








УДК 631.46:579.64:[576.8.095.38+581.557(575)
СЕЛЬСКОХОЗЯЙСТВЕННАЯ МИКРОБИОЛОГИЯ КАК ОСНОВА ЭКОЛОГИЧЕСКИ УСТОЙЧИВОГО АГРОПРОИЗВОДСТВА: ФУНДАМЕНТАЛЬНЫЕ И ПРИКЛАДНЫЕ АСПЕКТЫ
И.А. ТИХОНОВИЧ, Н.А. ПРОВОРОВ
Основу сельскохозяйственной микробиологии составляет изучение микробных сообществ, обитающих в растениях, животных и почвах. Поскольку растения и животные создают на поверхностях, в тканях и в клетках разнообразные ниши для размещения микробных партнеров, гены, контролирующие развитие этих ниш, служат важнейшими детерминантами активности хозяйственно ценных микроорганизмов. Знания об их генетике, физиологии, молекулярной биологии и экологии позволяют создавать системы экологически устойчивого агропроизводства, в которых продуктивность растений и животных повышена за счет изменения состава и свойств их микробных симбионтов, экологически опасные агрохимикаты (минеральные удобрения, пестициды) полностью или частично замещены препаратами микроорганизмов, снижена энергоемкость производства и повышено качество продукции.
Ключевые слова: сельскохозяйственная микробиология, микробно-растительные взаимодействия, азотфиксирующие симбиозы, арбускулярная микориза, эндофитные микроорганизмы, ризосферные ростсимулирующие бактерии, биоконтроль вредителей и патогенов, микробиота рубца, метагеномы почвы, экологически устойчивое земледелие.
В 2011 году исполняется 120 лет с начала развития сельскохозяйственной микробиологии в России: в 1891 году в Санкт-Петербурге при Департаменте земледелия и государственных имуществ была организована Бактериологическая лаборатория, задачей которой стало создание микробиологических методов борьбы с мышевидными грызунами. Вскоре к производству «мышеубивающих» препаратов добавилось изучение инфекционных болезней растений и животных, а также микробиологических основ изготовления молочных продуктов и вина. После преобразования лаборатории во Всесоюзный (ныне Всероссийский) НИИ сельскохозяйственной микробиологии по инициативе его организатора и первого директора С.П. Костычева началось активное исследование полезных взаимодействий микроорганизмов с растениями, которое и в настоящее время составляет основу научной тематики института. В результате развития этого комплекса направлений сельскохозяйственная микробиология переросла в самостоятельную научную дисциплину, органически сочетающую фундаментальные и прикладные аспекты. При этом речь идет не только о передаче теоретических знаний в производство: огромную роль играют обратные связи практики и теории, стимулирующие постановку новых фундаментальных задач.
Сельскохозяйственная микробиология стала особенно актуальной в связи с необходимостью экологизации сельскохозяйственного производства. Интенсивные агротехнологии, обеспечив «зеленую революцию» середины XX века, привели к непредвиденным последствиям — глобальному загрязнению биосферы, неблагоприятным изменениям климата, утрате биоразнообразия в большинстве природных экосистем и в конечном итоге к снижению качества жизни народонаселения Земли. Поэтому все большее внимание уделяется развитию экологически устойчивых сельскохозяйственных систем, в которых продуктивность растений и животных обеспечивается использованием их биологических возможностей, при минимальном применении экологически опасных агрохимикатов — минеральных удобрений, пестицидов, регуляторов роста (1, 2). Один из основных способов достижения этой цели — частичное или полное замещение агрохимикатов препаратами симбиотических микроорганизмов, которые в природе успешно обеспечивают своих хозяев питательными веществами и защищают их от биотических и абиотических стрессов (3, 4).
Сельскохозяйственное производство зависит от активности разнообразных микроорганизмов, обеспечивающих питание и развитие растений и животных, биоконтроль вредителей (насекомые-фитофаги, грызуны) и сорных растений, плодородие почвы (табл.).
