








УДК 633.31:581.557:631.461.5:575
ПОЛИМОРФИЗМ bet-ГЕНОВ У ШТАММОВ Sinorhizobium meliloti ИЗ ГЕНЦЕНТРОВ ЛЮЦЕРНЫ
М.Л. РУМЯНЦЕВА, В.С. БЕЛОВА, О.П. ОНИЩУК, Е.Е. АНДРОНОВ, О.Н. КУРЧАК, Е.П. ЧИЖЕВСКАЯ, Т.Б. РУМЯНЦЕВА, Б.В. СИМАРОВ
У 319 природных штаммов Sinorhizobium meliloti, выделенных из почв и клубеньков в 4 географически удаленных центрах разнообразия люцерны, изучали структурный полиморфизм хромосомного гена betC и плазмидного гена betT(S), которые ответственны за синтез и транспорт осмопротекторов. Показаны достоверные различия между клубеньковыми и почвенными субпопуляциями S. meliloti по частоте аллелей генов betC и betT(S). Выделены четыре преобладающих геномотипических bet-класса, частота которых в клубеньковых и почвенных субпопуляциях достоверно не различалась. Выявлен высокий уровень корреляции между солеустойчивостью и симбиотической эффективностью ризобий (r = 0,76). Полученные результаты указывают, что полиморфизм bet-генов у ризобий Северокавказского генцентра обусловлен преимущественно влиянием растения-хозяина, тогда как в Приаральском, подверженном засолению, — процессами адаптации к воздействию абиотического стресс-фактора.
Ключевые слова: клубеньковые бактерии, субпопуляции, bet-гены, полиморфизм, солеустойчивость, симбиоз, люцерна, генцентры.
Клубеньковые бактерии (ризобии) Sinorhizobium meliloti фиксируют азот атмосферы в симбиозе с люцерной, широко выращиваемой во всем мире. Люцерна — умеренный галофит, и это обусловливает перспективность ее возделывания на деградированных (например, засоленных) почвах (1, 2). Солеустойчивость люцерны повышается при инокуляции штаммами-микросимбионтами, подобранными в результате селекции (3). Природные изоляты S. meliloti существенно различаются по солеустойчивости, чем объясняется интерес к изучению генетического контроля этого признака.
Известно, что солеустойчивость ризобий зависит от их способности накапливать легкорастворимые соединения-осмопротекторы, из которых у многих видов микроорганизмов и растений наиболее распространен глицин-бетаин (4-6). Клубеньковые бактерии люцерны увеличивают внутриклеточную концентрацию глицин-бетаина за счет синтеза или экзогенного транспорта либо двух процессов одновременно (5). Ризобии вида S. meliloti используют глицин-бетаин в качестве источника углерода и азота, что не показано для других видов бактерий (5, 7). Биосинтез глицин-бетаина у S. meliloti контролируется bet-опероном, который расположен на хромосоме и выявлен у ряда солеустойчивых видов бактерий, а также у Escherichia сoli(8). В состав оперона входит регуляторный ген betI, ген betB (NAD-зависимая бетаинальдегиддегидрогеназа) и ген betА (кислородзависимая холиндегидрогеназа). В отличие от ряда солеустойчивых бактерийу S. melilotibet-оперонсодержит еще один, четвертый, ген — betC, кодирующий холинсульфатазу, ответственную за превращение холин-О-сульфата в холин (9). Накопление глицин-бетаина у клубеньковых бактерий люцерны происходит при участии продукта гена betS — мембранного белка BetS, относящегося к транспортной системе BCCT-типа (betain-cholin-carnitin transport), обеспечивающей накопление бетаина, холина, карнитина (10). Это единственная система из описанных в литературе, которая ответственна за экспресс-ответ ризобий на солевой шок (мгновенное увеличение концентрации соли в среде) (11). Ген betS впервые выявлен и изучен у штамма S. meliloti 102F34 и аннотирован в геноме штамма Rm1021 под номером SMb20333 как ген betT (10). Ген betT(S) локализован на высокомолекулярной симбиотической мегаплазмиде SMb (12). В клетках S. meliloti этот ген экспрессируется конститутивно, однако при выращивании в присутствии NaCl продукт гена — белок BetS претерпевает посттрансляционную модификацию. По мнению авторов, это позволяет бактериям быстро транспортировать глицин-бетаин через мембрану из ризосферы или из почвы (10). Показано, что ген betS экспрессируется в инфекционных нитях и в бактероидах, что свидетельствует о важной роли мембранного белка BetS при формировании и функционировании симбиоза в условиях засоления (11). К гену betT(S) прилегают открытые рамки SMb20332 и SMb20334, функциональная значимость которых не установлена. Применение программы BLAST-N позволило установить, что последовательность SMb20332 гомологична последовательности гена, кодирующего арилсульфатазу (100 %), SMb20334 — последовательности гена, кодирующего неспецифичную серин/треонин киназу. Изучение генетических основ формирования солеустойчивости предполагает анализ структурного полиморфизма bet-генов, участвующих в синтезе и транспорте осмопротекторов, у природных штаммов клубеньковых бактерий в различных агроклиматических и географически удаленных регионах.
