








УДК 631.46:579.64:581.557:581.138.1:576.08
РАСТИТЕЛЬНО-МИКРОБНЫЕ СИМБИОЗЫ КАК ЭВОЛЮЦИОННО ЦЕЛОСТНЫЕ СИСТЕМЫ
Н.А. ПРОВОРОВ, Н.И. ВОРОБЬЕВ
Структурно-функциональная целостность растительно-микробных симбиозов относится к их фундаментальным свойствам, проявляясь на организменном, клеточном, молекулярно-генетическом и популяционном уровнях. С использованием математической модели эта целостность представлена как степень согласованности адаптивных реакций (варьирования генотипических частот) партнеров на изменения внешних условий. Индекс целостности симбиоза, определяемый на основе анализа ковариации адаптивных реакций партнеров, коррелирует с их репродуктивной активностью, отражающей экологическую эффективность симбиоза. Компьютерные эксперименты показали, что эффективность и целостность симбиоза координировано повышаются под действием отбора. Предложенная модель может быть использована для определения генетической структуры симбиозов, обеспечивающей высокую продуктивность растений.
Ключевые слова: микробно-растительные взаимодействия, эволюция симбиоза, математическое моделирование, симбиотическая азотфиксация, клубеньковые бактерии, экологически устойчивое растениеводство, структуры симбиозов, обеспечивающие высокую продуктивность растений.
Растительно-микробные симбиозы (РМС) - удобные модели для разработки фундаментальных аспектов симбиологии и симбиогенетики, так как растения вступают в разнообразные симбиотические отношения с микроорганизмами, выполняющими трофические, защитные и регуляторные функции (1). Будучи неспособными к использованию многих источников питания и к активной защите от антагонистов (патогенов, животных-фитофагов), растения компенсируют это за счет биохимических возможностей бактерий и грибов ? фиксации N2, растворения фосфатов, деструкции органических полимеров, осмотрофности, синтеза токсинов и антибиотиков.
К фундаментальным свойствам симбиоза относится его целостность, которая поддерживается обратными связями, позволяющими партнерам согласованно реагировать на внешние воздействия (2). Изучение азотфиксирующих клубеньков бобовых, арбускулярной микоризы, эндофитных и ризосферных ассоциаций демонстрирует связь целостности с морфофизиологической сложностью и экологической эффективностью симбиоза. Структурно-функциональная и генетическая целостность симбиоза формируется в процессе коэволюции партнеров и может быть изучена на уровне как структуры их популяций, так и активности перекрестно регулируемых генов.
Цель нашей работы - проанализировать эволюционные механизмы, определяющие целостность симбиозов, и на этой основе предложить генетические подходы для улучшения растительно-микробных симбиозов, используемых в экологически устойчивых агросистемах.
М о р ф о ф и з и о л о г и ч е с к а я ц е л о с т н о с т ь с и м б и о-
з а. Симбиозы относят к биологическим системам с относительно низкой по сравнению с унитарными организмами степенью целостности (2). Однако это свойство весьма лабильно: целостность симбиосистем может усиливаться в процессе эволюции, что позволяет анализировать соотношения между структурно-функциональной организованностью и экологической эффективностью симбиоза.
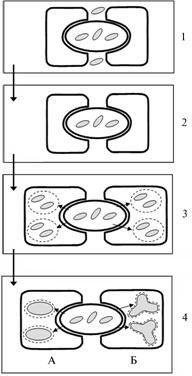
Целостность симбиоза непосредственно связана с его морфологической сложностью и степенью зависимости партнеров друг от друга, но эти признаки не всегда коррелирует между собой. Среди РМС наиболее сложно организованы азотфиксирующие бобово-ризобиальные симбиозы с многостадийными программами развития, которое начинается с инфицирования бактериями корня и закладки примордиев в его кортексе, а завершается формированием сложно устроенных органов ? клубеньков (3). Сравнительно-мор-фологический анализ позволил проследить эволюционное возрастание целостности симбиоза: сначала от внеклеточного поддержания бактерий (в инфекционных нитях или в межклетниках клубенька) к внутриклеточному симбиозу, образуемому в результате эндоцитоза, после которого бактерии формируют в растительных клетках стабильные симбиосомы, затем — от неспециализированных симбиосом, содержащих по несколько N2-фиксирующих бактероидов, внешне не отличающихся от свободноживущих бактерий, к специализированным симбиосомам с единичными структурно дифференцированными бактероидами (рис. 1). Важным фактором повышения целостности клубенькового симбиоза служит усложнение механизмов инфицирования растений бактериями ? от проникновения через разрывы эпидермиса, возникающие при росте боковых корней, до активного поглощения корневыми волосками, которое индуцируется бактериальными Nod-факторами, синтезируемыми под контролем nod-генов (4).