| Наиболее изученные примеры сельскохозяйственно значимых симбиозов растений и животных с микроорганизмами | |||
Тип симбиоза |
Функция |
Хозяин |
Микросимбионт |
Р а с т и т е л ь н о - м и к р о б н ы е в з а и м о д е й с т в и я |
|||
Трофический |
Фиксация N2: |
|
|
Ассимиляция нерастворимых фосфатов почвы |
80-90 % наземных растений |
Гломусовые грибы |
|
Защитный |
Подавление растительноядных |
Разные растения |
Спорыньевые грибы (Clavi-cipitaceae), Clavibactertoxicus |
Подавление патогенных грибов |
Разные растения |
Pseudomonas spp. |
|
Биоконтрольный |
Подавление сорняков |
Amaranthus, Orobanche, Senecio |
Различные патогенные грибы и бактерии (24, 25) |
В з а и м о д е й с т в и я ж и в о т н ы х с м и к р о о р г а н и з м а м и |
|||
Трофический |
Расщепление целлюлозы с одновременной утилизацией азотных шлаков хозяина и синтезом белка |
Жвачные животные (микробиота рубца) |
Многочисленные |
Защитный |
Активация иммунной системы хозяина |
||
Биоконтрольный |
Подавление насекомых-фитофагов Подавление грызунов |
Lepidoptera, Coleoptera, Diptera |
Bacillus thuringiensis |
Исторически сельскохозяйственная микробиология возникла на основе ветеринарии: начало работ по инфекционным болезням тутового шелкопряда, вызванным грибом Beauveriabassiana, датировано первой половиной XIX века (5). Однако методологически она наиболее тесно связана с почвенной микробиологией, поскольку большинство хозяйственно значимых симбиотических микроорганизмов представляют собой продукты онтогенетической трансформации и эволюции почвенной микробиоты (рис. 1). Изучение почвенных микроорганизмов инициировал С.Н. Виноградский. Он разделил их на метаболически активные организмы (R-стратеги), которые ассимилируют неорганические, низкомолекулярные органические вещества и быстро ферментируют высокомолекулярные органические соединения — белки, целлюлозу, пектин, хитин («зимогенная» микрофлора), и метаболически малоактивные организмы (k-стратеги), способные к деструкции и синтезу гумусовых веществ («аутохтонная» микрофлора) (6). Основной объект почвенной микробиологии — зимогенная микрофлора, доступная для изучения современными молекулярно-биологическими методами. На ее хозяйственное значение первым обратил внимание С.П. Костычев, связавший активность быстро развивающихся почвенных микроорганизмов с жизнедеятельностью растений (7). Подразумевалось, что растения служат источниками питательных субстратов — быстроусвояемых корневых экссудатов и медленнее утилизируемых остатков отмирающих органов. Используя эти субстраты, почвенные микробы формируют биологически активное окружение растений, поставляющее генетические ресурсы для эволюции симбиотически специализированных форм.
|
Рис. 1. Миграция сельскохозяйственно важных микроорганизмов в биоценозе. Широкими стрелками показаны основные направления миграции (белые — преобразование различных типов микробных сообществ, черные — их трансформация в двухкомпонентные симбиозы), пунктирные стрелки — гипотетические направления миграции, изогнутые — переход микроорганизмов из животных в почву. Эндофитная микрофлора разделена на две группы, обитающие в корнях и стеблях (1) и в листьях (2), с разными источниками инфицирования. П и т а н и е с п о- |
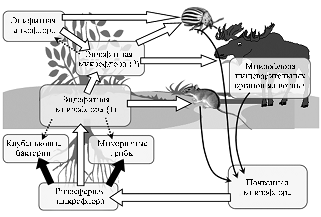
мируют симбиозы с разнообразными микроорганизмами, обеспечивающими питание своих хозяев. Значимость этих симбиозов определяется тем, что большинство высших организмов не может полностью обеспечить свое полноценное питание, испытывая дисбаланс основных его элементов: растения и животные-фитофаги получают избыток углерода, однако испытывают дефицит других макроэлементов, в первую очередь азота и фосфора. Наилучшими моделями для изучения трофических симбиозов служат двухкомпонентные растительно-микробные системы. Их мутуалистическая природа основана на положительных обратных связях партнеров: например, растения снабжают продуктами фотосинтеза преимущественно те части корня, из которых активно поступает азот (клубеньки) или фосфор (микоризованные участки кортекса). Установление подобных связей основано на системной регуляции, которая в случае бобово-ризобиального симбиоза включает элементы, как общие с системами защиты от патогенов (например, салициловую и жасмоновую кислоты), так и не участвующие в этой защите (гены группы CLAVATA, контролирующие формирование меристем, ген HAR1, участвующий в образовании боковых корней) (8). Важно отметить, что у бобовых системный ответ, регулирующий образование клубеньков, формируется в листьях (9), что отражает зависимость энергоемких симбиотических процессов от фотосинтеза. В то же время регуляция усвоения связанного азота (нитратов) в основном ограничена корнем (8).
Поскольку растения и животные образуют на своих поверхностях, в тканях, внутренних полостях, а иногда и в клетках разнообразные ниши для размещения микробных партнеров, гены хозяина, ответственные за развитие этих ниш, могут рассматриваться как важнейшие детерминанты экологических функций микросимбионтов. Изучение двухкомпонентных симбиозов позволило нам описать феномен генетической интеграции неродственных организмов, приводящей к образованию надорганизменных систем наследственности — симбиогеномов (4, 10).
На субклеточном уровне симбиотрофное питание растений азотом включает диалог нескольких партнеров — растения-хозяина, его перманентных эндосимбионтов (пластиды и митохондрии, эволюционно возникшие соответственно из цианобактерий и аэробных a-протеобактерий) и временных симбионтов (бактероиды ризобий, находящиеся в симбиосомах) (рис. 2). Таким образом, двухкомпонентная на первый взгляд бобово-ризо-биальная симбиоистема оказывается многокомпонентной и по степени функциональной целостности превосходит эндофитные сообщества, которые будут рассмотрены ниже.
