








УДК 633.31:581.557:631.461.5:575:575.224.46
ИСПОЛЬЗОВАНИЕ МЕТОДА САЙТ-НАПРАВЛЕННОГО МУТАГЕНЕЗА ДЛЯ ИЗУЧЕНИЯ ФУНКЦИЙ ГЕНА SMb20332 У КЛУБЕНЬКОВЫХ БАКТЕРИЙ Sinorhizobium meliloti
Е.П. ЧИЖЕВСКАЯ, О.П. ОНИЩУК, Е.Е. АНДРОНОВ, Б.В. СИМАРОВ
С использованием сайт-направленного мутагенеза у типового штамма 1021 клубеньковых бактерий люцерны Sinorhizobium meliloti получен мутант по гену SMb20332. Этот ген расположен на мегаплазмиде 2 и по результатам секвенирования предположительно кодирует арилсульфатазу. У полученного мутанта ACH-5 тестировали симбиотические свойства, солеустойчивость и способность к росту на диагностических средах. Обнаружено, что арилсульфатазы клубеньковых бактерий, как и арилсульфатазы морских бактерий, способны десульфатировать агаропектин. Мутация по гену SMb20332 не оказывала существенного влияния на солеустойчивость штамма 1021 S. meliloti, а также на его эффективность и конкурентоспособность при симбиозе с Medicago sativa (сорт Вега). Впервые показано, что у штамма 1021 S. meliloti мутация по гену SMb20332 приводит к изменению поверхностных полисахаридов и утрате устойчивости к ингибитору дыхания бактерий — гербициду 2М-4ХМ.
Ключевые слова: Sinorhizobium meliloti, арилсульфатаза, продукция полисахаридов, устойчивость к гербициду, солеустойчивость, конкурентоспособность, симбиотическая эффективность.
Клубеньковые бактерии люцерны(Sinorhizobium meliloti) способны фиксировать молекулярный азот атмосферы в симбиозе с бобовыми растениями — люцерной, донником и пажитником. Геном этого микроорганизма представлен хромосомой и двумя гигантскими мегаплазмидами, размер которых варьирует от 1100 до 1900 т.п.н. (1). