doi: 10.15389/agrobiology.2022.3.425rus
УДК 631.427.3.631.427.4:577.3
БИОЭЛЕКТРОХИМИЧЕСКИЕ СИСТЕМЫ НА ОСНОВЕ ЭЛЕКТРОАКТИВНОСТИ РАСТЕНИЙ И МИКРООРГАНИЗМОВ В КОРНЕОБИТАЕМОЙ СРЕДЕ (обзор)
Т.Э. КУЛЕШОВА1, А.С. ГАЛУШКО1, Г.Г. ПАНОВА1 ✉, Е.Н. ВОЛКОВА1, W. APOLLON2, C. SHUANG3, S. SEVDA4
Биоэлектрохимические системы (БЭС) на основе электроактивных процессов в корнеобитаемой среде растений и сопутствующих микроорганизмов — новая перспективная экологически чистая технология получения возобновляемой энергии. Хотя возможность практического использования биоэнергетических ресурсов уже показана во многих исследованиях, природа электрогенеза и влияние на нее внешних факторов до конца не изучены. Возникновение разности потенциалов в живых системах обусловлено комплексом физико-химических процессов, обеспечивающих поддержание неравномерного распределения ионов на уровне клеток, тканей и организма (N. Higinbotham, 1970). В процессе развития растений вдоль всего организма возникает градиент электрического потенциалов, обусловленный диффузией ионов, концентрационными эффектами и различиями в интенсивности биохимических процессов (T.A. Tattar с соавт., 1976). Наряду с этим микроорганизмы ризосферы способны окислять органические вещества в составе экссудатов корней (L. De Schamphelaire с соавт., 2010). Образовавшиеся в процессе окислительно-восстановительных реакций ионы и электроны диффундируют по корнеобитаемой среде, приводя к разделению зарядов (B.E. Logan, 2008), в результате устанавливается градиент электропотенциалов, связанный с различиями в концентрациях заряженных веществ. Комплекс этих процессов преобразования энергии в виде химических связей органических веществ в электрическую энергию лежит в основе устройства растительно-микробного топливного элемента (РМТЭ). Наиболее распространенная конфигурация устройства РМТЭ состоит из анодной и катодной камер, ионоселективной мембраны (D.P. Strik с соавт., 2008), существуют также различные модификации в виде плоской пластины (M. Helder с соавт., 2013), трубки (R.A. Timmers с соавт., 2013), направленные на увеличение выходных электрических характеристик. Одной из важнейших составляющих БЭС выступают электродные системы, наиболее часто используют углеродные материалы, которые обладают большой удельной площадью поверхности, высокой электропроводностью, коррозионностойкостью. Производительность БЭС зависит от состава корнеобитаемой среды, наличия потенциалобразующих ионов, параметров световой среды, эффективности фотосинтеза. Один из вариантов эксплуатации РМТЭ — их комбинирование с значимыми производственными процессами, в частности внедрение в аграрное производство. Возможность применения БЭС показана на ряде культурных и технических растений с получением следующего выхода энергии малой мощности: при выращивании риса — 140 (N. Ueoka с соавт., 2016), салата — 54 (Т.Э. Кулешова с соавт., 2021), манника — 80 (R.A. Timmers с соавт., 2012), тростника — 42 (J. Villasenor с соавт., 2013), рогоза — 93 (Y.L. Oon с соавт., 2016), спартины — 679 мВт/м2 (K. Wetser с соавт., 2015). Использование БЭС перспективно для обеспечения электропитанием датчиков окружающей среды (A. Schievanoс соавт., 2017), источников света (W. Apollon с соавт., 2020), беспроводных сенсорных сетей (E. Osorio-De-La-Rosa с соавт., 2021), интернета вещей (JayaramanP.P. с соавт., 2016), систем фитомониторинга в естественных условиях и защищенном грунте, удаленных районах, частичное энергоснабжение устройств поддержки жизнедеятельности растений в искусственных агроэкосистемах (Т.Э. Кулешова с соавт., 2021), очистки сточных вод (L. Kook с соавт., 2016).
Ключевые слова: «зеленая» энергия, растительно-микробный топливный элемент, биоэлектрогенез, электроактивные бактерии.
В настоящее время основную долю энергетического рынка занимают ископаемые виды топлива — уголь, нефть и природный газ, потребление которых приводит к загрязнению окружающей среды и изменению климата. В этой связи актуально использование экологически чистых возобновляемых природных энергоресурсов. Солнечная энергия, ветер, геотермальная теплота, гидротермальная энергия, биотопливо интенсивно применяются для генерации электроэнергии. Однако у них также есть недостатки, заключающиеся в высокой стоимости установок, зависимости от погодных условий и времени суток, трансформации ландшафта, географической локализации. На фоне этих ограничений потенциал развития имеют биоэлектрохимические системы (БЭС) на основе электроактивных процессов, сопутствующих жизнедеятельности растений и окружающих их ризосферных микроорганизмов.
Использование биоэнергетических ресурсов для становления новой области «зеленой» энергии — сложная и не до конца изученная задача, требующая интеграции широкого круга знаний в областях физики, электрохимии и биологии.
Цель настоящего обзора — анализ существующих конструкций биоэлектрохимических систем, описание электрогенных и потенциалобразующих реакций, протекающих в БЭС, и влияния на них отдельных факторов среды обитания, а также рассмотрение перспектив использования биоэнергетических устройств.
Электрические процессы в корнеобитаемой среде. Электрогенез растений. История исследований электрофизиологических свойств растений насчитывает более сотни лет, однако механизм биоэлектрогенеза, то есть способности к перемещению заряда и генерации электроэнергии (1), до сих пор остается предметом дискуссий. Принято считать, что возникновение разности потенциалов в живых системах обусловлено в первую очередь комплексом физико-химических процессов, обеспечивающих поддержание неравномерного распределения ионов на уровне клеток, тканей и организма.
Основной электрической характеристикой клетки служит мембранный потенциал, возникающей в первую очередь в результате диффузии и активного процесса переноса ионов между внеклеточной средой и внутриклеточными компартментами (2). Активному транспорту наиболее подвержены ионы K+, Na+, Ca2+, Mg2+, NO3-, Cl-, H2PO4-, SO4-. Многие другие органические вещества, подвижные внутри клеток и тканей, также несут заряды, например органические кислоты, аминокислоты, аденозин фосфаты и т.д. (3). Разности потенциалов между участками тканей и органов растений, генерируемые в результате электрогенного активного транспорта, определяются физиологическим состоянием и разделяются на потенциалы покоя, действия, повреждения и течения (4). Градиенты биоэлектрического потенциала (БЭП) возникают в результате протекания метаболических реакций вдоль всего растительного организма (3).
Таким образом, диффузия ионов, концентрационные эффекты и работа ионных насосов приводят к возникновению электрического тока в растительных организмах в процессе их жизнедеятельности (5). Электрогенные свойства наиболее интенсивно проявляются в системе корнеобитаемая среда — растения, что связано с поглощением и транспортом ионов в процессе минерального питания (6). Например, потенциал покоя клеток высших растений в среднем варьирует в пределах 50-120 мВ (7), тогда как биоэлектрический потенциал в прикорневой зоне может достигать 700 мВ (8).
Электроактивные бактерии. Наряду с диффузией ионов, сопутствующей жизнедеятельности растений, разделение и перемещение зарядов в корнеобитаемой среде может осуществляться электроактивными бактериями. В процессе развития микроорганизмы ризосферы способны окислять выделяемые корнями органические вещества, синтезируя при этом углекислый газ, протоны H+ и электроны e- (9). Преобразование энергии химических связей органических веществ в электрическую энергию лежит в основе биотехнологического устройства — микробного топливного элемента. В нем генерируемые электроны под действием разности окислительно-восстановительных потенциалов по внешней цепи перемещаются к противоположному электроду, где соединяются с протонами, мигрировавшим, например, через ионоселективную мембрану, и кислородом, образуя воду (10).
Транспорт электронов от электрохимически активных бактерий к поверхности электрода может быть осуществлен и прямым путем при непосредственном контакте с электродом, и с помощью электропроводящих отростков (пилей) или медиаторов (11). В частности, перенос электронов на анод от бактерий видов Shewanella и Geobacter осуществляется как прямо, так и с использованием пилей (12), а Pseudomonas секретируют медиаторы (флавины) (13).