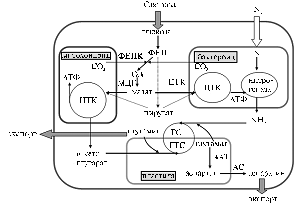 |
Рис. 2. Схема взаимодействия эндосимбиотических компонентов клетки при симбиотрофном питании растений азотом: ФЕП — фосфоенолпируват, ОА — оксалоацетат, ФЕПК — фосфоенолпируваткарбоксилаза, МДГ — малатдегидрогеназа, ГС — глутаминсинтетаза, ГТС — глутаматсинтаза, ААТ — аспартатаминотрансфераза, АС — аспарагинсинтаза, ЦТК — цикл трикарбоновых кислот. |
Экологически наиболее важная форма трофического симбиоза — арбускулярная микориза (АМ), которую наземные растения образуют с гломусовыми грибами (Glomeromycota). Благодаря этому симбиозу пода-вляющее большинство (до 90 %) наземных растений получают основную часть минерального (в первую очередь фосфорного) питания (11). Более того, АМ — эволюционный предшественник для большинства корневых симбиозов: растительные гены, гомологичные генам регуляции АМ, участвуют в развитии клубеньков, в формировании ризосферных ассоциаций, а также в защите от патогенов (12). К числу таких генов относятся факторы сигнального взаимодействия, например гены рецепторных киназ, которые играют важную роль практически во всех типах микробно-растительного симбиоза (13). Важно отметить, что в природе гломусовые грибы колонизируют растения в тесной кооперации с сателлитными бактериями, населяющими поверхности гиф и их цитоплазму, что делает АМ многокомпонентной симбиотической системой (14). Некоторые виды ризобий представляют собой продукты эволюции этих сателлитов, обеспечившей приобретение ими собственной, независимой от гриба симбиотической функции — снабжения растений азотом (15).
Несмотря на очевидные отличия от клубенькового симбиоза, ризосферные ассоциации, образуемые азотфиксирующими микроорганизмами, например Azospirillum (16), имеют с ним ряд общих черт. Так, азоспириллы образуют с хозяевами положительные обратные связи — стимулируют развитие корней с помощью фитогормонов (главным образом, ауксинов), а получаемые в результате усиленной корневой экссудации С-соединения используют для покрытия энергетической стоимости N2-фиксации и размножения бактерий (17). Однако ризосферные азотфиксаторы не могут обеспечить такую же высокую эффективность симбиотрофного питания растений азотом, как клубеньковые, поскольку в корень поступает лишь небольшая часть фиксируемого в ризосфере азота. Основные последствия от инокуляции растений азоспириллами связаны с продукцией гормонов, стимулирующих поглотительную активность корней: эффекты этой стимуляции и N2-фик-сации были разделены с помощью изотопных (15N) методов (18).
У эндофитных N2-фиксаторов, обитающих в тканях или в проводящей системе растений, эффективность выше, чем у ризосферных, так как продукты нитрогеназной активности эндофитов не рассеиваются в окружающей среде. Наиболее изученные эндофиты — представители родов Azoarcus, Gluconacetobacter, Herbaspirillum; их практическое значение определяется способностью к колонизации злаковых растений, включая рис и пшеницу (19). Однако общая эффективность N2-фиксации у эндофитов значительно ниже, чем у ризобий, для которых подача энергии и ассимиляция продуктов фотосинтеза обеспечиваются специализированными структурами клубенька.
Некоторые эндофиты проникают в растения при помощи векторных организмов, которые служат для них промежуточными или даже основными хозяевами. Так, Azoarcus — эндосимбионт гриба-аскомицета, обитающего в тканях злака Leptochloafusca, из которого часто выделяют этот азотфиксатор. Хотя штаммы Azoarcus легко культивируются на лабораторных средах, в почвах они обычно не выявляются, а с грибом образуют стабильные ассоциации, в которых развивают особые субклеточные структуры, содержащие нитрогеназу, — диазосомы (20). Находясь в растениях, Azoarcus фиксирует N2, переходя в некультивируемые формы, однако диазосомы при этом не образуются. Таким образом, эндофитная N2-фиксирующая система фактически трехкомпонентна, поскольку в ней роль Azoarcus заключается не только в улучшении азотного питания растения, но и в создании оптимальных условий для развития гриба-вектора.
М и к р о б и о л о г и ч е с к и й к о н т р о л ь в р е д и т е л е й: с и м б и
о з ы, о с н о в а н н ы е н а а н т а г о н и з м е. Образуя большое количество С-соеди-нений, растения привлекают многочисленных вредителей — патогенов и растительноядных животных. Для нейтрализации этих антагонистов растения поддерживают разнообразных защитных симбионтов. Их биоконтрольные функции выполняются благодаря двум механизмам — прямому подавлению вредителей (синтез токсинов, антибиотиков, литических экзоферментов; конкуренция за питательные субстраты) и индукции защитных реакций хозяина.
У защитных симбионтов способность к подавлению вредителей растений могла возникнуть до начала активного взаимодействия с самими растениями — в ходе коэволюции микроорганизмов с фитопатогенами или фитофагами, то есть растения культивируют уже сложившихся «врагов своих врагов». Например, Pseudomonas и Serratia обладают способностью к микофагии, лизируя гифы фузариума внеклеточными хитиназами. Многие штаммы аскомицетов рода Cordyceps являются бессимптомными эндо- и эпифитами растений и проходят полное развитие в теле насекомого-жертвы (21). Непрямое подавление патогенов часто связано с малоспецифическими эффектами: многие симбиотические организмы (ризобии, микоризные грибы, ризосферные бактерии) проникают в растения без образования некротических зон, вызывая системную устойчивость к патогенам (22). Защитные эндофиты, как и азотфиксирующие, обычно проникают в растение-хозяина с помощью организмов-векторов, которые представляют собой регулярных сожителей растений. Примером служит коринеформная бактерия Clavibacter toxicus: она проникает в растение вместе с нематодой Anguina, колонизируя образуемые ею галлы, откуда затем системно распространяется по всему растению (23). Присутствие этих эндофитов делает растение токсичным для животных, и таким образом Clavibacter «охраняет» его как удобную нишу для нематоды-вектора. Фактически при этом формируется многокомпонентная система, в которой разные организмы (бактерии, нематоды, животные-фитофаги) конкурируют за С-метаболиты хозяина, а тот, в свою очередь, благодаря динамичным обратным связям обеспечивает распределение С-ресурсов в пользу наиболее полезных симбионтов.
Естественные антагонисты вредителей растений могут быть использованы для биоконтроля независимо от того, обладают ли они способностью быть симбионтами растений. Один из основоположников сельскохозяйственной микробиологии С.С. Мережковский (26) для подавления грызунов (мышей, крыс, сусликов) предложил применять вид Salmonella enteritidis, не опасный для человека и сельскохозяйственных животных. Отметим, что некоторые бактерии, традиционно используемые для избирательного подавления определенных вредителей, оказались носителями весьма широкого спектра биоконтрольных функций. Так, Bacillus thuringiensis — вид, широко применяемый для подавления вредных насекомых (27, 28), активен также против многих фитопатогенных грибов (29). Другой пример полифункциональных симбионтов растений — штаммы Streptomyces, которые используются против фитопатогенов, но часто проявляют активность и против паутинных клещей (30, 31).
М н о г о к о м п о н е н т н ы е с и м б и о т и ч е с к и е с о о б щ е с т в а.
В предыдущих разделах мы отмечали, что двухкомпонентные симбиотические системы растений (бобово-ризобиальный симбиоз, арбускулярная микориза) представляют собой либо части более сложных многокомпонентных полифункциональных сообществ, либо продукты их эволюции. В последние годы такие сообщества стали объектами активного молекулярно-генетического исследования, основанного на подходах метагеномики, в первую очередь на анализе генов, кодирующих эволюционно консервативные рибосомные РНК (32). Оказалось, что все растения поддерживают в своих тканях эндофитную микрофлору, состоящую из сотен видов и почти столь же разнообразную, как и почвенные сообщества, хотя отличающуюся от них по таксономической структуре. Некоторые эндофиты генетически и функционально охарактеризованы (N2-фиксаторы Azoarcus, защитные симбионты Clavibacter), однако функции большинства из них (в первую очередь, некультивируемых форм) остаются неизвестными.
Изучение микробных сообществ растений показало, что концепция микробиома, изначально предложенная J. Lederberg с соавт. (33) для характеристики совокупного генома кишечной микрофлоры человека, может быть с полным основанием распространена на микробные сообщества растений. У них (в отличие от животных) эндосимбиотические сообщества распределены по разным органам и тканям, что делает взаимодействия с микроорганизмами чрезвычайно разнообразными. Основные функции эндофитных сообществ заключаются в биоконтроле патогенов и вредителей, а также в освобождении растений от поступающих извне ксенобиотиков, а возможно, и от собственных метаболических шлаков (32, 34), что весьма актуально в отсутствие у растений выделительных систем. Способность к фиксации N2 характерна лишь для некоторых эндофитов, так как nif-гены выявляются у небольшой части их видов. Однако эндофиты могут выполнять и другие трофические функции, включая ассимиляцию почвенных источников азота, фосфора и железа, а также трансформацию и перераспределение метаболитов между частями растения, что в определенной степени компенсирует отсутствие у него пищеварительных органов. Важной функцией эндофитов, особенно в условиях стрессов, может быть регуляция развития растений посредством активации синтеза гормонов, витаминов и других биологически активных веществ.
Основным механизмом формирования эндофитной микрофлоры служит инфицирование растений через разрывы эпидермиса, возникающие при росте боковых корней (35). Поэтому наиболее богатая микрофлора выявляется в корнях, из которых микроорганизмы мигрирует (главным образом, по проводящим сосудам) в надземные органы. У бактерий в этой миграции участвуют многие компоненты клетки, включая структуры клеточной стенки и капсулы, пили и жгутики, а также белковые эффекторы, выделяемые через различные секреторные системы (34). Однако в надземных органах имеются и собственные очаги проникновения инфекции: например, микроорганизмы могут попадать в растения через устьица, включаясь в состав микрофлоры листьев и стеблей. Наконец, существенную роль в формировании эндофитной микрофлоры играет вертикальная передача микроорганизмов через семена, а также их интродукция проникающими в растение векторными организмами — беспозвоночными или грибами.