|
Рис. 1. Повышение целостности бобово-ризобиального симбиоза в процессе макроэволюции (5): 1 ? бесклубеньковые симбиозы, когда N2-фиксирующие бактерии поддерживаются в инфекционных нитях (ИН) и в межклеточных пространствах кортекса корня (Gleditsia, Ceratonia,Cercis); 2 ? примитивные клубеньки, в которых отсутствует эндоцитоз бактерий в растительные клетки и N2-фиксация происходит в ИН (Cassia,Andira,Hymenolobium); 3 ? эволюционно продвинутые клубеньки с неспециализированными (мультибактериальными) внутриклеточными симбиосомами, содержащими слабо дифференцированные N2-фиксирующие бактероиды (большинство бобовых); 4 ? то же со специализированными (монобактериальными) симбиосомами, включающими глубоко дифференцированные бактероиды (А ? Medicago, Б ? Pisum). Ярким примером повышения биохимической целостности симбиоза служит переход от амидного к уреидному типу ассимиляции фиксированного азота, характерному для бобовых из трибы Phaseoleae (5). Включение азота в состав уреидов (аллантоин, аллантоиновая кислота), в которых соотношение N:С составляет 1:1, резко снижает количество потребляемого клубеньками углерода (в амидах и аминокислотах это соотношение равно 1:2 или 1:3). Этот способ ассимиляции азота сопряжен с дифференцировкой растительных клеток центральной зоны клубенька ? их разделением на N2-фиксирующие (инфицированные) и синтезирующие уреиды (не инфицированные) клетки. Системная регуляция также повышает целостность симбиоза. Она обеспечивает сопряжение симбиотрофного питания бобовых азотом с «автотрофным» питанием (например, с усвоением нитратов) и с фотосинтезом (рис. 2). Ключевыми фак- |
торами авторегуляции симбиоза выступают растительные гены CLAVATA, которые контролируют развитие клубеньковых меристем и не имеют гомологов в системах защиты растений от патогенов. Принципиально важен факт синтеза сигналов, ограждающих рас- |
|
тение от формирования избытка клубеньков, в листьях, что связано с зависимостью симбиоза от интенсивности фотосинтеза. При этом питание растений фиксированным азотом в гораздо меньшей степени зависит от системной регуляции: ассимиляция нитратов и подавление ими образования клубеньков в основном определяются процессами, происходящими в корнях (см. рис. 2).
Таким образом, на примере клубенькообразования можно видеть, что целостность симбиоза определяется метаболическими и сигнальными обратными связями, которые складываются между партнерами на разных стадиях взаимодействия. На этапах узнавания и проникновения бактерий в растение эти связи отрицательные: они обеспечивают хозяину строгий контроль над численностью и размножением микросимбионтов, поэтому задействованные здесь механизмы похожи на защитные реакции растений против патогенов (6). На поздних стадиях обратные связи приобретают положительный характер, обеспечивая метаболическую интеграцию партнеров — эквивалентный обмен продуктами азотфиксации и фотосинтеза (7, 8), характерный для мутуализма.
Ц е л о с т н о с т ь с и м б и о з а к а к о б ъ е к т д е й с т в и я
о т б о р а. Морфофизиологическая целостность симбиоза тесно связана с его генетической целостностью, основанной на поддержании в клубеньках сложно организованной популяции бактерий, структура которой зависит от характера инфицирования корней. При проникновении бактерий через разрывы эпидермиса клубеньки часто содержат смеси штаммов, в том числе способных и не способных к N2-фиксации, тогда как при проникновении через корневые волоски основная часть возникающих клубеньков содержит клоны, формируемые при размножении единичных бактерий. Для эффективного симбиоза благоприятна клональная структура бактериальной популяции, которая способствует формированию положительных обратных связей растения с N2-фиксирующими эндосимбионтами: хозяин поставляет продукты фотосинтеза преимущественно в те клубеньки, из которых активно поступают азотные соединения. Против бактерий, находящихся в не фиксирующих N2 клубеньках, могут быть применены защитные реакции (9), что усиливает отбор в пользу N2-фиксирующих штаммов.