В настоящее время известно множество видов бактерий (14), применимых в микробных топливных элементах. Методом флуоресцентной гибридизации in situ на корнях растений были идентифицированы бактерии, потенциально способные к осуществлению электрохимических реакций в корнеобитаемой среде, — Geobacter serreducens, Geobacter Metallireducens, Geobacter grbiciae, Geobacter Hydrogenophilus, Ruminococcus bromii, Clostridium sporosphaeroides и Clostridium leptum (15). Определено, что в качестве донора электронов бактерии Shewanellaputrefaciens могут использовать лактат, пируват и формиат (16); Clostridium butyricum и Clostridium beijerinckii — глюкозу, крахмал, лактат (17); Rhodopseudomonaspalustris — ацетат, лактат, валерат, фумарат, этанол, глицерин (18); Geobacter serreducens (19) ,Geobacter sulfurreducens (20) и Geobacter metallireducens (21) — ацетат; Rhodoferax ferrireducens (22), Alcaligenes faecalis, Enterococcus gallinarum и Pseudomonas aeruginosa (23) — глюкозу; Enterobactercloacae — целлюлозу (24). Большинство из перечисленных соединений присутствуют в корнеобитаемой среде как продукты жизнедеятельности биоты и служат энергоресурсом для электрохимически активных бактерий.
Градиент электрического потенциала в корнеобитаемой среде. Химические реакции в корнеобитаемой среде, протекающие в результате жизнедеятельности растений и сопутствующих микроорганизмов, также служат источником электронов и ионов (9, 25, 26). Ионы и электроны, образовавшиеся в процессе окислительно-восстановительных реакций, диффундируют по корнеобитаемой среде, приводя к разделению зарядов. В результате в почве или почвозаменителе устанавливается градиент электропотенциалов, связанный с различиями в концентрациях заряженных веществ (27, 28).
Формирование различной плотности подвижных зарядов, обусловленной диффузией и адсорбцией их носителей (29), — неотъемлемая часть обмена веществ, сопутствующего функционированию и развитию растений и микроорганизмов. В эксперименте по сравнению градиента электрических потенциалов в почве и при выращивании на ней фитотест-объекта — ярового ячменя (Hordeum vulgare L.) сорта Ленинградский (30), было показано, что изменение разности потенциалов по профилю почвы связано со стадией развития растений и степенью прорастания корневой системы (табл. 1).
Создание градиента электрического потенциала наблюдается и при развитии корневой системы в почве или почвозаменителе, и в самой почвенной структуре без растений, что свидетельствует о наличии процессов ионного транспорта в ней, например за счет диффузии вместе с водным потоком. Растения в сообществе с микроорганизмами ризопланы и ризосферы, по-видимому, запускают дополнительные реакции, поглощая и выделяя различные органические и минеральные соединения, и увеличивают интенсивность процессов в почве.
Биоэлектрические измерения и особенности конструкции. Растительно-микробный топливный элемент. На основе способности микроорганизмов выступать в качестве катализаторов окислительно-восстановительных реакций, включающих внеклеточный перенос электронов от микробов к электроду (31), была разработана биоэлектрохимическая система, названная растительно-микробным топливным элементом (РМТЭ) (32).
РМТЭ представляет собой модификацию микробного топливного элемента и включает, помимо катода и анода с размещенными на нем микроорганизмами, живые растения, которые вырабатывают ризодепозиты — субстрат для электроактивных бактерий (рис. 1). Наиболее распространенная конфигурация устройства РМТЭ состоит из трех частей: анодной камеры, ионоселективной мембраны и катодной камеры. В анодной камере микроорганизмы как катализаторы процесса окисления преоб-разуют органические вещества, выделяемые корнями, согласно реакции C6H12O6 + 6H2O → 6CO2 + 24e- + 24H+. Попадая на анод, электроны перемещаются по внешней цепи к катоду. Протоны мигрируют через ионоселективную мембрану в катодную камеру, где с участием электронов происходит восстановление кислорода с образованием молекул воды (33).
В настоящее время направление РМТЭ активно развивается, создаются различные модификации устройства, направленные на увеличение эффективности и электрических характеристик. Например, для уменьшения расстояния переноса протонов между электродами разработана конфигурация плоской пластины: в ней между близко расположенными анодной и катодной камерами размещается катионообменная мембрана. Мощность такого РМТЭ составила 240 мВт/м2 при длительной работе в течение 151 сут (34). Для упрощения интеграции в естественную среду была разработана модель, в которой анод и катод объединены в единый блок в виде трубки. Максимальная выходная мощность для этого варианта достигала 72 мВт/м2 (35).
Перспективный вариант использования РМТЭ — их комбинирование со значимыми производственными процессами. Например, возможно внедрение таких топливных элементов в системы очистки сточных вод (36) и в аграрное производство.
Биосовместимые электродные системы. Одна из важнейших характеристик эффективности работы БЭС — электродные материалы. Кроме высоких электрических показателей, они должны обладать свойствами химической стабильности и биосовместимости. Наиболее часто в качестве электродов (анода и катода) используют материалы на основе углерода (табл. 2): графитовые войлок, ткань, гранулы, стержень, углеродную бумагу, сетчатый стекловидный углерод (37).
В РМТЭ на основе папируса (Cyperus papyrus nanus L.) и красного корня (Wachendorfia thyrsiflora Burm.) были использованы графитовые гранулы в качестве анода и углеродный лист в виде катода. Обеспеченная в таком варианте большая площадь соприкосновения электродов с корнями растений и доступность кислорода для катодных реакций образования воды привели к получению высоких показателей выходной мощности до 1036±59 мВт/м3 (38). В работах с использованием манника (Glyceria maxima Hartm.) исследовано влияние материала электрода на внутреннее сопротивление системы, которое в основном складывается из сопротивлений анода и мембраны (40). Поэтому подходящий биосовместимый анодный электрод играет очень важную роль в снижении энергетических потерь. Максимальная выработка энергии в предложенном варианта РМТЭ в виде трубки составила 10 мВт/м2 при использовании графитового войлока в качестве анода и 12 мВт/м2 с применением графитовых гранул (35). Перспективно применение биокатода, на котором восстановление кислорода катализируется микроорганизмами. С его использованием выработка электроэнергии была увеличена до 679 мВт/м2 в РМТЭ на основе кордграсса (Spartina anglica Hubbard) (39). В условиях реального применения, например развивающегося направления выращивания растений на крышах, максимально достигнутая мощность РМТЭ составила 88 мВт/м2 в сравнении с 440 мВт/м2, получаемыми на лабораторной установке (41), что, вероятнее всего, связано с изменениями свойства субстрата в результате погодных условий. Следовательно, для применения в естественных условиях электродные системы еще необходимо модифицировать, снижая их площадь и сопротивление и увеличивая толерантность к внешним факторам.
Электрические параметры. Измеряемой характеристикой, отражающей биоэлектрическую активность корневой системы и сопутствующих микроорганизмов и протекание метаболических процессов в корнеобитаемой среде, служит электрическое напряжение U (В), определяемое по закону Ома: U = ε - I·r, где ε характеризует электродвижущую силу (ЭДС) БЭС, то есть работу сторонних сил по перемещению заряда, а произведение силы тока I на внутреннее сопротивление системы r определяет падение напряжения внутри системы. Из этого следует, что производительность БЭС и ее выходные характеристики сильно зависят от способности питательной среды пропускать электрический ток. Одним из наиболее распространенных способов снижения влияния внутреннего сопротивления служит уменьшение расстояния между электродами, то есть промежутка на который необходимого перенести заряд (42).
Для эффективного исследования электрических явлений в живом организме и окружающей их среде способ отведения электропотенциалов должен удовлетворять следующим условиям: 1) обеспечивать надежный электрический контакт электрода с исследуемым объектом, 2) исключать возможность возникновения поляризационных потенциалов, 3) учитывать электрокинетические явления, возникающие в корнеобитаемой среде, 4) исключать возможность повреждения биообъекта (43). Метод измерения генерируемой в системе корнеобитаемая среда — растения разности потенциалов, удовлетворяющий этим условиям, был предложен Т.Э. Кулешовой с соавт. (44). Он основан на не повреждающем неинвазивном способе обеспечения поверхностного электрического контакта между корневой системой и электродами. Скорость получения электрической энергии с помощью БЭС на основе растений и микроорганизмов характеризуется единицами электрической мощности P = I·U и, как правило, нормируется на площадь, занимаемую растениями. В таблице 3 представлены некоторые полученные значения мощностей для РМТЭ с различными конфигурациями, растительными объектами и питательными средами. Несмотря на то, что в настоящее время БЭС представляют маломощные устройства, они обладают рядом уникальных свойств, позволяющих обеспечивать экологически чистую автономную энергию воспроизводимым путем, что имеет большие перспективы применения.