В связи с этим мы проанализировали структурный полиморфизм хромосомного гена betC и плазмидного гена betT(S) у природных штаммов Sinorhizobiummeliloti, адаптированных к различным абиотическим стрессорам, во взаимосвязи с солеустойчивостью и симбиотической активностью в условиях засоления.
Методика. Коллекция природных штаммов S. melilotiсобрана в очагах разнообразия бобовых растений-хозяев, относящихся к Кавказскому, Среднеазиатскому центрам разнообразия культурных растений (описаны Н.И. Вавиловым) (13), а также в Европейско-Сибирском генцентре (14, 15) и в центре современной интрогрессивной гибридизации люцерны (предгорье Мугоджар, Приаралье, Казахстан) (16). Почвенные образцы и части корней с клубеньками отбирали в различных сайтах генцентров (ГЦ), которые далее обозначены как Приаральский (ПАГ), Северо-Кавказский (СКГ), Европейско-Сибирский (ЕСГ) и Среднеазиатский (САГ) (3, 17). Перечисленные ГЦ географически удалены, подвержены резким перепадам атмосферных и почвенных температур, испытывают недостаток влаги. В Приаралье происходит интенсивное вторичное засоление (17). Популяции изолятов ризобий состоят из почвенных (П) и клубеньковых (К) субпопуляций (17). Штаммы бактерий из клубеньков или из почвенных вытяжек выделяли по стандартным методикам (3). Всего проанализировано 167 изолятов S. meliloti из ПАГ, 97 — из СКГ, 30 — из ЕСГ и 25 — из САГ.
Солеустойчивостьштаммов определяли на основании измерения оптической плотности (OD600) бактериальных культур после 4 сут роста в жидкой среде TY с 0,6 М NaCl (3,5 %) (3). По результатам роста изоляты S. meliloti разделяли на солеустойчивые и солечувствительные (3).
Режим ПЦР включал 30 циклов. Продолжительность денатурации ДНК в первом цикле составляла 3 мин. Последующие циклы включали денатурацию (30 с при 94 °С), отжиг (30 с при 55 °С) и элонгацию (1 мин при 72 °С).
При изучении последовательности гена betC использовали праймеры, с помощью которых амплифицировали последовательность гена betC, межгенную область и часть гена betB: betCB_F — 3´-AACTACACCTCGTCGCCCCTGTGTG-5´ иbetCB_R — 3´-TCT-CGACGATCCCAGGCGTT-5´. Локус betТ(S) изучали с тремя парами праймеров — на последовательности SMb20332 (BetS1_F: 3´-GCTTCTTCTTCCTGCCATCA-5´ и BetS1_R 3´-CACCTTCTTTTCGGACGGTA-5´), SMb20333 (betS) (BetS2_F: 3´-TACCGTCCGAAAAGAA-GGTG-5´, BetS2_R: 3´-CGATGCTATTTCTGGCGTTC-5´) и SMb20334 (BetS3_F: 3´-GAT-TGTAGCGGTAGCCGAAA-5´). Рестрикцию амплифицированных ДНК-фрагментов эндонуклеазами MspI, HaeIII, AluI («MBI Fermentas», Литва) выполняли по стандартной методике (18), электрофоретический анализ рестриктов проводили в 3 % агарозном геле (18). В результате для каждого штамма выявили присущий ему состав ДНК-фрагментов определенного размера — так называемый RFLP-тип. Каждому RFLP-типу присваивалось буквенное обозначение (сходные составы фрагментов ДНК относили к одному RFLP-типу). RFLP-тип а был всегда аналогичен RFLP-типу типового штамма Rm1021 S. meliloti (19). Если анализ штамма проводили с применением двух и более эндонуклеаз, его RFLP-тип описывали как комплексный (по буквенным обозначениям RFLP-типов для каждой эндонуклеазы).
ДНК-ДНК гибридизацию выполняли по методу Саузерна (18). Гибридизационными пробами служили DIG-меченные амплифицированные фрагменты ДНК, содержавшие последовательности исследуемых генов.
Симбиотическую эффективность штаммов S. meliloti оценивали в условиях стерильных микровегетационных опытов по изменению сухой массы растений люцерны Medicagosativa L. (сорт Вега) или M. truncatula (сорт Jemalong) относительно варианта без инокуляции (3). Эффективными считались штаммы, при инокуляции которыми сухая масса у инокулированных растений на 75 % превышала таковую у растений в контроле (без инокуляции).
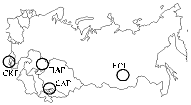 |
Рис. 1. Генцентры культурных растений, в которых проводился сбор образцов почвы и корней люцерны с клубеньками для выделения природных штаммов бактерий Sinorhizobiummeliloti: СКГ — Северо-Кавказский, ПАГ — Приаральский, САГ — Среднеазиатский, ЕСГ — Европейско-Сибирский. |
Статистическую обработку проводили с использованием программ Statistica 6.0 (корреляционный анализ), Microsoft Excel 2000 (c2; ошибка доли, %) и коэффициента гетерогенности Н (20).