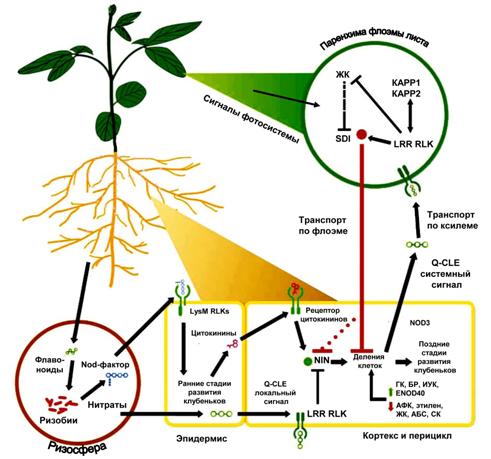 |
Рис. 2. Системная регуляция бобово-ризобиального симбиоза (18): LysM RLKs — рецептор-подобная киназа с доменами, связывающими олигохитиновые остатки; Q-CLE — сигнальные белки; LRR RLK — рецептор-подобная киназа с лейцин-богатыми повторами (узнает сигнальные факторы белковой природы); NIN — белок, активирующий развитие клубеньков при азотном голодании (гомологичен белку Mid Chlamydomonas, активирующему гаметогенез на безазотной среде); NOD3 — нодулин (клубенек-специфичный белок), накапливающийся в паренхиме клубенька; ENOD40 — нодулин, регулирующий баланс ауксинов и цитокининов при развитии клубенька; ГК — гибберелловая кислота; БР — брассиностероиды; ИУК — индолилуксусная кислота; АФК — активные формы кислорода; ЖК — жасмоновая кислота; АБС — абсцизовая кислота; СК — салициловая кислота; SDI — сигнальный фактор, синтезируемый в листьях и мигрирующий в корень, где он подавляет деление клеток, приводящее к образованию клубеньковых примордиев; KAPP1, KAPP2 — фосфорилазы белков, связанные с киназами. |
1. Коэффициенты корреляции индексов функциональной интегрированности симбиоза (ФИС) с его адаптивно значимыми свойствами, полученные с использованием математической модели (10) |
||||||
Индекс ФИС |
ЭМС |
Индекс полиморфизма (17) |
Частота бактериальных генотипов |
|||
растений |
бактерий |
Р |
М1 + М2 + М3 |
М3 |
||
InPM |
+0,728 |
–0,818 |
–0,978 |
–0,887 |
+0,887 |
+0,959 |
InM |
+0,708 |
–0,805 |
–0,973 |
–0,860 |
+0,860 |
+0,959 |
П р и м е ч а н и е. Моделируемая система состоит из диморфной популяции растений и низкополиморфной популяции бактерий — родительский штамм Р, который образует не фиксирующие N2 клубеньки с обоими растительными генотипами, и его мутанты, фиксирующие N2 только с одним (М1, М2) или с обоими (М3) генотипами растений. Индексы ФИС вычислены на основании анализа ковариации частот генотипов партнеров при малых изменениях системных параметров модели для всей системы (InPM) или для бактериальной популяции (InM). Критическое значение коэффициента корреляции (r) составляет 0,575 (P0 < 0,01); для частот М1 и М2 корреляции недостоверны. Эффективность мутуалистического симбиоза (ЭМС) определяли на основании числа семян, сформированных растениями после колонизации клубеньков бактериями. |
||||||
Исходя из того, что отбор на эффективность мутуализма, осуществляемый в эндосимбиотических популяциях ризобий, представляет собой результат действия положительных обратных связей партнеров, мы провели математическое моделирование микроэволюционных процессов, основанных на этом отборе (10). Анализ построенных моделей позволил нам связать эффективность мутуализма с функциональной интегрированностью симбиоза (ФИС), а также с генотипической специфичностью взаимодействия партнеров. Для оценки ФИС мы предложили индексы, которые отражают согласованность адаптивных реакций партнеров ? ковариацию их генотипических частот при изменениях параметров модели. Высокие корреляции эффективности симбиоза и ФИС (табл. 1) объясняются отбором в пользу интегрированности симбиоза, которая достигается в ходе его эволюции на повышение азотфиксирующей активности.
