., Straube E. Use of MALDI-TOF mass spectrometry to detect nosocomial outbreaks of Serratia marcescens and Citrobacter freundii. European Journal of Clinical Microbiology & Infectious Diseases, 2019, 38: 581-591 (doi: 10.1007/s10096-018-03462-2).
- Moehario LH., Tjoa E., Putranata H., Joon S., Edbert D., Robertus T. Performance of TDR-300B and VITEK®2 for the identification of Pseudomonas aeruginosa in comparison with VITEK®-MS. Journal of International Medical Research,2021, 49(2): 30006052198989 (doi: 10.1177/0300060521989893).
- Strejcek Μ., Smrhova Τ., Junkova P., UhlikFront O. Whole-cell MALDI-TOF MS versus 16S rRNA gene analysis for identification and dereplication of recurrent bacterial isolates. Frontiers in Microbiology, 2018, 9: 1294 (doi: 10.3389/fmicb.2018.01294).
- Crowley E., Bird P., Fisher K., Goetz K., Boyle M., Benzinger M.J., Jr, Juenger M., Agin J., Goins D., Johnson R., Collaborators. Evaluation of the VITEK 2 Gram-Negative (GN) Microbial Identification Test Card: collaborative study. Journal of AOAC International, 2012, 95(3): 778-785 (doi: 10.5740/jaoacint.CS2011_17).
- Vagelas I., Sugar I.R. Potential use of olive mill wastewater to control plant pathogens and post harvest diseases. Carpathian Journal of Food Science & Technology, 2020, 12(4): 140-145 (doi: 10.34302/crpjfst/2020.12.4.14).
- Vagelas I.K., Giurgiulescu L. Bioactivity of olive oil mill wastewater against grey mould disease. Carpathian Journal of Food Science & Technology, 2019, 11(4): 161-164 (doi: 10.34302/2019.11.4.15).
- Vagelas I.K. Efficacy of Pseudomonas oryzihabitans as a biocontrol agent of root pathogens. Thesis (PhD). University of Reading, UK, 2002.
- Vagelas I., Kalorizou H., Papachatzis A., Botu M. Bioactivity of olive oil mill wastewater against plant pathogens and post-harvest diseases. Biotechnology & Biotechnological Equipment, 2009, 23(2): 1217-1219 (doi: 10.1080/13102818.2009.10817641).
- Agarry O.O., Akinyosoye F.A., Adetuyi F.C. Antagonistic properties of microogranisms associated with cassava (Manihot esculenta, Crantz) products. African Journal of Microbiology, 2005, 4(7): 627-632 (doi: 10.5897/AJB2005.000-3114).
- Georgiakakis P., Papadatou E. Chiroptera. In: Deliverable 7, C-Phase of Study 7: “Monitoring and evaluation of conservation status mammal species of Community interest in Greece” /G. Papamichael, T. Arapis, K. Petkidi, I. Fytou, V. Chatirvasanis. Ministry of Environment, Energy and Climatic Change, Athens, Scholars Partnership and Consultancy Firms “ARAPIS THOMAS EYAGGELOY, GEOANALYSI S.A. and PAPACHARISI ALEXANDRA THEODORA”, Athens. 2015 (in Greek).
- Posteraro B., De Carolis E., Vella A., Sanguinetti M. MALDI-TOF mass spectrometry in the clinical mycology laboratory: identification of fungi and beyond. Expert Review of Proteomics, 2013, 10(2): 151-164 (doi: 10.1586/epr.13.8).
- Caroll K., Patel R. Systems for identification of bacteria and fungi. In: Manual of clinical microbiology, 11th edition /J.H. Jorgensen, K.C. Carroll, G. Funke, M.A. Pfaller, M.L. Landry, S.S. Richter, D.W. Warnock (eds.). ASM Press N.W., Washington, 2015: 29-43 (doi: 10.1128/9781555817381.ch4).
- Nomura F. Proteome-based bacterial identification using matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS): A revolutionary shift in clinical diagnostic microbiology. Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics, 2015, 1854(6): 528-537 (doi: 10.1016/j.bbapap.2014.10.022).