Результаты. Расположение центров, в которых проводили сборы почвенных образцов и корней люцерны с клубеньками, представлено на рисунке 1.
С т р у к т у р н ы й п о л и м о р ф и з м
х р о м о с о м н о г о г е н а betC в п о п у л я ц и я х
и с у б п о п у л я ц и я х р и з о б и й.П о п у л я ц и и. RFLP-полиморфизм гена betC (1401 п.н.) и части гена betB (143 п.н.; рис. 2) изучили у 319 природных штаммов ризобийиз четырех ГЦ. При использовании эндонуклеазы MspI выявлено три RFLР-типа (а, b и c), при использовании HaeIII — два (а, b) (см. рис. 2, а, в). RFLP-типы, отличавшиеся от типа а, присущего типовому штамму S. meliloti Rm1021, имели либо дополнительные MspI сайты и/или меньшее число HaeIII сайтов преимущественно в центральной части гена betC между 691 по 1120 нуклеотидами (см. рис. 2). При учете двух рестрикционных профилей, полученных для каждого изолята, определили пять комбинированныхMspI-HaeIII RFLP-типов — aa, ab, ba, bb и cb, которые рассматривали как аллели гена betC.
А |
Б |
|
|
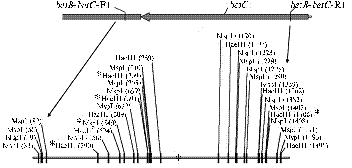
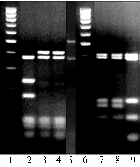
Рис. 2. Рестрикционная карта гена betC штамма Rm1021Sinorhizobiummeliloti (А, звездочками отмечены сайты, в которых при рестрикционном анализе выявили структурные различия) и RFLP-типы (Б) природных изолятов S.meliloti: RFLP-типы (a, b, c), полученные при обработке ПЦР-ДНК-фрагментов эндонуклеазой MspI (2, 3, 4, 5) или HaeIII (7, 8, 9); 1, 6 — ДНК-маркер (М100) (описание см. в разделе «Методика»). |
|
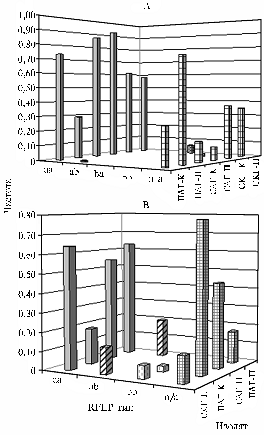
Аллели аа и ab гена betC выявили у всех изолятов ПАГ, САГ и ЕСГ, при этом аллель аа доминировал по частоте (средние значения для аллелей составили соответственно 0,61 и 0,32; рис. 3, А). Только у изолятов СКГ по обоим аллелям частота была сходной — соответственно 0,37 и 0,49. Остальные аллели (ba, bb, cb) идентифицировали у 1-3 изолятов САГ, ПАГ и СКГ и не выявили у изолятов ЕСГ. Частота этих аллелей составила от 0,02 до 0,12 в зависимости от ГЦ, в связи с чем их рассматривали как редко встречающиеся или уникальные. Особо следует отметить, что последовательность гена betC не амплифицировалась с праймерами, подобранными на основе данных сиквенса генома Rm1021, у 9 изолятов СКГ, 4 изолятов ПАГ и у 1 изолята СКГ. У ряда природных штаммов последовательности генов betC или betТ(S) (см. далее) не удалось амплифицировать с использо-
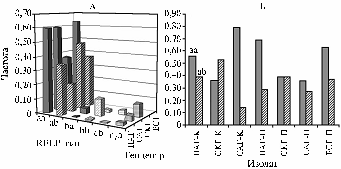 |
Рис. 3. Частота RFLP-типов betC у изолятов Sinorhizobiummeliloti из различных генцентров: А — частота встречаемости всех выявленных RFLP-типов в популяциях, Б — частота встречаемости RFLP-типов aa и ab в субпопуляциях; aa, ab, ba, bb, cb — RFLP-типы гена betC; n/a — отсутствие амплификации гена betC; ПАГ, САГ, СКГ, ЕСГ — генцентры (описание см. в разделе «Методика»); К — клубеньковые, П — почвенные изоляты. |
ванием праймеров, подобранных подобным образом, причем гибридизационный анализ общей ДНК таких изолятов с ДНК-пробами на соответствующие гены Rm1021 для части изолятов дал положительный ответ, который у других не проявился. Отсутствие амплификата у изолятов с положительным гибридизационным ответом свидетельствовало в поль-зу того, что последовательность изучаемого гена структурно изменена (дивергентна). Если у штамма не получен амплификат и положительный ответ при ДНК-ДНК гибридизации, принято считать, что соответствующий ген отсутствует в геноме изучаемого(ых) природного(ых) штамма(ов) либо его последовательность дивергентна (21).
Таким образом, аллель аа, характерный для типового штамма Rm1021, доминирует в очагах разнообразия культурных растений ПАГ и САГ, расположенных в Средней Азии, и ЕСГ (вторичный центр разнообразия).
К л у б е н ь к о в ы е и п о ч в е н н ы е с у б п о п у л я ц и и. К- и П-субпопуляции ризобий из изучаемых ГЦ также были проанализированы по частотам пяти аллелей гена betC. Аллели аа и аb преобладали, поскольку их выявили с частотой соответственно 0,51-0,58 и 0,42-0,33 у К- и П-изолятов (см. рис. 3, Б). Остальные типы аллелей гена betC идентифицированы у 1-3 П- или К-изолятов. Вместе с тем при сравнении изолятов из К-субпопуляций ПАГ и СКГ обнаружилось, что частоты аллелей аа и аb достоверно различались (χ2 = 4,6, Р < 0,05; см. рис. 3, Б). Аллель аа преимущественно находили у К-изолятов ПАГ (0,53), аллель аb — у К-изолятов СКГ (0,52; см. рис. 3, Б). Среди К-изолятов САГ доминантой оказался аллель аа (0,78). Таким образом, этот аллель преобладал в К-субпопуляциях ПАГ и САГ, относящихся к Среднеазиатскому региону; у К-изолятов СКГ преобладал аллель аb. Анализ П-субпопуляций показал, что аллель аа гена betC превалировал в ПАГ и ЕСГ (соответственно 0,70 и 0,63; см. рис. 3, Б). Частота аллеля аb в П-субпопуляциях всех изучаемых ГЦ была сходной и составила в среднем 0,33. В П-субпопуляции СКГ аллели аа и ab встречались с равной частотой (0,39), в П-субпопуляции САГ — выявлялись с частотой соответственно 0,36 и 0,27. Следовательно, аллель аа доминировал в трех из четырех исследованных П-субпопуляций клубеньковых бактерий люцерны S. meliloti (χ2 = 11,8, Р < 0,001).
Поскольку аллели аа и ab гена betC обнаружили у клубеньковых и почвенных изолятов из различных ГЦ, их распределение не зависело от географического происхождения штаммов. Вместе с тем преобладание штаммов, содержащих, например, аллель ab в К-субпопуляции СКГ и аллеля аа у К- и П-изолятов ПАГ, могло быть обусловлено селективным влиянием со стороны растения-хозяина и/или адаптацией к абиотическим стресс-факторам.
Анализ встречаемости изолятов, у которых ген betC не был амплифицирован, не выявил достоверных различий между К- и П-изолятами в СКГ и ПАГ (c2 = 0,36, Р > 0,1). Следовательно, изоляты, у которых ген betC отсутствует или последовательность гена дивергентна, встречаются с равной вероятностью в клубеньковой и почвенной субпопуляциях.
При определении коэффициентов Н на основании частоты рассмотренных RFLP-типов betC (20) установили, что у изолятов САГ, ПАГ и СКГ гетерогенность была сходно высокой (Н соответственно 0,63; 0,60 и 0,65), тогда как для ЕСГ значение коэффициента было ниже (Н = 0,5). При сравнении субпопуляций оказалось, что П-изоляты САГ и СКГ гетерогеннее К-изолятов из тех же ГЦ (соответственно 0,72 и 0,70; 0,55 и 0,61), тогда как в зоне экстремального засоления (ПАГ) у К-изолятов этот показатель был выше, чем у П-изолятов (соответственно 0,64 и 0,56).
|
|
Рис. 4. Структурный полиморфизм локуса SMb20332-SMb20334 у клубеньковых (К) и почвенных (П) изолятов Sinorhizobiummeliloti из различных генцентров: А — SMb20333, Б — SMb20332, В — SMb20334; aa, ba, bb, ab— RFLP-типы соответствующего гена; n/a — отсутствие амплификации; i-xi — комбинированные MspI-HaeIII RFLP-типы SMb20332; ПАГ, САГ, СКГ — генцентры (описание см. раздел «Методика»). С т р у к т у р н ы й п о л и- |
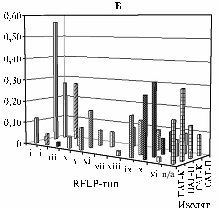
4, А). Дополнительное использование в RFLP-анализе мелкощепящей эндонуклеазы AluI позволило выделить у MspI-HaeIII RFLP-типа аа два подтипа. Первый из них был также аналогичен RFLP-типу Rm1021, второй идентифицировали у двух изолятов СКГ (0,11), тогда как у изолятов из других ГЦ полиморфизм последовательности гена betТ(S) при использовании эндонуклеазы AluI не выявлен.
Изоляты, не содержавшие последовательность SMb20333 или имевшие дивергентную структуру этого гена (возможное обоснование причин обсуждалось выше), обнаружились во всех изучаемых ГЦ. У К- и П-субпопуляций СКГ частоты были сходными и составили в среднем соответственно 0,37. У К- и П-субпопуляций САГ частоты также были сходными и составили в среднем 0,12. Однако в районе экстремального засоления (ПАГ) доля таких изолятов достоверно превалировала в П-субпопуляции (χ2 = 8,4; Р < 0,01; см. рис. 4, А). При ДНК-ДНК гибридизационном анализе 13 К- и П-изолятов СКГ и ПАГ, у которых при использовании праймеров на ген betТ(S)амплификация не происходила, были получены результаты, позволяющие считать, что у 4 из них последовательность гена betТ(S) дивергентна, а в геномах остальных 9 она, по-видимому, отсутствует. Изоляты, у которых последовательность гена betТ(S) дивергентна, оказались солечувствительными, не имевшие его — проявляли различную степень солеустойчивости. Полученные данные подтверждают мнение о том, что в транспорте осмопротекторов у S. meliloti могут, помимо betТ(S), участвовать иные, не известные в настоящее время гены (10).