Роль факторов окружающей среды. Параметры окружающей среды играют существенную роль в жизнедеятельности растений и сопутствующей им микрофлоры, включая процессы биоэлектрогенеза. Наиболее значимые факторы, влияющие на функционирование БЭС, — состав и условия корнеобитаемой и световой сред.
Влияние состава корнеобитаемой среды. В БЭС на основе электроактивности растений и ризосферных бактерий используются разнообразные корнеобитаемые среды, включая: почвы сельскохозяйственных, лесных и водно-болотных угодий, почвозаменители, а также песок, глина, компост, ил, солончак и др. (47, 62, 63). При этом состояние и концентрации компонентов питательной среды растений (по-видимому, так же, как и свойства электролита в гальваническом элементе) имеют решающую роль в выходных электрических характеристиках БЭС.
Основным «топливом», окисляемым электрохимически активными бактериями, служат ризодепозиты: из всего фотосинтезированного углерода около 20-40 % попадает в корнеобитаемую среду в различных формах в виде корневых экссудатов, метаболитов и отмерших частей растений (64). Выделяемые корнями органические соединения в основном включают органические кислоты, фенолы, сахара и аминокислоты, и высокомолекулярные соединения, такие как полисахариды и белки (65). Их состав зависит от вида растений, способа связывания углерода, интенсивности роста, возраста растений, а также условий окружающей среды (66).
Наряду с составом корнеобитаемой среды потенциалобразующую роль при разделении зарядов играет подвижность ионов. Наибольшей скоростью движения в водных растворах обладают катионы H+, H3O+ (36,2 м2/В·с), NH4+, K+ (7,6 м2/В·с), Fe3+ (7 м2/В·с) и анионы OH- (20,5 м2/В·с), Cl- (7,9 м2/В·с), NO3- (7,4 м2/В·с) (67). Кроме того, величина разности потенциалов зависит от влажности почвы, включая как изменение сопротивления среды, так и процессы поглощения и транспорта воды, связанные с жизнедеятельности растений. Например, Т.Э. Кулешовой с соавт. (68) показана зависимость электрического напряжения, создаваемого в прикорневой зоне, от водного режима, включая вододефицитные условия.
Зависимость от световой среды. Известно, что свет играет важную роль в формировании биоэлектрических потенциалов. Например, при включении света наблюдается резкое падение БЭП листовой поверхности, а затем быстрый скачок (69). Если одну часть листа осветить, а вторую затенить, то разность потенциалов между ними будет изменяться от 50 до 100 мВ (70). Такая вариация метаболических потенциалов связана в первую очередь с различиями в интенсивности биохимических процессов в разных частях растения.
Изменение разности потенциалов в ответ на световое воздействие отмечается и при электроактивных реакци-ях в корнеобитаемой сре-де. Показано, что при смене темновой стадии на световую напряжение в прикорневой зоне плавно увеличивается на 10-15 % и затем равномерно спадает (рис. 2), эта полусуточная динамика может быть описана полиномом второй степени. Параболические изменения напряжения во время световой стадии фотосинтеза и стационарная генерация при его темновом режиме, вероятнее всего, связаны с интенсивностью транспорта воды, минеральных и органических веществ в зависимости от формируемых световых условий. Также известно, что электрическое сопротивление листовой поверхности зависит как от температуры и влагосодержания ткани, так и подвижности и концентрации ионов в тканевой среде: сопротивление листа увеличивается при подвядании и снижается при поливе до первоначальной величины (71).
Таким образом, факторы внешней среды играют значительную роль в формировании и протекании биоэлектрических процессов. Показана (72) корреляция динамики пропускания света листовой пластиной и разности потенциалов в системе корнеобитаемая среда-растений, которая свидетельствует о возможной конверсии световой энергии листьями растений в электрический ток в ризосфере.
Применение БЭС. БЭС, комбинированная с производством растительной продукции. РМТЭ — возобновляемый источник энергии, который может одновременно производить биоэлектричество и биомассу экологически чистым, устойчивым и эффективным образом (28). Использование гибридной технологии, позволяющей производить растительную продукцию и осуществлять генерацию электроэнергии за счет активации окислительных процессов в ризосфере, — инновационное направление с перспективой применения в области автономного автоматизированного агропроизводства. Возможность получения энергии малой мощности с помощью БЭС при выращивании растений уже была показана на некоторых культурных и технических растениях, включая рис (73), салат (47), манник (15), камыш (74), рогоз (75), используемые в качестве пищи, топлива, строительных материалов, корма для животных.
Универсальное устройство БЭС, предложенное Т.Э. Кулешовой с соавт. (76) и пригодное для выращивания овощных культур (зеленные, томаты, огурцы), основано на технологии тонкослойной панопоники (77). В таком РМТЭ электродные системы размещаются в емкости для культивирования перпендикулярно росту корневой системы, тем самым обеспечивается не повреждающий растения поверхностный контакт корней с электропроводящим материалом (44). При этом предполагается, что формирование градиента электропотенциалов в БЭС — это следствие перемещения ионов вдоль корневой системы и концентрационных эффектов, а возникновение ЭДС между электродами обеспечивается за счет жизнедеятельности растений и электроактивности микробного сообщества, окружающего корневую систему.
Технология РМТЭ позволяет производить «зеленую» энергию практически везде, где растут растения, и применима как в естественной среде, так и для выращивания сельскохозяйственных культур в открытом грунте и теплицах, в фитотехнических комплексах и регулируемых агроэкосистемах, что особенно важно для районов, географически изолированных от единой энергосистемы. Агротехнологический энергетический комплекс на базе БЭС способен обеспечить получение не только экологически чистой энергии, но и высококачественной растительной продукции.
Биобатареи в окружающей среде. Биобатареи — это экологически чистые комплексные биоэлектрохимические системы, которые из или с помощью биоресурсов преобразуют химическую энергия в электричество. В отличие от обычных батарей, которые вызывают загрязнение окружающей среды (78), биобатареи считаются устойчивым и возобновляемым источником энергии. Однако оценка жизненного цикла (life cycle assessment, LCA) этих систем до и после их внедрения все еще остается актуальной задачей (79) и зависит от типов материала, используемых при изготовлении конструкции.
Биобатареи включают электродные системы из анода и катода и могут использовать различные биохимические источники энергии (80-84).
Биобатареи уже были протестированы на разных видах растений и в варьируемых условиях окружающей среды (85). В частности, были использованы растения семейства кактусовые (Opuntia Miller), осуществляющие CAM-фотосинтез и применимые в засушливых условиях (86).
Биобатарея была сконструирована с использованием вертикально интегрируемой керамической трубки, содержащей графитовой войлок в качестве материала анода и фольгу из цинка в качестве катода. Максимально достигнутая плотность мощности с использованием опунции (Opuntia albicarpa Scheinvar) составила 103,6 мВт/м2 в условиях длительной эксплуатации. Добавление в биобатарею на основе опунции (Opuntia joconostle Weber ex Diguet) нитрата аммония (150 мг·л-1·нед-1) привело к увеличению выхода энергии с 40 до 500 мВт/м3 (87). Разработанные биобатерии были эффективно применены для электропитания светодиодов и цифровых часов, обеспечивая их автономную работу в течение недели (85). Использование в БЭС растений с CAM-фотосинтезом перспективно для районов с ограниченными ресурсами и на полузасушливых территориях. Однако для этого необходимо проведение крупномасштабных исследований.
Фитомониторинг и электропитание датчиков. БЭС могут выполнять двойную функцию, выступая в качестве биосенсора для фитомониторинга и обеспечивая электропитание датчиков параметров окружающей среды. В работе D. Brunelli с соавт. (88) РМТЭ был использован для отслеживания физиологического состояния растений и мониторинга интенсивности света и влажности почвы. Энергия от РМТЭ накапливалась на суперконденсаторе и затем использовалась для отправки сигнала с датчиков с интервалом 15 мин.