- Ford B.A., Burham C.A. Optimization of routine identification of clinically relevant gram-negative bacteria by use of matrix-assisted laser desorption ionization-time of flight mass spectrometry and the Bruker Biotyper. Journal of Clinical Microbiology, 2013, 51(5): 1412-1420 (doi: 10.1128/JCM.01803-12).
- Hotta Y., Sato J., Sato H., Hosoda A., Tamura H. Classification of the genus Bacillus based on MALDI-TOF MS analysis of ribosomal proteins coded in S10 and spc operons. Journal of Agricultural and Food Chemistry, 2011, 59(10): 5222-5230 (doi: 10.1021/jf2004095).
- Seng P., Drancourt M., Couriet F., La Scola B., Fournier P-E., Rolain J.M., Raoult D. Ongoing revolution in bacteriology: routine identification of bacteria by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clinical Infectious Diseases, 2009, 49(4): 543-551 (doi: 10.1086/600885).
- Cherkaoui A., Hibbs J., Emonet S., Tangomo M., Girard M., Francois P., Schrenzel J. Comparison of two matrix-assisted laser desorption ionization-time of flight mass spectrometry methods with conventional phenotypic identification for routine identification of bacteria to the species level. Journal of Clinical Microbiology, 2010, 48(4): 1169-1175 (doi: 10.1128/jcm.01881-09).
- Marko D.C., Saffeert R.T., Cunningham S.A., Hyman J., Walsh J., Arbefeville S., Howard W., Pruessner J., Safwat N., Cockerill F.R., Bossier A.D., Patel R., Richter S.S. Evaluation of the Bruker Biotyper and Vitek MS matrix-assisted laser desorption ionization-time of flight mass spectrometry systems for identification of nonfermenting gram-negative bacilli isolated from cultures from cystic fibrosis patients. Journal of Clinical Microbiology, 2012, 50(6): 2034-2039 (doi: 10.1128/jcm.00330-12).
- Mancini N., De Carolis E., Infurmari L., Vella A., Clementi N., Vaccaro L., Ruggeri A., Posteraro B., Burioni R., Clementi M., Sanguinetti M. Comparative evaluation of the Bruker Biotyper and Vitek MS matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) mass spectrometry systems for identification of yeasts of medical importance. Journal of Clinical Microbiology, 2013, 51(7): 2453-2457 (doi: 10.1128/jcm.00841-13).
- Mather C.A., Rivera S.F., Butler-Wu S.M. Comparison of the Bruker Biotyper and Vitek MS matrix-assisted laser desorption ionization-time of flight mass spectrometry systems for identification of mycobacteria using simplified extraction protocols. Journal of Clinical Microbiology, 2014, 52(1): 130-138 (doi: 10.1128/JCM.01996-13).
- Alby K., Gilligan P., Miller M. Comparison of matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) mass spectrometry platforms for the identification of gram-negative rods from patients with cystic fibrosis. Journal of Clinical Microbiology, 2013, 51(11): 3852-3854 (doi: 10.1128/JCM.01618-13).
- Kärpänoja P., Haruju I., Rantakokko-Jalava K., Haanperä M., Sarkkinen H. Evaluation of two matrix-assisted laser desorption ionization-time of flight mass spectrometry systems for identification of viridans group streptococci. European Journal of Clinical Microbiology & Infectious Diseases, 2014, 33: 779-788 (doi: 10.1007/s10096-013-2012-8).
- Sanguinetti M., Posteraro B. Mass spectrometry applications in microbiology beyond microbe identification: progress and potential. Expert Review of Proteomics, 2016, 13(10): 965-977 (doi: 10.1080/14789450.2016.1231578).
- Ramesh Babu N.G., Simrah Fathima K.A, Nandhini V., Nandhini V. Extraction of prodigiosin from Serratia marcescens and its application as an antibacterial spray. IP International Journal of Medical Microbiology and Tropical Diseases, 2019, 5(4): 207-209 (doi: 10.18231/j.ijmmtd.2019.047).