| 2. Адаптивно значимые свойства бобово-ризобиального симбиоза при различных схемах его развития, изученные с помощью математических моделей (10) | ||||
Схема развития (индекс ФИС) |
Набор штаммов, способных размножаться в клубеньках растений генотипа Г1 |
Доля генотипа Г1 в |
Соотношение давлений |
ЭМС |
С0 (0,216) |
Р, М1, М2, М3 |
50 |
+0,244 |
0,270 |
С1 (0,245) |
М1, М2, М3 |
58 |
–0,007 |
0,324 |
С2 (0,899) |
М1, М3 |
68 |
–1,071 |
0,376 |
П р и м е ч а н и е. Индексы функциональной интегрированности симбиоза (ФИС) варьируют от 1 (при максимальной согласованности адаптивных реакций партнеров) до 0. Отбор, действующий на растительный генотип Г1 со стороны «специфичного» мутуалиста М1, всегда положителен, со стороны «неспецифичного» мутуалиста М3 ? по мере ограничения круга штаммов, способных размножаться в клубеньках Г1 (С0 → С1 → С2), переходит от положительного к отрицательному, что отражает возрастание селективного преимущества М1 над М3. ЭМС ? эффективность мутуалистического симбиоза (см. табл. 1). |
||||
Анализ модели с точки зрения целостности симбиоза показал, что ФИС тесно связана с симбиотической эффективностью, которая возрастает при ограниченном размножении неактивных азотфиксаторов в клубеньках. Оно определяется усложнением инфекционного процесса, приводящим к координированному повышению интегрированности, генотипической специфичности и эффективности симбиоза. Так, при ограниченном размножении неактивных штаммов в клубеньках усиливается селективное преимущество специфичных мутуалистов над неспецифичными (табл. 2), вследствие чего одновременно повышается экологическая эффективность и генотипическая специфичность мутуализма (см. табл. 1). Эта связь ранее была выявлена экспериментально при изучении межвидового (11) и внутривидового (12) варьирования у бобовых. Важно отметить, что полной элиминации неспецифичных мутуалистов из системы не происходит, поскольку они обеспечивают высокий уровень общей эффективности симбиоза (см. табл. 1).
Таким образом, ФИС можно рассматривать как экологически значимый показатель симбиоза, который повышается в процессе адаптивной эволюции. В то же время связь ФИС со структурной и биохимической целостностью симбиоза, возрастающей в ходе его макроэволюции, а также с генетической целостностью, которая выражается в формировании симбиогенома (1), требует специального изучения.
П р а к т и ч е с к и е а с п е к т ы. Изучение целостности симбиозов имеет огромное практическое значение, поскольку позволяет формировать генетически обоснованную методологию их улучшения и использования в адаптивном земледелии. Анализ предложенной нами математической модели (10) предоставляет возможность наметить основные направления оптимизации генетической структуры симбиосистемы, в первую очередь соотношения долей специфичных и неспецифичных мутуалистов, которое повышается при формировании клональной популяции ризобий. Эксперименты показывают, что при выращивании бобовых в полевых условиях клоны ризобий содержатся только в части (40-95 %) клубеньков (13). Однако степень клональности внутриклубеньковой популяции ризобий может быть повышена, например за счет ускорения инфекционного процесса, при котором корневые волоски захватывают клетки бактерий. В частности, такое ускорение достигается благодаря дефициту азота, повышающему скорость перестроек стенок корневого волоска, через которые происходит поглощение ризобий (14), или активной экскреции растением флавоноидов, активирующих nod-гены (15, 16), что приводит к росту числа клубеньков.
Знания об эволюционных основах целостности симбиосистем могут использоваться в генетико-селекционных работах по их улучшению. Наибольшая эффективность симбиоза обеспечивается при взаимодействии двух комплементарных друг другу (специально подобранных или сконструированных) генотипов растений и бактерий. Однако в реальных симбиотических системах оба партнера генетически гетерогенны: у растений полиморфизм определяет экологическую пластичность сортов, у бактерий он обусловлен нестабильностью генома, а также присутствием в почвах спонтанной микрофлоры, конкурирующей с производственными штаммами за инфицирование растений. Следовательно, важной для практики задачей служит формирование моделей (прототипов) эффективных систем ? построение их оптимальных популяционно-генетических структур, которое следует проводить в сочетании с конструированием индивидуальных генотипов растений и бактерий (8). Для решения этой задачи целесобразно использовать предложенную нами математическую модель (10), показавшую, что эффективность симбиоза максимальна при умеренном полиморфизме растительной популяции, которая в простейшем случае состоит из доминирующего и минорного генотипов. Такая популяция развивает наиболее эффективный симбиоз при взаимодействии с двумя штаммами ризобий, один из которых специфичен к доминирующему генотипу растений, а другой взаимодействует с обоими генотипами растений (неспецифичный симбионт) (5).
Итак, важнейшее условие успешного улучшения растительно-микробных симбиозов ? перенос знаний об их естественной эволюции в сферу селекции, генной инженерии и биотехнологии. Правомерность такого подхода определяется тем, что повышение экологической эффективности симбиозов, необходимое для создания экологически устойчивых систем растениеводства, представляет собой магистральное направление их естественной эволюции. В переносе знаний из фундаментальных областей биологии в агробиологию огромную роль играют математические модели, которые могут быть использованы для построения прототипов эффективных симбиотических систем.