Разрабатываемые беспроводные сенсорные сети (wireless sensor network, WSN) и Интернет вещей (internet of things, IoT) имеют приоритетное значение для задач интеллектуального земледелия, непрерывного мониторинга состояния и потребностей растений (89), в особенности для применения в удаленных от энергосетей районах. РМТЭ могут обеспечить экологически безопасный вариант для электропитания этих систем. В настоящее время проблема маломощного и прерывистого получения энергии с помощью БЭС решается интегрированием в нее суперконденсаторов (90). В работе E. Osorio-De-La-Rosa с соавт. (91) был продемонстрирован запуск датчика на основе IoT. Установка была способна генерировать 3,5 мВт/см2 с выходным напряжением 0,5 В, что достаточно для безбатарейной работы узла датчиков для сбора данных о температуре и их облачного хранения.
Таким образом, БЭС может быть применена для создания маломощных необслуживаемых возобновляемых источников электроэнергии, способных частично поддерживать жизнедеятельность растений посредством подачи питания на источники света, насосы, датчики параметров состояния растений и окружающей среды. Также ее можно использовать в научных исследованиях и в растениеводстве в качестве биосенсора настройки технологий выращивания и осуществления фитомониторинга.
Очистка сточных вод на основе БЭС. Внедрение БЭС стремительно развивается и в области очистки сточных вод. По сравнению с традиционными технологиями, БЭС более рентабельны и устойчивы, так как имеют преимущество, связанное с возобновляемостью биоэнергетических ресурсов (92). Изучаются различные возможности их применения, в частности перспективным считается использование электрохимически активных микроорганизмов для удаления органических веществ и тяжелых металлов. БЭС могут продуктивно работать для окисления органических веществ в анодной камере, особенно в отношении городских и промышленных сточных вод с высоким химическим потреблением кислорода, например пивоваренные, пищевые и текстильные (93-96). Органические отходы также могут выступать в качестве донора электронов для микроорганизмов при восстановлении тяжелых металлов. В работе Y.V. Nancharaiah с соавт. (97) приведены сведения о восстановлении в катодной камере БЭС ионов Ag (I), Au(III), Co(III), Cr(VI), Cu(II), Hg(II), Se(IV) или V(V). Эффективность восстановления Cr (VI) в работе H. Yu с соавт. (98) достигала 99,93 %.
В результате протекания микробных процессов в БЭС также возможно удаление азота (99). Например, N. Yang с соавт. (100) показали высокую эффективность удаления NH4+-N (99 %) и общего азота (TN, 99 %) в БЭС с восходящим потоком, в которых микробный метаболизм был усилен для осуществления одновременной нитрификации, денитрификации и других биоэлектрохимических реакций.
Итак, биоэлектрохимические системы на основе электроактивных процессов в корнеобитаемой среде растений и сопутствующих микроорганизмов — новая перспективная экологически чистая технология получения возобновляемой энергии. Производительность БЭС зависит от целого ряда показателей, включая генетически обусловленные физиологические особенности растений и их состояние на протяжении развития, состав и активность микробного сообщества, параметры корнеобитаемой среды, факторы окружающей среды, конструкцию биореактора, тип и расположение электродных систем. Несмотря на небольшую выходную мощность, растительно-микробные энергетические устройства имеют свою нишу применение и в настоящем, и в будущем — обеспечение электропитанием датчиков окружающей среды, источников света, беспроводных сенсорных сетей, интернета вещей, систем фитомониторинга в естественных условиях и защищенном грунте, удаленных районах, частичное энергоснабжение устройств поддержки жизнедеятельности растений в искусственных агроэкосистемах. Получение «зеленой» энергии может сопровождаться производством растительной продукции и очисткой сточных вод. Дальнейшие перспективы развития лежат в создании многофункциональных электрических схем, учитывающих свойства корнеобитаемой среды и растений, для повышения эффективности и увеличения количества генерируемой электроэнергии.
1. Разность электропотенциалов в почве по ее профилю на протяжении роста растений ярового ячменя (Hordeum vulgare L.) сорта Ленинградский |
|||||||||
Номер электрода |
Расстояние от поверхности почвы до электрода, мм |
Вегетационный период |
|||||||
0-5-е сут |
5-13-е сут |
13-18-е сут |
18-51-е сут |
||||||
МПД, мВ |
ДРЗ, мВ |
МПД, мВ |
ДРЗ, мВ |
МПД, мВ |
ДРЗ, мВ |
МПД, мВ |
ДРЗ, мВ |
||
1-й |
30 |
112 |
18 |
151 |
18 |
107 |
14 |
190 |
23 |
2-й |
80 |
151 |
27 |
176 |
21 |
132 |
16 |
151 |
30 |
3-й |
130 |
103 |
18 |
73 |
4 |
34 |
5 |
298 |
63 |
4-й |
180 |
73 |
14 |
103 |
15 |
98 |
13 |
337 |
81 |
5-й |
230 |
98 |
22 |
132 |
13 |
103 |
10 |
132 |
23 |
6-й |
280 |
73 |
13 |
73 |
14 |
54 |
11 |
63 |
9 |
Примечание. МПД — максимальное падение напряжения относительно электрода на дне сосуда, ДРЗ — диапазон рассеивания значений (по материалам 30). |
|||||||||
2. Электроды, используемые в растительно-микробных топливных элементах (РМТЭ) |
||||||
Конструкция РМТЭ |
Материал |
Материал |
Вид растения |
Субстрат |
Выходная |
Ссылка |
Горшок (5 л) |
Графитовые гранулы |
Углеродный лист с платиновым покрытием (0,4 мг/см2) |