Таким образом, растение обладает двумя типами симбиоза — локализованным и дисперсным (системным), функции которых регулируются с помощью положительных и отрицательных обратных связей. Эти связи хорошо изучены в случае локализации симбионтов в определенных частях растения: например, оно наиболее активно снабжает продуктами фотосинтеза те участки корня, из которых в надземную часть поступают значительные количества азота или фосфора (15). Механизмы контроля эндофитного микробного сообщества мало изучены, хотя показано, что важную роль здесь может играть регуляция по механизму quorum sensing, определяющему автоиндукцию или авторепрессию клеточных функций бактерий с помощью лактонов гомосерина. Растения могут вмешиваться в эту регуляцию, синтезируя сигнальные факторы, которые имитируют действие бактериальных авторегуляторов (36).
Важно отметить, что роль строгой локализации в организме хозяина для определения биологического эффекта взаимодействия различна при трофических и защитных симбиозах. Для трофических симбионтов, которые обычно не проявляют тесного родства с фитопатогенами, строгая локализация в организме хозяина сопряжена с максимальной экологической эффективностью. Например, у бобовых клубеньковые формы N2-фиксирующего симбиоза с ризобиями более эффективны, чем бесклубеньковые (внутрикорневые эндофитные системы) (37). Возникновение сложных механизмов проникновения в корень, например переход от инфицирования через разрывы эпидермиса к поглощению ризобий через корневые волоски, индуцируемому бактериальными Nod-факторами (38), определяет повышение эффективности N2-фиксакции.
Иное соотношение биологических эффектов системных и локализованных форм типично для защитных симбионтов. Например, у спорыньевых грибов, для которых характерны тесные эволюционные связи с фитопатогенами, наиболее полезные для хозяина формы обычно распределены по его организму системно (Neotyphodium, Epichloe), а усиление антагонистического действия (Claviceps) сопряжено со строгой локализацией микроорганизма в определенных органах растения. Такая эволюционная стратегия согласуется с данными о мутантах корневого патогена Colletotrichum magna, которые приобрели свойства защитных эндосимбионтов, сопряженные с колонизацией верхней части стебля (39). В растениях банана выявлены эндофитные штаммы Fusarium oxysporum, обеспечивающие контроль эндопаразитических нематод, обусловленный как их прямым подавлением микотоксинами, так и индуцированными защитными реакциями (40). Эти примеры показывают, что многие широко распространенные патогены могли возникать из защитных симбионтов в связи с дисбалансом регуляторных связей партнеров, вызванным доместикацией и селекцией растений.
У животных в отличие от растений микробиомы локализованы в особых органах, например в пищеварительных полостях, так как иммунные системы не позволяют микробам распространяться в тканях: у животных генетический гомеостаз поддерживается во внутренней среде более строго, чем у растений. Наиболее изученным примером микробиома животных служит сообщество рубца, обеспечивающее деструкцию растительных полимеров (целлюлозы, гемицеллюлозы, пектина), а также сопряженную с ней рециклизацию азотных шлаков и основанный на ней синтез белка (41). Активизация иммунной системы животных относится к важным функциям микрофлоры рубца. Эта система ограничивает распространение симбионтов внутренними полостями и поверхностью организма, тогда как у растений симбионты распространяются по многим тканям: растения толерантнее животных к наличию в организме симбиотической микрофлоры, поскольку более зависимы от нее.
И с п о л ь з о в а н и е в а г р о п р о и з в о д с т в е и б и о т е х н о л о г и и. Основой использования микроорганизмов в сельском хозяйстве служит эволюционно сложившаяся зависимость растений и растительноядных животных от симбиотических функций, выполняемых разнообразными микросимбионтами. При взаимодействии с растениями и животными они функционально замещают агрохимикаты (средства питания и защиты), что и определяет важность вклада микробиологии в растениеводство и животноводство. Однако это замещение происходит, как правило, частично и лишь иногда может быть полным — например, у эволюционно молодых бобовых культур, сохранивших природный симбиотический потенциал (42). У большинства бобовых наибольшая урожайность достигается при комбинированном применении обоих источников азота (отметим, что энергетическая стоимость фиксации N2 и усвоения нитратов различается не более чем на 10 %) (43, 44). В этом случае использование трофических симбионтов должно базироваться на миксотрофности растений, то есть на одновременном использовании симбиотически фиксированного и связанного азота.
К сожалению, доместикация и селекция растений привели к резкому снижению адаптивного резерва формируемых ими симбиосистем: например, у бобовых культур произошло резкое нарушение баланса между симбиотрофным и автотрофным типами азотного питания (42). Одним из следствий этого стало преобразование клубеньковых бактерий в не фиксирующие N2 «симбионты-обманщики», которые блокируют инокуляцию растений высокоактивными производственными штаммами (45).