- Batah R., Loucif L., Olaitan A.O., Boutefnouchet N., Allag H., Rolain, J.M. Outbreak of Serratia marcescens coproducing ArmA and CTX-M-15 mediated high levels of resistance to aminoglycoside and extended-spectrum beta-lactamases, Algeria. Microbial Drug Resistance, 2015, 21(4): 470-476 (doi: 10.1089/mdr.2014.0240).
- Claydon M.A., Davey S.N., Edwards-Jones V., Gordon D.B. The rapid identification of intact microorganisms using mass spectrometry. Nature Biotechnology, 1996, 14: 1584-1586 (doi: 10.1038/nbt1196-1584).
- Holland R.D., Wilkes J.G., Rafii F., Sutherland J.B., Persons C.C., Voorhees K.J., Lay J.O., Jr. Rapid identification of intact whole bacteria based on spectral patterns using matrix-assisted laser desorption/ionization with time-of-flight mass spectrometry. Rapid Communications in Mass Spectrometry, 1996, 10(10): 1227-1232 (doi: 10.1002/(SICI)1097-0231(19960731)10:10<1227::AID-RCM659>3.0.CO;2-6).
- Dieckmann R., Graeber I., Kaesler I., Szewzyk U., von Döhren H. Rapid screening and dereplication of bacterial isolates from marine sponges of the Sula Ridge by Intact-Cell-MALDI-TOF mass spectrometry (ICM-MS). Applied Microbiology and Biotechnology, 2005, 67: 539-548 (doi: 10.1007/s00253-004-1812-2).
- Ghyselinck J., van Hoorde K., Hoste B., Heylen K., De Vos P. Evaluation of MALDI-TOF MS as a tool for high-throughput dereplication. Journal of Microbiological Methods, 2011, 86(3): 327-336 (doi: 10.1016/j.mimet.2011.06.004).
- Spitaels F., Wieme A.D., Vandamme P. MALDI-TOF MS as a novel tool for dereplication and characterization of microbiota in bacterial diversity studies. In: Applications of mass spectrometry in microbiology: from strain characterization to rapid screening for antibiotic resistance /P. Demirev, T.R. Sandrin (eds.). Springer, Cham, 2016: 235-256 (doi: 10.1007/978-3-319-26070-9_9).
- Mellmann A., Cloud J., Maier T., Keckevoet U., Ramminger I., Iwen P., Dunn J., Hall G., Wilson D., LaSala P., Kostrzewa M., Harmsen D. Evaluation of matrix-assisted laser desorption ionization-time-of-flight mass spectrometry in comparison to 16S rRNA gene sequencing for species identification of nonfermenting bacteria. Journal of Clinical Microbiology, 2008, 46(6): 1946-1954 (doi: 10.1128/JCM.00157-08).
- Uhlík O., Strejček M., Junková P., Šanda M., Hroudová M., Vlček C., Mackova M., Macek T. Matrix-assisted laser desorption ionization (MALDI)-time of flight mass spectrometry- and MALDI Biotyper-based identification of cultured biphenyl-metabolizing bacteria from contaminated horseradish rhizosphere soil. Applied and Environmental Microbiology, 2011, 77(19): 6858-6866 (doi: 10.1128/AEM.05465-11).
- Koubek J., Uhlík O., Ječná K., Junková P., Vrkoslavová J., Lipov J., Kurzawova V., Macek T., Mackova M. Whole-cell MALDI-TOF: rapid screening method in environmental microbiology. International Biodeterioration & Biodegradation, 2012, 69: 82-86 (doi: 10.1016/j.ibiod.2011.12.007).
- Wieser A., Schneider L., Jung J., Schubert S. MALDI-TOF MS in microbiological diagnostics — identification of microorganisms and beyond (mini review). Applied Microbiology and Biotechnology, 2012, 93: 965-974 (doi: 10.1007/s00253-011-3783-4).
- Seng P., Abat C., Rolain J.M., Colson P., Lagier J.-C., Gouriet F., Fournier P.E., Drancourt M., La Scola B., Raoult D. Identification of rare pathogenic bacteria in a clinical microbiology laboratory: impact of MALDI-TOF mass spectrometry. Journal of Clinical Microbiology, 2013, 51(7): 2182-2194 (doi: 10.1128/JCM.00492-13).