Папирус (Cyperuspapyrusnanus L.), красный корень (Wachendorfia thyrsiflora Burm.) |
Почвенная смесь, ил |
1036±59 мВт/м3 (Wachendorfia thyrsiflora Burm.), 510±92 мВт/м3 (Cyperus papyrus L.) |
(38) |
Стеклянный цилиндр |
Графитовые гранулы |
Графитовый войлок |
Манник большой (Glyceria maxima Hartm.) |
Раствор Хогланда с буферным раствором фосфата калия (8 ммоль/л) |
0,39 Вт/м2 |
(15) |
В виде трубки |
Графитовый войлок и графитовые гранулы |
Графитовый войлок |
Манник большой (Glyceria maxima Hartm.) |
Раствор Хогланда, богатый аммонием |
10 мВт/м2 для войлока, |
(35) |
Плоская пористая пластина |
Три слоя графитового войлока |
Один слой графитового войлока |
Кордграсс (Spartina anglica Hubbard) |
Безнитратная, богатая аммонием среда для роста растений |
679 мВт/м2 |
(39) |
Условия для выращивания на крыше |
Графитовые гранулы |
Графитовый войлок |
Кордграсс (Spartina anglica Hubbard) |
Почвенная смесь и дождевая вода |
88 мВт/м2 |
(34) |
3. Вырабатываемые электрические мощности растительно-микробных топливных элементов |
|||
Растительный объект |
Корнеобитаемая среда |
Плотность мощности, мВт/м2 |
Ссылка |
Хлорофитум хохлатый (Chlorophytum comosum Thunberg) |
Почва |
18 |
(45) |
Тростник обыкновенный (Phragmites australis Cavanilles) |
Глюкоза + ацетат натрия |
43 |
(46) |
Салат (Lactuca sativa L.) |
Питательный раствор |
54 |
(47) |
Ипомея водяная (Ipomoea aquatic L.) |
Анаэробный ил и питательные |
55 |
(48) |
Горчица сарептская (Brassica juncea L.) |
Компостная почвенная смесь |
70 |
(49) |
Манник крупный (Glyceria maxima Hartm.) |
Гранулы графита |
80 |
(50) |
Пажитник сенной (Trigonella foenumgraecum L.) |
Почва |
80 |
(49) |
Бескильница расставленная (Puccinellia distans Jacq.) |
Почвенные смеси |
84 |
(51) |
Рогоз (Typha latifolia L.) |
Синтетические сточные воды |
93 |
(52) |
Солончаковая трава Sporobolus arabicus Boiss. и Cynodon dactylon L. |
Почва |
120 |
(53) |
Рис (Oryza sativaL.) |
Рисовые поля |
140 |
(54) |
Перистощетинник щетинистый (Pennisétum setaceum Forsskal) |
Красная почва |
163 |
(55) |
Элодея (Elodea Michaux) |
Смешанный культуральный ил |
185 |
(56) |
Канна Штутгарт (Canna stuttgart L.) |
Морской осадок |
223 |
(49) |
Водяной гиацинт (Eichhornia crassipes Mart.) |
Осадки |
225 |
(57) |
Ветивер (Chrysopogon zizanioides L.) |
Садовая земля |
242 |
(58) |
Канна индийская (Canna indica L.) |
Сброженный навоз |
320 |
(59) |
Ряска (Lemna L.) |
Источники углерода |
380 |
(60) |
Спартина английская (Sporobolus anglicus Hubbard) |
Почва |
679 |
(61) |
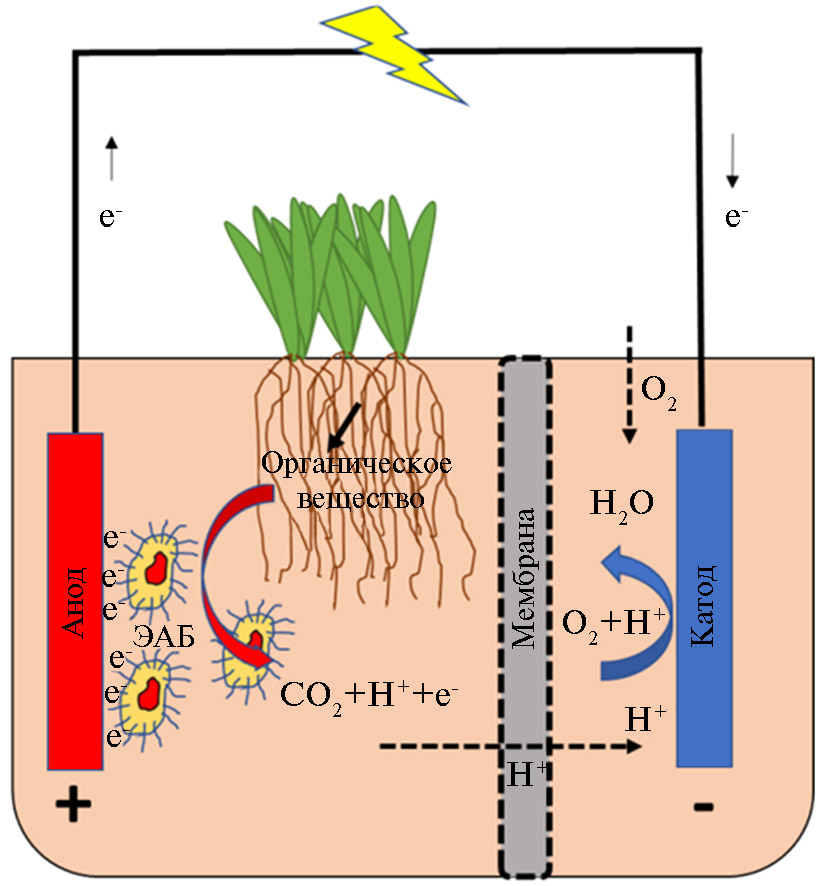
Рис. 1. Схематичное представление генерации электроэнергии в растительно-микробном топливном элементе: электроактивные бактерии (ЭАБ) окисляют органическое вещество, выделяемое корнями растений, в результате чего образуются углекислый газ, электроны, попадающие на анод, и протоны, мигрирующие через ионообменную мембрану к катоду по градиенту потенциалов, где с участием поступающих по внешней цепи электронов и кислорода образуются молекулы воды.
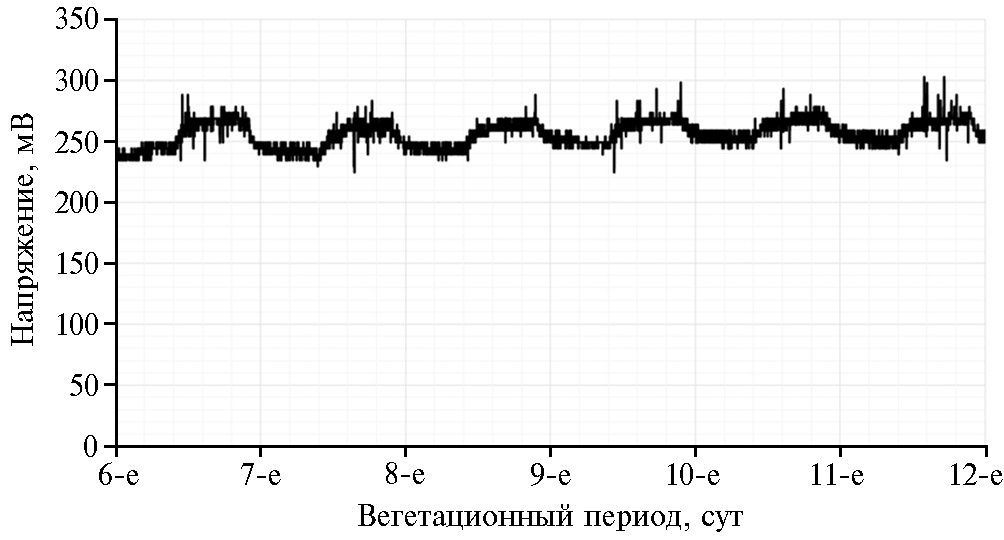
Рис. 2. Изменение электрического напряжения в системе корнеобитаемая среда — растения хлорофитума хохлатого (Chloro-phytum comosum Thunberg) при смене 12-часовых светового и темнового периодов.
ЛИТЕРАТУРА
- Дедю И.И. Экологический энциклопедический словарь. Кишинев, 1990.
- Brenner E.D., Stahlberg R., Mancuso S., Vivanco J., Baluška F., Van Volkenburgh E. Plant neurobiology: an integrated view of plant signaling. Trends in Plant Science, 2006, 11(8): 413-419 (doi: 10.1016/j.tplants.2006.06.009).
- Higinbotham N. Movement of ions and electrogenesis in higher plant cells. American Zoologist, 1970, 10(3): 393-403 (doi: 10.1093/icb/10.3.393).
- Медведев С.С. Электрофизиология растений. СПб, 1998.
- Tattar T.A., Blanchard R.O. Electrophysiological research in plant pathology. Annual Review of Phytopathology, 1976, 14(1): 309-325 (doi: 10.1146/annurev.py.14.090176.001521).
- Поздняков А.И. Биоэлектрические потенциалы в системе почва растение. Почвоведение, 2013, 7: 813-813 (doi: 10.7868/S0032180X13070095).
- Опритов В.А., Тятыгин С.С., Ретивин В.Г. Биоэлектрогенез у высших растений. М., 1991.
- Moqsud M.A., Yoshitake J., Bushra Q.S., Hyodo M., Omine K., Strik D. Compost in plant microbial fuel cell for bioelectricity generation. Waste Management, 2015, 36: 63-69 (doi: 10.1016/j.wasman.2014.11.004).
- De Schamphelaire L., Cabezas A., Marzorati M., Friedrich M.W., Boon N., Verstraete W., Microbial community analysis of anodes from sediment microbial fuel cells powered by rhizodeposits of living rice plants. Applied and Environmental Microbiology, 2010, 76(6): 2002-2008 (doi: 10.1128/AEM.02432-09).
- Logan B. E. Microbial fuel cells. John Wiley & Sons, 2008.
- Kabutey F.T., Zhao Q., Wei L., Ding J., Antwi P., Quashie F.K., Wang W. An overview of plant microbial fuel cells (PMFCs): configurations and applications. Renewable and Sustainable Energy Reviews, 2019, 110: 402-414 (doi: 10.1016/j.rser.2019.05.016).
- El-Naggar M.Y., Wanger G., Leung K.M., Yuzvinsky T.D., Southam G., Yang J., Lau W.M., Nealson K.H., Gorby Y.A. Electrical transport along bacterial nanowires from Shewanella oneidensis MR-1. Proceedings of the National Academy of Sciences of the USA, 2010, 107(42): 18127-18131 (doi: 10.1073/pnas.1004880107).