Л и т е р а т у р а
1. Т и х о н о в и ч И.А., П р о в о р о в Н.А. Симбиозы растений и микроорганизмов: молекулярная генетика агросистем будущего. СПб, 2009.
2. Ш а п о ш н и к о в Г.Х. Живые системы с малой степенью целостности. Журн. общ. биол., 1975, 36: 323-335.
3. B r e w i n N.J. Plant cell wall remodeling in the Rhizobium-legume symbiosis. Crit. Rev. Plant. Sci., 2004, 23: 1-24.
4. S p r e n t J.I. Evolving ideas of legume evolution and diversity: a taxonomic perspective on the ocurrence of nodulation. New Phytologist, 2007, 174: 11-25.
5. P r o v o r o v N.A., V o r o b y o v N.I. Evolutionary genetics of plant-microbe symbioses /I.A. Tikhonovich (ed.). N.Y., 2010.
6. J o n e s K.M., K o b a y a s h i H., D a v i e s B.W. e.a. How rhizobial symbionts invade plants: the Sinorhizobium—Medicago model. Nature Review, 2007, 5: 619-633.
7. П р о в о р о в Н.А., В о р о б ь е в Н.И., А н д р о н о в Е.Е. Макро- и микроэволюция бактерий в системах симбиоза. Генетика, 2008, 44: 12-28.
8. О н и щ у к О.П., В о р о б ь е в Н.И., П р о в о р о в Н.А. и др. Симбиотическая активность ризобий люцерны (Sinorhizobium meliloti) с генетическими модификациями системы транспорта дикарбоновых кислот. Экологическая генетика, 2009, 7: 3-10.
9. D e n i s o n R.F. Legume sanctions and the evolution of symbiotic cooperation by rhizobia. The American Naturalist, 2000, 156: 567-576.
10. В о р о б ь е в Н.И., П р о в о р о в Н.А. Моделирование эволюции бобово-ризобиального симбиоза на повышение функциональной интегрированности партнеров и экологической эффективности их взаимодействия. Экологическая генетика, 2010, 8(3): 16-26.
11. Д о р о с и н с к и й Л.М., Л а з а р е в а Н.М. О специфичности клубеньковых бактерий сои и люпина. Микробиология, 1968, 37: 115-121.
12. П р о в о р о в Н.А., Т и х о н о в и ч И.А. Эколого-генетические принципы селекции растений на повышение эффективности взаимодействия с микроорганизмами. С.-х. биол., 2003, 3: 11-25.
13. K i e r s E.T., R o u s s e a u R.A., W e s t S.A. e.a. Host sanctions and the legume-Rhizobium mutualism. Nature,2003, 425: 78-81.
14. Я к о в л е в а З.М. Бактероиды клубеньковых бактерий. Новосибирск, 1975.
15. K a p u l n i k J., J o s e p h C.M., P h i l l i p s D.A. Flavone limitation to root nodulation and symbiotic nitrogen fixation in alfalfa. Plant Physiol., 1987, 84: 1193-1196.
16. H e r n a n d e z G., R a m i r e z M., S a u r e z R. Root-exuded nod-gene inducing signals limit the nodulation capacity of different alfalfa varieties with Rhizobium meliloti. Plant Cell Reports, 1995, 14: 626-629.
17. N e i M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 1978, 89: 583-590.
18. F e r g u s s o n B.J., I n d r a s u m u n a r A., H a y s h i S. e.a. Molecular analysis of legume nodule development and autoregulation. J. Integr. Plant Biol., 2010, 52: 61-76.
PLANT-MICROBE SYMBIOSES AS THE EVOLUTIONARY INTEGRATED SYSTEMS
N.A. Provorov, N.I. Vorobyov
The structural and functional integrity of plant-microbe symbioses represents their fundamental property expressed at the organismic, cellular, molecular-genetic and population levels. Using the mathematical model this integrity is represented as a degree of concordance of adaptive reactions (alterations of the partners’ frequencies) for the environmental changes. The indices of symbiosis integrity determined using the partners’ frequencies co-variation correlate to the symbiosis efficiency determined as its impacts on the partners’ reproductive activities. The computer experiments suggest that efficiency and integrity of symbiosis may increase in parallel under impacts of natural selection. The developed model may be used to create the symbioses genetic structures ensuring the high plant productivity.
Keywords: microbe-plant interactions, evolution of symbiosis, mathematical simulation, symbiotic N2 fixation, nodule bacteria, ecologically sustainable agriculture, structures of symbioses ensuring high plant productivity.
ГНУ Всероссийский НИИ сельскохозяйственной |
Поступила в редакцию |