В настоящее время для приготовления препаратов используется широкий круг симбиотических микроорганизмов, стимулирующих развитие растений и животных (46). Для улучшения трофических и защитных микробно-растительных симбиозов должны быть использованы различные подходы. Трофические симбиозы требуют эффективной колонизации растений, специфичной по отношению к генотипам обоих партнеров, что наиболее убедительно показано для бобово-ризобиального симбиоза. В защитных симбиозах целесообразно сочетание прямого и непрямого подавления патогенов. В этом случае специфичность микросимбионтов по отношению к хозяину не столь важна, как в трофических симбиозах, однако для подавления патогенов необходима специфичность микроорганизмов по отношению к их генотипам. Роль индукторов защитных реакций растения способны играть не только живые микроорганизмы, но и их очищенные метаболиты, например хитозаны. Некоторые из них могут быть получены с помощью систем синтеза Nod-факторов в гетерологичной бактерии Escherichiacoli (47). Применение микробных сигналов или их синтетических аналогов, регулирующих развитие и адаптивные функции растений, относится к перспективным областям сельскохозяйственной биотехнологии.
Создание микробных препаратов — только первый этап использования сельскохозяйственно ценных микроорганизмов. В дальнейшем необходимо переходить к биоинженерии сложных полифункциональных систем. Началом таких работ может быть создание многокомпонентных инокулятов — аналогов природных микробиомов растений. Например, целесообразным представляется сочетание симбионтов, снабжающих растения азотом и фосфором, что обеспечит сбалансированное питание растений. Универсальные Sym-гены бобовых, которые отвечают за мониторинг симбиотических микробов, изначально, возможно, были факторами контроля за многокомпонентными эндофитными сообществами. Однако условием для одновременного поддержания большого числа микроорганизмов служит создание генотипов растений, способных обеспечивать энергией высокую метаболическую активность одновременно у нескольких типов эндосимбионтов (48, 49).
Развитие сельскохозяйственной микробиологии должно учитывать экологические и генетические риски широкого распространения интродуцированных микроорганизмов в агросистемах. Один из таких рисков — поддержание в растениях бактерий и грибов, патогенных для человека (включая виды Burkholderia, Escherichia, Klebsiella, Salmonella, Staphylococcus). К счастью, было показано, что растения люцерны или арабидопсиса слабо колонизируются патогенными для человека штаммами бактерий, хотя активно заселяются непатогенными формами, лишенными флагелл и систем экскреции белков (34). Однако в колонизацию некоторых растений энтеробактериями могут быть вовлечены поверхностные структуры, участвующие в инфицировании человека (50), что указывает на необходимость тщательного мониторинга эндофитных сообществ, особенно в тех растениях, которые человек употребляет в пищу в сыром виде.
Итак, стремительное расширение знаний о взаимосвязях микроорганизмов с растениями и животными создает возможность для перехода к системам экологически устойчивого земледелия, в которых производство продукции экономически более выгодно, чем в системах интенсивного земледелия, и осуществляется при минимальной нагрузке на окружающую среду. Эффективное управление симбиотическими сообществами должно быть основано на целостности микробного населения агроценоза, связанной с его циркуляцией в системе ниш, предоставляемых растениями, животными и почвой (51, 52). Анализ механизмов этой циркуляции позволит не только эффективно использовать микроорганизмы в сельском хозяйстве, но и контролировать эколого-генетические последствия их широкомасштабной интродукции в агроценозы.
Л и т е р а т у р а
1. R e g a n o l d J.P., P a p e n d i c k R.I., P a r r J.F. Sustainable agriculture. Sci. Amer., 1990, 262: 112-120.
2. V a n c e C.P. Symbiotic nitrogen fixation and phosphorous acquisition. Plant nutrition in the world of declining renewable resources. Plant Physiology, 2001, 127: 390-397.
3. H i g a T., P a r r J.F. Beneficial and effective microorganisms for a sustainable agriculture and environment. Intern. Nature Farming Research Center, Atami, Japan, 1994: 1-16.
4. Т и х о н о в и ч И.А., П р о в о р о в Н.А.Симбиозы растений и микроорганизмов: молекулярная генетика агросистем будущего. СПб, 2009.
5. Ш т е й н х а у з Э. Микробиология насекомых. М., 1950.
6. В и н о г р а д с к и й С.Н. Микробиология почвы. М., 1952.
7. К о с т ы ч е в С.П. Новейшие исследования по биодинамике почв. Природа, 16: 355-372.
8. F e r g u s o n B.J., I n d r a s u m u n a r A., H a y a s h i S. e.a. Molecular analysis of legume nodule development and autoregulation. J. Integr. Plant Biol., 2010, 52: 61-76.
9. K i n k e m a M., S c o t t P.T., G r e s s h o f f P.M. Legume nodulation: successful symbiosis through short- and long-distance signaling. Funct. Plant Biol., 2006, 33: 707-721.
10. Т и х о н о в и ч И.А., П р о в о р о в Н.А. Симбиогенетика микробно-растительных взаимодействий. Экологическая генетика, 2003, 1: 36-46.