- Sekar N., Ramasamy R.P. Electrochemical impedance spectroscopy for microbial fuel cell characterization. Journal of Microbial and Biochemical Technology, 2013: 6 (doi: 10.4172/1948-5948.S6-004).
- Lovley D.R., Holmes D.E. Electromicrobiology: the ecophysiology of phylogenetically diverse electroactive microorganisms. Nature Reviews Microbiology, 2022, 20(1): 5-19 (doi: 10.1038/s41579-021-00597-6).
- Timmers R.A., Rothballer M., Strik D.P., Engel M., Schulz S., Schloter M., Hartmann A., Hamelers B., Buisman C. Microbial community structure elucidates performance of Glyceria maxima plant microbial fuel cell. Applied Microbiology and Biotechnology, 2012, 94: 537-548 (doi: 10.1007/s00253-012-3894-6).
- Park D.H., Kim B.H. Growth properties of the iron-reducing bacteria, Shewanella putrefaciens IR-1 and MR-1 coupling to reduction of Fe (III) to Fe (II). Journal of Microbiology, 2001, 39(4): 273-278.
- Niessen J., Schröder U., Scholz F. Exploiting complex carbohydrates for microbial electricity generation — a bacterial fuel cell operating on starch. Electrochemistry Communications, 2004, 6(9): 955-958 (doi: 10.1016/j.elecom.2004.07.010).
- Xing D., Zuo Y., Cheng S., Regan J.M., Logan B.E. Electricity generation by Rhodopseudomonas palustris DX-1. Environmental Science & Technology, 2008, 42(11): 4146-4151 (doi: 10.1021/es800312v).
- Galushko A.S., Schink B. Oxidation of acetate through reactions of the citric acid cycle by Geobacter sulfurreducens in pure culture and in syntrophic coculture. Archives of Microbiology, 2000, 174: 314-321 (doi: 10.1007/s002030000208).
- Bond D.R., Lovley D.R. Reduction of Fe (III) oxide by methanogens in the presence and absence of extracellular quinones. Environmental Microbiology, 2002, 4(2): 115-124 (doi: 10.1046/j.1462-2920.2002.00279.x).
- Min B., Cheng S., Logan B.E. Electricity generation using membrane and salt bridge microbial fuel cells. Water Research, 2005, 39(9): 1675-1686 (doi: 10.1016/j.watres.2005.02.002).
- Chaudhuri S.K., Lovley D.R. Electricity generation by direct oxidation of glucose in mediatorless microbial fuel cells. Nature Biotechnology, 2003, 21: 1229-1232 (doi: 10.1038/nbt867).
- Rabaey K., Verstraete W. Microbial fuel cells: novel biotechnology for energy generation. Trends in Biotechnology, 2005, 23(6): 291-298 (doi: 10.1016/j.tibtech.2005.04.008).
- Rezaei F., Xing D., Wagner R., Regan J.M., Richard T.L., Logan B.E. Simultaneous cellulose degradation and electricity production by Enterobacter cloacae in a microbial fuel cell. Applied and Environmental Microbiology, 2009, 75(11): 3673-3678 (doi: 10.1128/AEM.02600-08).
- Deng H., Chen Z., Zhao F. Energy from plants and microorganisms: progress in plant-microbial fuel cells. ChemSusChem, 2012, 5(6): 1006-1011 (doi: 10.1002/cssc.201100257).
- He Z., Kan J., Wang Y., Huang Y., Mansfeld F., Nealson K.H. Electricity production coupled to ammonium in a microbial fuel cell. Environmental Science & Technology, 2009, 43(9): 3391-3397 (doi: 10.1021/es803492c).
- Higinbotham N. Electropotentials of plant cells. Annual Review of Plant Physiology, 1973, 24(1): 25-46 (doi: 10.1146/annurev.pp.24.060173.000325).
- Regmi R., Nitisoravut R., Ketchaimongkol J. A decade of plant-assisted microbial fuel cells: looking back and moving forward. Biofuels, 2018, 9(5): 605-612 (doi: 10.1080/17597269.2018.1432272).
- Поздняков А.И., Позднякова А.Д. Электрофизика почв. М., 2004.
- Кулешова Т.Э., Блохин Ю.И., Галль Н.Р., Панова Г.Г. Изучение градиента биоэлектрического потенциала в корнеобитаемой среде. In: Genetica, fiziologia şi ameliorarea plantelor. Ediția 7, 4-5 octombrie 2021, Chişinău. Chişinău, 2021: 38-41 (doi: 10.53040/gppb7.2021.09).
- Logan B.E., Regan J.M. Electricity-producing bacterial communities in microbial fuel cells. TrendsinMicrobiology, 2006, 14(12): 512-518 (doi: 10.1016/j.tim.2006.10.003).
- Strik D.P.B.T.B., Hamelers H.V.M., Snel J.F.H., Buisman C.J.N. Green electricity production with living plants and bacteria in a fuel cell. International Journal of Energy Research, 2008, 32: 870-876 (doi: 10.1002/er.1397).
- Bennetto H.P. Electricity generation by microorganisms. Biotechnology Education, 1990, 1(4): 163-168.
- Helder M., Chen W.-S., van der Harst E.J., Strik D.P.B.T.B., Hamelers H.B.V., Buisman C.J., Potting J. Electricity production with living plants on a green roof: environmental performance of the plant-microbial fuel cell. Biofuels, Bioproducts and Biorefining, 2013, 7(1): 52-64 (doi: 10.1002/bbb.1373).
- Timmers R.A., Strik D.P.B.T.B., Hamelers H.V., Buisman C.J. Electricity generation by a novel design tubular plant microbial fuel cell. Biomass and Bioenergy, 2013, 51: 60-67 (doi: 10.1016/j.biombioe.2013.01.002).
- Mohan S.V., Mohanakrishna G., Chiranjeevi P. Sustainable power generation from floating macrophytes based ecological microenvironment through embedded fuel cells along with simultaneous wastewater treatment. Bioresource Technology, 2011, 102(14): 7036-7042 (doi: 10.1016/j.biortech.2011.04.033).
- Rahimnejad M., Adhami A., Darvari S., Zirepour A., Oh S.-E. Microbial fuel cell as new technology for bioelectricity generation: a review. Alexandria Engineering Journal, 2015, 54(3): 745-756 (doi: 10.1016/j.aej.2015.03.031).
- Gulamhussein M., Randall D.G. Design and operation of plant microbial fuel cells using municipal sludge. Journal of Water Process Engineering, 2020: 38 (doi: 10.1016/j.jwpe.2020.101653).
- Wetser K., Sudirjo E., Buisman C., Strik D. Electricity generation by a plant microbial fuel cell with an integrated oxygen reducing biocathode. Applied Energy, 2015, 137: 151-157 (doi: 10.1016/j.apenergy.2014.10.006).
- Timmers R.A., Strik D.P.B.T.B., Hamelers H.V.M., Buisman C.J.N. Characterization of the internal resistance of a plant microbial fuel cell. Electrochimca Acta, 2012, 72: 165-171 (doi: 10.1016/j.electacta.2012.04.023).
- Helder M., Strik D.P.B.T.B., Timmers R.A., Raes S.M.T., Hamelers, H.V.M., Buisman C.J.N. Resilience of roof-top plant-microbial fuel cells during dutch winter. Biomass and Bioenergy, 2013, 51: 1-7 (doi: 10.1016/j.biombioe.2012.10.011).
- Ahn Y., Logan B.E. Altering anode thickness to improve power production in microbial fuel cells with different electrode distances. Energy and Fuels, 2013, 27(1): 271-276 (doi: 10.1021/ef3015553).
- Бичиашвили Т.Г., Цанава В.П., Соловьев Е.В., Маричев Г.А. Комплекс электрофизиологических и электрохимических методов диагностики физиологического состояния субтропических культур. В сб.: Биофизика растений и фитомониторинг. Л., 1990: 128-139.
- Кулешова Т.Э., Бушлякова А.В., Галль Н.Р. Неинвазивное измерение биоэлектрических потенциалов растений. Письма в журнал технической физики, 2019, 45(5): 6-8 (doi: 10.21883/PJTF.2019.05.47387.17541).