ПоследовательностьSMb20332, прилегающая к гену betT(S), наиболее полно изучали у изолятов САГ и ПАГ. При использовании рестриктазы MspI выявили семь RFLP-типов, причем те из них, которые отличались от типа а (Rm1021), имели на 1-7 сайтов рестрикции меньше (исключение — тип d, у которого обнаружен новый дополнительный сайт; данные не приведены). Четыре из семи MspI RFLP-типов (a, c, f, g) были общими для изолятов САГ и ПАГ, три остальных имелись у изолятов либо из ПАГ (e), либо из САГ (b, d). При использовании эндонуклеазы HaeIII также идентифицировали семь RFLP-типов, из которых шесть (a, b, d, h, g и e) найдены в САГ и ПАГ и один (уникальный c-тип) — у изолятов из САГ. Число комбинированных MspI-HaeIII RFLP-типов SMb20332 составило 11 (далее обозначали как i-xi, RFLP-типу штамма Rm1021 соответствует тип i) (см. рис. 4, Б). При анализе К-субпопуляций САГ и ПАГ идентифицировали соответственно 5 и 9 из 11 этих MspI-HaeIII типов (см. рис. 4, Б). Четыре RFLP-типа (i, v, x и xi) преимущественно встречались у К-изолятов ПАГ (соответственно 0,12; 0,1; 0,18 и 0,27), тип i — у К-изолятов САГ (0,57). При анализе П-субпопуляций ПАГ и САГ выявлено соответственно 5 и 6 из 11 идентифицированных MspI-HaeIII RFLP-типов. Среди П-изолятов из ПАГ преобладали типы v, vii и ix (0,17, 0,15 и 0,32), из САГ — i и ii (по 0,27 для каждого).
Общими в К- и П-субпопуляциях были четыре RFLP-типа — i, v, vii и ix. Последний из перечисленных выявили у значительной части К- и П-изолятов ПАГ (частоты соответственно 0,27 и 0,32), в субпопуляциях САГ — только у единичных образцов. Типы v и vii имелись преимущественно у К- и П-изолятов ПАГ (см. выше), тогда как в субпопуляциях САГ их идентифицировали только дважды. Тип i (Rm1021) обнаружили с частотой соответственно 0,12 и 0,57 у К-изолятов из ПАГ и САГ и у единичных П-изолятов из обоих ГЦ. Таким образом, RFLP-тип, характерный для Rm1021, выявлен преимущественно в К-субпопуляции САГ (различия недостоверны;χ2= 0,6; P > 01). Остальные 4 из 11 MspI-HaeIII RFLP-типов, были идентифицированы у единичных К- и П-изолятов из ПАГ и САГ (средняя частота соответственно 0,02 и 0,08).
Последовательность SMb20332 отсутствовала или была дивергентной с частотой соответственно 0,12 и 0,23 у изолятов из САГ и ПАГ. Анализ 9 таких изолятов показал, что у 3 из них указанная последовательность дивергентна, а у 6 отсутствует. Изоляты, содержавшие дивергентную последовательность SMb20332, также имели дивергентную структуру гена bet(Т)S и были солечувствительными, то есть у них нарушался механизм формирования устойчивости к стресс-фактору.
Последовательность SMb20334, также прилегающую к гену bet(Т)S, изучали у 25 изолятов САГ и 66 изолятов ПАГ (см. рис. 4, В). Аллель аа доминировал у К- и П-изолятов САГ и К-изолятов ПАГ (средняя частота по всем трем субпопуляциям 0,61), у П-изолятов ПАГ был выявлен в 3 раза реже (0,19). Кроме того, у единичных К-изолятов САГ и ПАГ имелись аллели ab и bb. Последовательность SMb20334 отсутствовала или была дивергентна преимущественно у К-изолятов ПАГ (0,78) (у П-изолятов из этого же региона — 0,14). В то же время в САГ аналогичные изоляты идентифицировали в основном среди П- (0,45), но не К-изолятов (0,17). То есть в районе экстремального засоления П-изоляты преимущественно наследовали аллель аа, характерный для Rm1021, а в К-субпопуляции достоверно чаще встречались изоляты, не содержавшие последовательность SMb20334 (χ2= 19,5; Р < 0,001).