11. S m i t h S.E., R e a d D.J. Mycorrhizal symbiosis. London, 2008.
12. П р о в о р о в Н.А., Б о р и с о в А.Ю., Т и х о н о в и ч И.А. Сравнительная генетика и эволюционная морфология симбиозов растений с микробами-азотфиксаторами и эндомикоризными грибами. Журн. общ. биол., 2002, 63: 451-472.
13. D a r d i c k C., R o n a l d P. Plant and animal pathogen recognition receptors signal through non-RD kinases. PLOS Pathogens, 2006, 2: 14-28.
14. A r t u r s s o n V., F i n l a y R.D., J a n s s o n J.K. Interactions between arbuscular mycorrhizal fungi and bacteria and their potential for stimulating plant growth. Environ. Microbiol., 2006, 8: 1-10.
15. P r o v o r o v N.A., V o r o b y o v N.I. Evolutionary genetics of plant-microbe symbioses /I.A. Tikhonovich (ed.). N.Y., 2010.
16. D ö b e r e i n e r J. Isolation and identification of root associated diazotrophs. Plant and Soil, 1988, 110: 207-212.
17. K r a v c h e n k o L.V., L e o n o v a E.I., T i k h o n o v i c h I.A. Effect of root exudates of non-legume plants on the response of auxin production by associated diazotrophs. Microb. Releases, 1994, 2: 267-271.
18. H a r d a r s o n G. Methods for enhancing symbiotic nitrogen fixation. Plant and Soil,1993, 152: 1-17.
19. H u r e k T., H a n d l e y L.L., R e i n h o l d - H u r e k B. e.a. Azoarcus grass endophytes contribute fixed nitrogen to the plant in an unculturable state. Molec. Plant-Microbe Interact.,2002, 15: 233-242.
20. K a r g T., R e i n h o l d - H u r e k B. Global changes in protein composition of N2-fixing Azoarcus sp. strain BH72 upon diazosome formation. Journal of Bacteriology,1996, 178: 5748-5754.
21. W h i t e J.F., S u l l i v a n R.F., M o y M. e.a. Evolution of Epichloë/Neotyphodium endophytes and other Clavicipitacean biotrophs. In: Symbiosis: mechanisms and model systems /J. Seckbach (ed.). Dordrecht, Boston, L., 2002: 413-424.
22. W h i p p s J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exper. Bot., 2001, 52: 487-511.
23. R i l e y I.T., R e a r d o n T.B. Isolation and characterization of Clavibacter tritici associated with Anguina tritici in wheat from Western Australia. Plant Pathology, 1995, 44: 805-810.
24. M ü l l e r - S c h ä r e r H., S c h e e p e n s P.C. Biological control of weeds in crops: a coordinated European research program (COST-816). Inegr. Pest Manag. Rev., 1997, 2: 45-50.
25. C h a r u d a t t a n R., D i n o o r A. Biological control of weeds using plant pathogens: accomplishments and limitations. Crop Protection, 2000, 19: 691-695.
26. М е р е ж к о в с к и й С.С. Результаты полевых опытов истребления мышей бациллом, выделенным из сусликов. Архив ветеринарных наук, 1895: 1-23.
27. M o h a m m e d S.H., S e a d y M.A., E n a n M.R. e.a. Biocontrol efficiency of Bacillus thuringiensis toxins against root-knot nematode, Meloidogyne incognita. J. Cell. Mol. Biol., 2008, 7: 57-66.
28. R o m e i s J., M e i s s l e M., B i g l e r F. Transgenic crops expressing Bacillus thuringiensis toxins and biological control. Nat. Biotechnol., 2006, 24: 63-71.
29. С м и р н о в О.В., Г р и ш е ч к и н а С.Д. Изучение действия биопрепаратов на основе Bacillus thuringiensis на фитопатогенные грибы. Вестник защиты растений, 2010, 1: 27-35.
30. P u t t e r I., M a c C o n n e l l J.G., P r e i s e r F.A. e.a. Avermectins: novel insecticides, acaricides and nematicides from a soil microorganism. Experientia, 1981, 37: 963-964.
31. К а н д ы б и н Н.В. Биопестициды: теория и практика. Защита растений, 1991, 1: 5-6.
32. R y a n R.P., G e r m a i n e K., F r a n k s A. e.a. Bacterial endophytes: recent developments and applications. FEMS Microbiol. Lett., 2008, 278: 1-9.
33. L e d e r b e r g J., M c C r a y A.T. «Ome sweet» omics — a genealogical treasury of words. Scientist, 2001, 15: 8.
34. R o s e n b l u e t h M., M a r t i n e z - R o m e r o E. Bacterial endophytes and their interactions with hosts. Mol. Plant-Microbe Interact., 2006, 19: 827-837.
35. Y r j ä l ä K., M a n c a n o G., F o r t e l i u s C. e.a. The incidence of Burkholderia in epiphytic and endophytic bacterial cenoses in hybrid aspen grown on sandy peat. Boreal Environ. Res., 2010, 15: 81-96.
36. S a n c h e z - C o n t r e r a s M., B a u e r W.D., G a o M. e.a. Quorum-sensing regulation in rhizobia and its role in symbiotic interactions with legumes. Phil. Trans. R. Soc. B., 2007, 362: 1149-1163.