- Azri Y.M., Tou I., Sadi M., Benhabyles L. Bioelectricity generation from three ornamental plants: Chlorophytum comosum, Chasmanthe floribunda and Papyrus diffusus. International Journal of Green Energy, 2018, 15(4): 254-263 (doi: 10.1080/15435075.2018.1432487).
- Villaseñor J., Capilla P., Rodrigo M.A., Cañizares P., Fernández F.J. Operation of a horizontal subsurface flow constructed wetland-microbial fuel cell treating wastewater under different organic loading rates. Water Research, 2013, 47(17): 6731-6738 (doi: 10.1016/j.watres.2013.09.005).
- Кулешова Т.Э., Галль Н.Р., Галушко А.С., Панова Г.Г. Электрогенез растительно-микробного топливного элемента при параллельном и последовательном соединении ячеек. Журнал технической физики, 2021, 91(3): 510-518 (doi: 10.21883/JTF.2021.03.50531.185-20).
- Liu S., Song H., Wei S., Yang F., Li X. Bio-cathode materials evaluation and configuration optimization for power output of vertical subsurface flow constructed wetland — Microbial fuel cell systems. Bioresource Technology, 2014, 166: 575-583 (doi: 10.1016/j.biortech.2014.05.104).
- Sophia A.C., Sreeja S. Green energy generation from plant microbial fuel cells (PMFC) using compost and a novel clay separator. Sustainable Energy Technologies and Assessments, 2017, 21: 59-66 (doi: 10.1016/j.seta.2017.05.001).
- Timmers R.A., Strik D.P.B.T.B., Arampatzoglou C., Buisman C.J.N., Hamelers H.V.M. Rhizosphere anode model explains high oxygen levels during operation of a Glyceria maxima PMFC. Bioresource Technology, 2012, 108: 60-67 (doi: 10.1016/j.biortech.2011.10.088).
- Khudzari J.M., Kurian J, Gariépy Y., Tartakovsky B., Raghavan G.S.V. Effects of salinity, growing media, and photoperiod on bioelectricity production in plant microbial fuel cells with weeping alkaligrass. Biomass Bioenergy, 2018, 109: 1-9 (doi: 10.1016/j.biombioe.2017.12.013).
- Oon Y.-L., Ong S.-A., Ho L.-N., Wong Y.-S., Dahalan F.A., Oon Y.-S., Lehl H.K., Thung W.-E. Synergistic effect of upflow constructed wetland and microbial fuel cell for simultaneous wastewater treatment and energy recovery. Bioresource Technology, 2016, 203: 190-197 (doi: 10.1016/j.biortech.2015.12.011).
- Gilani S.R., Yaseen A., Zaidi S.R.A., Zahra M., Mahmood Z. Photocurrent generation through plant microbial fuel cell by varying electrode materials. Journal of the Chemical Society of Pakistan, 2016, 38(1): 17-27.
- Ueoka N., Sese N., Sue M., Kouzuma A., Watanabe K. Sizes of anode and cathode affect electricity generation in rice paddy-field microbial fuel cells. Journal of Sustainable Bioenergy Systems, 2016, 6(1): 10-15 (doi: 10.4236/jsbs.2016.61002).
- Chiranjeevi P., Mohanakrishna G., Mohan S.V. Rhizosphere mediated electrogenesis with the function of anode placement for harnessing bioenergy through CO2 sequestration. Bioresource Technology, 2012, 124: 364-370 (doi: 10.1016/j.biortech.2012.08.020).
- Oon Y.-L., Ong S.-A., Ho L.-N., Wong Y.-S., Dahalan F.-A., Oon Y.-S., Lehl H.K., Thung W.-E., Nordin N. Role of macrophyte and effect of supplementary aeration in up-flow constructed wetland-microbial fuel cell for simultaneous wastewater treatment and energy recovery. Bioresource Technology, 2017, 224: 265-275 (doi: 10.1016/j.biortech.2016.10.079).
- Mohan S.V., Mohanakrishna G., Chiranjeevi P. Sustainable power generation from floating macrophytes based ecological microenvironment through embedded fuel cells along with simultaneous wastewater treatment. Bioresource Technology, 2011, 102(14): 7036-7042 (doi: 10.1016/j.biortech.2011.04.033).
- Regmi R., Nitisoravut R., Charoenroongtavee S., Yimkhaophong W., Phanthurat O. Earthen pot-plant microbial fuel cell powered by vetiver for bioelectricity production and wastewater treatment. Clean - Soil, Air, Water, 2018, 46(3): 1700193 (doi: 10.1002/clen.201700193).
- Srivastava P., Yadav A.K., Mishra B.K. The effects of microbial fuel cell integration into constructed wetland on the performance of constructed wetland. Bioresource Technology, 2015, 195: 223-230 (doi: 10.1016/j.biortech.2015.05.072).
- Hubenova Y., Mitov M. Conversion of solar energy into electricity by using duckweed in direct photosynthetic plant fuel cell. Bioelectrochemistry, 2012, 87: 185-191 (doi: 10.1016/j.bioelechem.2012.02.008).
- Wetser K., Sudirjo E., Buisman C.J., Strik D.P.B.T.B. Electricity generation by a plant microbial fuel cell with an integrated oxygen reducing biocathode. Applied Energy, 2015, 137: 151-157 (doi: 10.1016/j.apenergy.2014.10.006).
- Nitisoravut R., Regmi R. Plant microbial fuel cells: a promising biosystems engineering. Renewable and Sustainable Energy Reviews, 2017, 76: 81-89 (doi: 10.1016/j.rser.2017.03.064).
- Jiang D., Li B., Jia W., Lei Y. Effect of inoculum types on bacterial adhesion and power production in microbial fuel cells. Applied Biochemistry and Biotechnology, 2010, 160: 182 (doi: 10.1007/s12010-009-8541-z).
- Kuzyakov Y., Domanski G. Carbon input by plants into the soil. Review. Journal of Plant Nutrition and Soil Science, 2000, 163(4): 421-431 (doi: 10.1002/1522-2624(200008)163:4<421::AID-JPLN421>3.0.CO;2-R).
- Hassan M.K., McInroy J.A., Kloepper J.W. The interactions of rhizodeposits with plant growth-promoting rhizobacteria in the rhizosphere: a review. Agriculture, 2019, 9(7): 142 (doi: 10.3390/agriculture9070142).
- Kuzyakov Y. Review: factors affecting rhizosphere priming effects. Journal of Plant Nutrition and Soil Science, 2002, 165(4): 382-396 (doi: 10.1002/1522-2624(200208)165:4<382::AID-JPLN382>3.0.CO;2-%23).
- Сваровская Н.А., Колесников И.М., Винокуров В.А. Электрохимия растворов электролитов. М., 2017.
- Кулешова Т.Э., Галль Н.Р. Динамика биоэлектрического потенциала в прикорневой зоне растений при поливах. Почвоведение, 2021, 3: 338-346 (doi: 10.31857/S0032180X21030084).
- Пасичный А.П., Карманов В.Г. Измерение биоэлектрического потенциала листьев высших растений бесконтактным способом. Вопросы экспериментальной биофизики и кибернетики растений. Труды АФИ, 1969, 24: 161-168.
- Коловский Р.А. Биоэлектрические потенциалы древесных растений. Новосибирск, 1980.
- Мелещенко С.Н. К вопросу о природе изменений электрических свойств растительной ткани при изменении внешних условий. Биофизика, 1965, 10: 78-98.
- Кулешова Т.Э., Галль Н.Р., Удалова О.Р., Панова Г.Г. Многофункциональный комплекс датчиков для фитомониторинга в условиях интенсивной светокультуры. Агрофизика, 2020, 4: 33-39.
- Cabezas A., Pommerenke B., Boon N., Friedrich M.W. Geobacter, Anaeromyxobacter and Anaerolineae populations are enriched on anodes of root exudate‐driven microbial fuel cells in rice field soil. Environmental Microbiology Reports, 2015, 7(3), 489-497 (doi: 10.1111/1758-2229.12277 ).
- Guan C.-Y., Tseng Y.-H., Tsang D.C.W., Hu A., Yu C.-P. Wetland plant microbial fuel cells for remediation of hexavalent chromium contaminated soils and electricity production. Journal of Hazardous Materials, 2019, 365: 137-145 (doi: 10.1016/j.jhazmat.2018.10.086).