Коэффициент гетерогенности (Н) определили для всех трех рамок считывания (SMb20332-SMb20334). Высокий полиморфизм гена betТ(S) (SMb20333) выявили у К-изо-лятов ПАГ (0,63), тогда как у П-изолятов ПАГ и К- и П-изолятов САГ последовательность гена консервативна (соответственно 0,30; 0,38 и 0,28). Коэффициент гетерогенности, определенный для SMb20332, указывал на высокий полиморфизм этой последовательности у К- и П-изолятов САГ (соответственно 0,81 и 0,85) и ПАГ (0,70 и 0,77). Вторая прилегающая последовательность SMb20334 у К-изолятов САГ была гетерогеннее, чем у П-изо-лятов из того же региона (соответственно 0,70 и 0,51), тогда как в районе экстремального засоления (ПАГ), наоборот, последовательность оказалась более гетерогенной у П-изо-лятов, чем у К-изолятов (соответственно 0,66 и 0,47).
Таким образом, высокий полиморфизм характерен для последовательности гена betT(S) у К-изолятов и последовательности SMb20334 у П-изолятов ПАГ. У К- и П-изолятов САГ выявлен низкий полиморфизм гена betT(S) и высокий — SMb20334. Исключение составила последовательность SMb20332, для которой полиморфизм оказался высоким у всех изолятов ПАГ и САГ.
Частота изолятов S.meliloti, у которых все три последовательности локуса SMb20332-SMb20334 дивергентны, составила 0,13. Изоляты, у которых отсутствовала последовательность SMb20332 или SMb20333, встречались с равной частотой (0,30), не имеющие SMb20334 — значительно реже (0,09). Эти данные указывают, что последовательности SMb20332 и SMb20333 утрачивались в 3 раза чаще, чем SMb20334. Преимущественная делеция участка SMb20332-SMb20333 второй симбиотической мегаплазмиды S. melilotiмогла быть обусловлена присутствием элемента IS17 в межгенной области, расположенной перед SMb20332 (22).
Г е н о м о т и п и ч е с к и е к л а с с ы betC/betT(S)п р и р о д н ы х
ш т а м
м о в р и з о б и й. К- и П-изоляты ПАГ, САГ и СКГ объединили и распределили по генотипическим классам согласно RFLP-типам генов betC и betT(S) (табл.). Из 12 выявленных классов 4 были доминирующими: к ним относились 232 из 319 изученных образцов (72 %). Частота генотипа А (присущ Rm1021) и В была сходной для К- и П-изолятов, однако и тот и другой генотип встречался у первых в 1,6 раза чаще, чем у вторых (см. табл.). Интересно, что у П-изолятов, наследовавших аллель аа гена betC, достоверно чаще наблюдалось отсутствие гена betT(S) (χ2 = 6,99; Р < 0,01). При сравнении К- и П-субпопуляций из разных очагов разнообразия растений-хозяев оказалось, что изоляты с генотипом А достоверно чаще встречались в К-субпопуляции ПАГ (χ2= 3,3; Р < 0,1; данные не представлены), тогда как среди К- и П-изолятов из СКГ и САГ преимущественного генотипа не выявили.
| Распределение генотипов у природных штаммов Sinorhizobiummeliloti по структурной организации betCи betT(S) генов, солеустойчивости и симбиотической активности в субпопуляциях клубеньковых (К) и почвенных (П) изолятов из ПАГ, СКГ и САГ | ||||||||
bet-Генотип |
bet-RFLP-тип |
Частота генотипа |
Доля, % |
|||||
солеустойчивых |
симбиотически |
|||||||
betC |
betT(S) |
К |
П |
К |
П |
К |
П |
|
A1 |
aa |
aa |
0,39 |
0,25 |
0,40 |
0,34 |
0,48*** |
0,17 |
B |
ab |
aa |
0,35 |
0,22 |
0,42 |
0,34 |
0,24 |
0,44 |
С |
аа |
н/а |
0,12 |
0,40** |
0,14 |
0,23 |
0,16 |
0,11 |
D |
ab |
н/а |
0,14 |
0,13 |
0,05 |
0,09 |
0,13 |
0,28 |
Всего изолятов |
147 |
85 |
88 |
35 |
147 |
85 |
||
Доля изолятов, % (Х±х) |
100 |
100 |
59,9±4,0 |
41,2±5,3 |
42,8±4,1 |
21,7±4,7 |
||
П р и м е ч а н и е. 1 — генотип, сходный с таковым у S. meliloti Rm1021; н/а — нет амплификата гена betT(S); ПАГ, СКГ и САГ — генцентры (описание см. в разделе «Методика»). |
||||||||
С о л е у с т о й ч и в о с т ь и с и м б и о т и ч е с к и е с в о й с т в а
и з о л я
т о в р и з о б и й с р а з л и ч н о й с т р у к т у р н о й о р г а н и з а ц и е й г е н о в betC и betT(S). Доля солеустойчивых среди К- и П-изолятов составила соответственно 59,9 и 41,2 % (см. табл.), однако эти различия недостоверны (χ2= 2,4; Р > 0,01), следовательно, выявление солеустойчивых изолятов в К- и П-субпопуляциях равновероятно. Солеустойчивые К- и П-изоляты преимущественно имели генотипы А и В, однако достоверных различий по их частоте между анализируемыми группами не обнаружили (χ2= 0,014; Р > 0,1). У солеустойчивых К- и П-изолятов также не наблюдалась корреляция между наличием аллеля аа или abгена betC и отсутствием гена betT(S). Симбиотически эффективными были преимущественно солеустойчивые К-изоляты с генотипом А и П-изоляты с генотипом В (χ2= 5,7; Р < 0,02). Солечувствительные изоляты достоверно чаще встречались в П-субпопуляции ПАГ (χ2= 9,25; Р < 0,01; данные не приведены). Оказалось, что солечувствительные П-изоляты преимущественно содержали аллель аа гена betC и не содержали гена betT(S) (χ2= 12,24; Р < 0,001) либо такие П-изо-ляты наследовали аллели генов betC и betT(S), отличные от описанных для типового штамма Rm1021. Солечувствительные изоляты были преимущественно симбиотически неэффективными. Следовательно, отсутствие гена betT(S) или структурные изменения в последовательности гена приводили у ризобий к нарушениям в формировании устойчивости в ответ на воздействие солевого стресса.