- Fenselau C., Demirev P.A. Characterization of intact microorganisms by MALDI mass spectrometry. Mass Spectrometry Reviews, 2001, 20(4): 157-171 (doi: 10.1002/mas.10004).
- Lay J.O. Jr. MALDI-TOF mass spectrometry of bacteria. Mass Spectrometry Reviews, 2001, 20(4): 172-194 (doi: 10.1002/mas.10003).
- Newman M.M., Kloepper L.N., Duncan M., McInroy J.A., Kloepper J.W. Variation in bat guano bacterial community composition with depth. Frontiers in Microbiology, 2018, 9: 914 (doi: 10.3389/fmicb.2018.00914).
- Veikkolainen V., Vesterinen E.J., Lilley T.M., Pulliainen A.T. Bats as reservoir hosts of human bacterial pathogen, Bartonella mayotimonensis. Emerging Infectious Diseases, 2014, 20(6): 960-967 (doi: 10.3201/eid2006.130956).
- Wolkers-Rooijackers J., Rebmann K., Bosch T., Hazeleger W. Fecal bacterial communities in insectivorous bats from the Netherlands and their role as a possible vector for foodborne diseases. Acta Chiropterologica, 2019, 20: 475 (doi: 10.3161/15081109acc2018.20.2.017).
- Banskar S., Bhute S.S., Suryavanshi M.V., Punekar S., Shouche Y.S. Microbiome analysis reveals the abundance of bacterial pathogens in Rousettus leschenaultii guano. Scientific Reports, 2016, 6: 36948 (doi: 10.1038/srep36948).
- Selvin J., Lanong S., Syiem D., De Mandal S., Kayang H., Kumar N.S., Kiran G.S. Culture-dependent and metagenomic analysis of lesser horseshoe bats’ gut microbiome revealing unique bacterial diversity and signatures of potential human pathogens. Microbial Pathogenesis, 2019, 137: 103675 (doi: 10.1016/j.micpath.2019.103675).
- AL-Ghanem M.M. Serratia A novel source of secondary metabolites. Advances in Biotechnology & Microbiology, 2018, 11(3): 555814 (doi: 10.19080/AIBM.2018.11.555814 ).
- Bhadra B., Ro P., Chakraborty R. Serratia ureilytica sp. nov., a novel urea-utilizing species. International Journal of Systematic and Evolutionary Microbiology, 2005, 55(5): 2155-2158 (doi: 10.1099/ijs.0.63674-0).
- Frankowski J., Lorito M., Scala F., Schmid R., Berg G., Bahl H. Purification and properties of two chitinolytic enzymes of Serratia plymuthica HRO-C48. Archives of Microbiology, 2001, 176(6): 421-426 (doi: 10.1007/s002030100347).
- Czajkowski R., Wolf J.M. Draft genome sequence of the biocontrol strain Serratia plymuthica A30, isolated from rotting potato tuber tissue. Journal of Bacteriology, 2012, 194: 6999-7000 (doi: 10.1128/jb.01699-12).
- Petersen L.M., Tisa L.S. Friend or foe? A review of the mechanisms that drive Serratia towards diverse lifestyles. Canadian Journal of Microbiology, 2013, 59(9): 627-640 (doi: 10.1139/cjm-2013-0343).
- Kimyon Ö., Das T., Ibugo A.I., Kutty S.K., Ho K.K., Tebben J., Kumar N., Manefield M. Serratia secondary metabolite prodigiosin inhibits Pseudomonas aeruginosa biofilm development by producing reactive oxygen species that damage biological molecules. Frontiers in Microbiology, 2016, 7: 972 (doi: 10.3389/fmicb.2016.00972).
- Stankovic N., Senerovic L., Ilic-Tomic T., Vasiljevic B., Nikodinovic-Runic J. Properties and applications of undecylprodigiosin and other bacterial prodigiosins. Applied Microbiology and Biotechnology, 2014, 98(9): 3841-3858 (doi: 10.1007/s00253-014-5590-1).