- Oon Y.-L., Ong S.-A., Ho L.-N., Wong Y.-S., Oon Y.-S., Lehl H.K., Thung W.-E. Hybrid system up-flow constructed wetland integrated with microbial fuel cell for simultaneous wastewater treatment and electricity generation. Bioresource Technology, 2015, 186: 270-275 (doi: 10.1016/j.biortech.2015.03.014 ).
- Кулешова Т.Э., Галль Н.Р., Галушко А.С., Удалова О.Р., Вертебный В.Е., Панова Г.Г. Растительно-микробный топливный элемент на примере салата при культивировании методом панопоники. Аграрный научный журнал, 2021, 1: 24-28 (doi: 10.28983/asj.y2021i1pp24-28).
- Панова Г.Г., Удалова О.Р., Канаш Е.В., Галушко А.С., Кочетов А.А., Прияткин Н.С., Архипов М.В., Черноусов И.Н. Основы физического моделирования. Журнал технической физики, 2020, 90(10): 1633-1639 (doi: 10.21883/JTF.2020.10.49792.429-19).
- Ashok K., Babu M., Jula V., Mullai N.K. Impact of used battery disposal in the environment. Linguistics and Culture Review, 2021, 5(S1): 1276-1286.
- Zhang J., Yuan H., Abu-Reesh I.M., He Z., Yuan C. Life cycle environmental impact comparison of bioelectrochemical systems for wastewater treatment. Procedia CIRP, 2019, 80: 382-388 (doi: 10.1016/j.procir.2019.01.075).
- Sudirjo E., Buisman C.J.N., Strik D.P.B.T.B. Marine sediment mixed with activated carbon allows electricity production and storage from internal and external energy sources: a new rechargeable bio-battery with bi-directional electron transfer properties. Frontiers in Microbiology, 2019, 10: 934 (doi: 10.3389/fmicb.2019.00934).
- Liu B., Yan C., Si W., Sun X., Lu X., Ansorge-Schumacher, M., Schmidt O.G. Ultralong-discharge-time biobattery based on immobilized enzymes in bilayer rolled-up enzymatic nanomembranes. Small, 2018, 14(13): 1704221 (doi: 10.1002/smll.201704221).
- Adekunle A., Raghavan V., Tartakovsky B. Real-time performance optimization and diagnostics during long-term operation of a solid anolyte microbial fuel cell biobattery. Batteries, 2019, 5(1): 9 (doi: 10.3390/batteries5010009).
- Hussain Z., Zuhra, Rukh G., Zada A., Naz M.Y., Khan K.M., Shukrullah S., Sulaiman S.A. Construction of rechargeable bio-battery cells from electroactive antioxidants extracted from wasted vegetables. Cleaner Engineering and Technology, 2021, 5: 100342 (doi: 10.1016/j.clet.2021.100342).
- Rusyn I.B., Medvediev O.V., Valko B.T. Enhancement of bioelectric parameters of multi-electrode plant–microbial fuel cells by combining of serial and parallel connection. International Journal of Environmental Science and Technology, 2021, 18: 1323-1334 (doi: 10.1007/s13762-020-02934-3).
- Apollon W., Luna-Maldonado A.I., Kamaraj S.K., Vidales-Contreras J.A., Rodríguez-Fuentes H., Gómez-Leyva J.F., Aranda-Ruíz J. Progress and recent trends in photosynthetic assisted microbial fuel cells: a review. Biomass Bioenergy, 2021, 148: 106028 (doi: 10.1016/j.biombioe.2021.106028).
- Apollon W., Kamaraj S.-K., Silos-Espino H., Perales-Segovia C., Valera-Montero L.L., Maldonado-Ruelas V.A., Vázquez-Gutiérreza M.A., Ortiz-Medinab R.A., Flores-Beníteza S., Gómez-Leyva J.F. Impact of Opuntia species plant bio-battery in a semi-arid environment: demonstration of their applications. Applied Energy, 2020, 279: 115788 (doi: 10.1016/j.apenergy.2020.115788).
- Apollon W., Valera-Montero L.L., Perales-Segovia C., Maldonado-Ruelas V.A., Ortiz-Medina R.A., Gómez-Leyva F.F., Vázquez-Gutiérrez M.A., Flores-Benítez S., Kamaraj S.K. Effect of ammonium nitrate on novel cactus pear genotypes aided by biobattery in a semi-arid ecosystem. Sustainable Energy Technologies and Assessments, 2022, 49: 101730 (doi: 10.1016/j.seta.2021.101730).
- Brunelli D., Tosato P., Rossi M. Flora health wireless monitoring with plant-microbial fuel cell. Procedia Engineering, 2016, 168: 1646-1650 (doi: 10.1016/j.proeng.2016.11.481).
- Jayaraman P.P., Yavari A., Georgakopoulos D., Morshed A., Zaslavsky A. Internet of things platform for smart farming: Experiences and lessons learnt. Sensors, 2016, 16(11): 1884 (doi: 10.3390/s16111884).
- Schievano A., Colombo A., Grattieri M., Trasatti S.P., Liberale A., Tremolada P., Pino C., Cristiani P. Floating microbial fuel cells as energy harvesters for signal transmission from natural water bodies. Journal of Power Sources, 2017, 340: 80-88 (doi: 10.1016/j.jpowsour.2016.11.037).
- Osorio-de-la-Rosa E., Vazquez-Castillo J., Castillo-Atoche A., Heredia-Lozano J., Castillo-Atoche A., Becerra-Nunez G., Barbosa R. Arrays of plant microbial fuel cells for implementing self-sustainable wireless sensor networks. IEEE Sensors Journal, 2021, 21(2): 1965-1974 (doi: 10.1109/JSEN.2020.3019986).
- Koók L., Rózsenberszki T., Nemestóthy N., Bélafi-Bakó K., Bakonyi P. Bioelectrochemical treatment of municipal waste liquor in microbial fuel cells for energy valorization. Journal of Cleaner Production, 2016, 112(5): 4406-4412 (doi: 10.1016/j.jclepro.2015.06.116).
- Wang X., Feng Y.J., Lee H. Electricity production from beer brewery wastewater using single chamber microbial fuel cell. Water Science & Technology, 2008, 57(7): 1117-1121 (doi: 10.2166/wst.2008.064).
- Dong Y., Qu Y., He W., Du Y., Liu J., Han X., Feng Y. A 90-liter stackable baffled microbial fuel cell for brewery wastewater treatment based on energy self-sufficient mode. Bioresouce Technology, 2015, 195: 66-72 (doi: 10.1016/j.biortech.2015.06.026).
- An Z., Feng Q., Zhao R., Wang X. Bioelectrochemical methane production from food waste in anaerobic digestion using a carbon-modified copper foam electrode. Processes, 2020, 8(4): 416 (doi: 10.3390/pr8040416).
- Pushkar P., Mungray A.K. Real textile and domestic wastewater treatment by novel cross-linked microbial fuel cell (CMFC) reactor. Desalination and Water Treatment, 2016, 57(15): 6747-6760 (doi: 10.1080/19443994.2015.1013994).
- Nancharaiah Y.V., Venkata Mohan S., Lens P.N.L. Metals removal and recovery in bioelectrochemical systems: a review. Bioresource Technology, 2015, 195: 102-114 (doi: 10.1016/j.biortech.2015.06.058).
- Yu H., Zhao Q., Liu X., Meng F., Ruan L., Sun T., Liu W., Zhu Y., Li W., Meng F., Liang Z. Enhanced chromium recovery and simultaneous sludge degradation in a novel bioelectrochemical system assembled with bio/abio-cathodes. Separation and Purification Technology, 2020, 250: 117229 (doi: 10.1016/j.seppur.2020.117229).
- Nancharaiah Y.V., Lens P.N.L., Mohan S. Recent advances in nutrient removal and recovery in biological and bioelectrochemical systems. Bioresource Technology, 2016, 215: 173-185 (doi: 10.1016/j.biortech.2016.03.129).
- Yang N., Zhang G., Luo H., Xiong X., Li D. Integrated simultaneous nitrification/denitrification and comammox consortia as efficient biocatalysts enhance treatment of domestic wastewater in different up-flow bioelectrochemical reactors. Bioresource Technology, 2021, 339: 125604 (doi: 10.1016/j.biortech.2021.125604).