Установлено, что эффективный симбиоз с люцерной посевной (Medicagosativa) формировали около 42,8 % К- и не более 21,7 % П-изолятов. К-изоляты, преимущественно наследовавшие аллели генов betC и betT(S), характерные для Rm1021, образовывали эффективный симбиоз с M. sativa и M. truncatula (соответственно χ2= 5,35 и χ2=4,028; Р < 0,05). Доля симбиотически неэффективных образцов была выше среди П-изолятов (χ2 = 10,29; P < 0,01). Достоверно чаще изоляты, несущие аллель ab гена betC, формировали неэффективный симбиоз либо c M. sativa (П-изоляты, χ2= 11,97; P < 0,001), либо с M. truncatula(К- и П-изоляты, соответственно χ2= 12,1 при P <0,001 и χ2= 4,4 при P < 0,05).
Корреляция между солеустойчивостью и симбиотической эффективностью была выявлена только для К-изолятов (r = 0,76), у П-изолятов ее не обнаружили (r = 0,16).
Итак, впервые изучен полиморфизм bet-генов, вовлеченных в синтез и транспорт осмопротекторов, в природных популяциях штаммов Sinorhizobiummeliloti в очагах разнообразия люцерны, относящихся к четырем различным генцентрам культурных растений. Выявлены существенные различия между клубеньковыми и почвенными субпопуляциями ризобий люцерны по частоте аллелей bet-генов, локализованных на хромосоме и мегаплазмиде. В то же время по частоте bet-генотипов достоверных различий между субпопуляциями не обнаружено. Впервые изучена взаимосвязь между bet-генотипом природных штаммов, их солеустойчивостью и симбиотической эффективностью. Показано, что изоляты с bet-генотипом, аналогичным таковому у типового штамма Rm1021, преобладали как в клубеньковой, так и в почвенной субпопуляциях ризобий, были солеустойчивыми и формировали эффективный симбиоз с люцерной посевнойMedicagosativa и люцерной однолетней M. truncatula. Изоляты S. meliloti, наследовавшие иные, чем у типового штамма Rm1021, аллели хромосомного гена betС, проявляли сниженную симбиотическую эффективность, а имеющие дивергентную последовательность гена betT(S) или не содержавшие этот ген обладали измененной солеустойчивостью. Особо следует отметить, что самый высокий уровень полиморфизма выявлен в последовательностях bet-генов, локализованных на хромосоме и плазмиде, у изолятов СКГ (Северо-Кавказский генцентр). Географический район, в котором собирали образцы, представляет собой часть первичного центра разнообразия диплоидных форм люцерны, и здесь влияние растения-хозяина на генетическую стабильность микросимбионтов может иметь большее значение, чем эффект абиотических стресс-факторов. В пользу этого свидетельствует тот факт, что изменения обнаружены преимущественно у изолятов, выделенных из клубеньков. В районе, испытывающем вторичное засоление (Приаралье), главную роль, очевидно, играют процессы адаптации к стресс-фактору (засолению), что подтверждается выявлением структурных перестроек в bet-генах преимущественно у почвенных изолятов S. meliloti.
Л И Т Е Р А Т У Р А
1. Ш а м с у т д и н ов З.Ш., С а в ч е н к о И.В., Ш а м с у т д и н о в Н.З. Галофиты России, их экологическая оценка и использование. М., 2000.
2. Z a h r a n H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev., 1999, 63: 968-989.
3. И б р а г и м о в а М.В., Р у м я н ц е в а М.Л., О н и щ у к О.П. и др. Симбиоз клубеньковых бактерий Sinorhizobiummelilotiс люцерной Medicagosativa в условиях засоления. Микробиология, 2006, 75: 94-100.
4. M i l l e r K.J., W o o d J.M. Osmoadaptation by rhizosphere bacteria. Review. Annu. Rev. Microbiol., 1996, 50: 101-136.
5. B o n c o m p a g n i E., О s t e r a s M., P o g g i M.C., L e R u d u l i e r D. Occurrence of choline and glycine betaine uptake and metabolism in the family Rhizobiaceae and their roles in osmoprotection. Appl. Environ. Microbiol., 1999, 65: 2072-2077.
6. H i n c h a D.K., H a g e m a n n M. Stabilization of model membranes during drying by compatible solutes involved in the stress tolerance of plants and microorganisms. Biochem. J., 2004, 383: 277-283.
7. S m i t h L.T., P o c a r d I.A., B e r n a r d T., L e R u d d i e r D. Osmotic control of glycine betaine biosynthesis and degradation in Rhizobium meliloti. J. Bacteriol., 1988, 170: 3142-3149.
8. L a m a r k T., K a a s e n I., E s h o o M. W., F a l k e n b e r g P., M c D o u g a l l J., S t r o m A.R. DNA sequence and analysis of the bet genes encoding the osmoregulatory choline-glycine betaine pathway of Escherichia coli. J. Mol. Microbiol., 1991, 5: 1049-1064.
9. O s t e r a s M., B o n c o m p a g n i E., V i n c e n t N. e.a. Presence of a gene encoding choline sulfatase in Sinorhizobium meliloti bet operon: choline-O-sulfate is metabolized into glycine betaine. PNAS, 1998, 95: 11394-11399.
10. B o s c a r i A., M a n d o n K., D u p o n t L. e.a. BetS is a major glycine betaine/proline betaine transporter required for early osmotic adjustment in Sinorhizobium meliloti. J. Bacteriol., 2002, 184: 2654-2663.
11. B o s c a r i A., V a n d e S y p e G., L e R u d u l i e r D., M a n d o n K. Overexpression of BetS, a Sinorhizobium meliloti high-affinity betaine transporter, in bacteroids from Medicago sativa nodules sustains nitrogen fixation during early salt stress adaptation. MPMI, 2006, 19: 896-903.
12. G a l i b e r t F., F i n a n T.M., L o n g S.R. e.a. The composite genome of the legume symbiont Sinorhizobium meliloti. Science, 2001, 293: 668-672.
13. В а в и л о в Н.И. Центры происхождения культурных растений. Тр. по прикл. бот. и сел., 1926, 16(2): 3-138.
14. Ж у к о в с к и й П.М. Мировой генофонд растений для селекции. В кн.: Вавилов и сельскохозяйственная наука. М., 1971: 120-202.
15. К о р о в и н а О.Н. Н.И. Вавилов о первичных центрах происхождения культурных растений. Ботанический журнал, 1987, 6: 721-731.
16. И в а н о в А.И. Люцерна. М., 1980.
17. Р у м я н ц е в а М.Л. Генетические ресурсы клубеньковых бактерий (обзор). Генетика, 2009, 45: 1-15.
18. М а н и а т и с Т., Ф р и ч Э., С э м б р у к Дж. Методы генной инженерии. Молекулярное клонирование. М., 1984.
19. R o u m i a n t s e v a M.L., A n d r o n o v E.E., S h a r y p o v a L.A. e.a. Diversity of Sinorhizobium meliloti from the Central Asian alfalfa gene center. Appl. Environ. Microbiol., 2002, 68: 4694-4697.
20. N e i M. Estimation of average heterozygosity and genetic distance from a small sample of individuals. Genetics, 1978, 89: 583-590.
21. G i u n t i n i E., Me n g o n i A., D e F i l i p p o C. e.a. Large-scale genetic variation of the symbiosis-required megaplasmid pSymA revealed by comparative genomic analysis of Sinorhizobium meliloti natural strains. BMC Genomics, 2005, 158, doi:10.1186/147-2164-6-158.
22. P i t s i k E.V., A n d r o n o v E.E., R o u m i a n t s e v a M.L., S i m a r o v B.V. The investigation of the betS gene polymorphism of the SymB plasmid in nodule bacteria Sinorhizobium meliloti. Abstr. of Postgraduate Course «Applied and fundamental aspects of responses, signaling and developmental process in the root-microbe systems» and Meeting of the Research Consortium on Evolution of Plant-Microbe Interactions. St.-Petersburg, 2007: 42.
POLYMORPHISM OF bet-GENES AMONG Sinorhizobium meliloti ISOLATES NATIVE TO GENE CENTERS OF ALFALFA
M.L. Roumiantseva, V.S. Belova, O.P. Onishchouk, E.E. Andronov, O.N. Kurchak, E.P. Chizhevskaya, T.B. Rumyantseva, B.V. Simarov
Polymorphism of chromosomal gene betC and plasmid localized loci betT(S) involved in osmoprotectants synthesis and transport was studied among 319 native isolates of Sinorhizobium meliloti, recovered from nodules and soil samples at four geographically distant gene centers of alfalfa. The significant difference between nodule and soil groups of isolates was shown by allelic rate of betC and betT(S) genes. Four dominant genotyping bet-classes among native isolates have similar popularity among nodule and soil groups of S. meliloti isolates. The linear correlation between salt tolerance of rhizobia and their symbiotic effectivity was revealed (r = 0,76). Proceeded data specified that bet-genes polymorphism at the north of Caucus gene center of alfalfa have been caused mainly by host-plant influence, whereas in salt affected area (Aral Sea region) the adaptation processes to abiotic stress factor have had a crucial role.
Key words: rhizobia, subpopulations, bet-genes, polymorphism, salt tolerance, symbiosis, alfalfa, gene centers.
ГНУ Всероссийский НИИ сельскохозяйственной |
Поступила в редакцию |