37. B r y a n J.A., B e r l y n G.P., G o r d o n J.C. Towards a new concept of the evolution of symbiotic nitrogen fixation in the Leguminosae. Plant and Soil,1996, 186: 151-159.
38. S p r e n t J.I. Evolving ideas of legume evolution and diversity: a taxonomic perspective on the ocurrence of nodulation. New Phytologist, 2007, 174: 11-25.
39. R e d m a n R.S., R a n s o n J.C., R o d r i g u e z R.J. Conversion of the pathogenic fungus, Colletotrichum magna to a nonpathogenic, endophytic mutualist by gene disruption. Mol. Plant-Microbe Interact., 1999, 12: 969-975.
40. S i k o r a R.A., P o c a s a n g e L., F e l d e A. e.a. Mutualistic endophytic fungi and in planta suppressiveness to plant parasitic nematodes. Biol. Control, 2008, 46: 15-23.
41. H o b s o n P.N., S t e w a r t C.S. The rumen microbial ecosystem. Berlin, 1997.
42. П р о в о р о в Н.А., Т и х о н о в и ч И.А. Эколого-генетические принципы селекции растений на повышение эффективности взаимодействия с микроорганизмами. С.-х. биол., 2003, 3: 11-25.
43. К р е т о в и ч В.Л. Биохимия усвоения азота воздуха растениями. М., 1994.
44. A n d r e w s M., L e a P.J., R a v e n J.A. e.a. Nitrogen use efficiency. 3. Nitrogen fixation: genes and costs. Ann. Appl. Biol., 2009, 155: 1-13.
45. П р о в о р о в Н.А., В о р о б ь е в Н.И. Роль горизонтального переноса генов в эволюции клубеньковых бактерий, направляемой растением-хозяином. Усп. совр. биол., 2010, 130: 336-345.
46. Биопрепараты в сельском хозяйстве /Под ред. И.А. Тихоновича, Ю.В. Круглова. М., 2005.
47. L e p p y a n e n I.V., O v t s y n a A.O., D o l g i k h E.A. e.a. New approach to the synthesis of chitin olygomers — elisitors of plant defense reactions. Heterologous expression of Mesorhizobium loti unique enzymes controlling this synthesis. Proc. 4th Baltic Sea Symposium «Novel technologies for management of beneficial and harmful microbes in the root system». Tune, 2008: 10.
48. Ш т а р к О.Ю., Д а н и л о в а Т.Н., Н а у м к и н а Т.С.и др. Анализ исходного материала гороха посевного (Pisumsativum L.) для селекции сортов с высоким симбиотическим потенциалом и выбор параметров для его оценки. Экологическая генетика, 2006, 4: 22-28.
49. B o r i s o v A.Y., D a n i l o v a T.N., S h t a r k O.Y. e.a. Tripartite symbiotic system of pea (Pisum sativum L.): applications in sustainable agriculture. In: Biological nitrogen fixation: towards poverty alleviation through sustainable agriculture /F.D. Dakora e.a. (eds.). Berlin, 2008: 15-17.
50. V a n d e r L e l i e D., T a g h a v i S., M o n c h y S. e.a. Poplar and its bacterial endophytes: coexistence and harmony. Crit. Rev. Plant Sci., 2009, 28: 346-358.
51. P o p o v a L.Yu., L o b o v a T.I., K r y l o v a T.Yu. e.a. Population dynamics of microorganisms in the different microecosystem conditions. Adv. Space Res., 2001, 27: 1571-1579.
52. К у п р и я н о в А.А., С е м е н о в А.М., В а н Б р у г г е н А.Х.К. Перемещение энтеропатогенных и сапротрофных бактерий в цикле экониш: животные—экскременты—почва—растения—животные. Изв. РАН, сер. биол., 2010, 3: 318-323.
AGRICULTURAL MICROBIOLOGY AS THE BASIS OF ECOLOGICALLY SUSTAINABLE AGRICULTURE: FUNDAMENTAL AND APPLIED ASPECTS
I.A. Tikhonovich, N.A. Provorov
Agricultural microbiology is based on the research of microbial communities living in the plants, animals and soils. Since the plants and animals create on their surfaces, in tissues and cells the variable niches for hosting the microbial partners, the genes controlling development of these niches are considered as the major determinants for the activities of agriculturally valuable microbes. Knowledge on their genetics, physiology, molecular biology and ecology enables us to construct the sustainable agricultural systems wherein: 1) productivity of plants and animals in increased by changing the composition and properties of the microbial symbionts; 2) ecologically dangerous agrochemicals (mineral fertilizers, pesticides) are completely or partially substituted by microbial preparations; 3) the energy consumption is decreased while the quality of agricultural products is improved.
Keywords: agricultural microbiology, microbe-plant interactions, N2-fixing symbioses, arbuscular mycorrhiza, endophytic microorganisms, rhizospheric growth-stimulating bacteria, biocontrol of pests and pathogens, microbiota of rumen, metagenomes of soil, ecologically sustainable soil tillage.
ГНУ Всероссийский НИИ сельскохозяйственной |
Поступила в редакцию |




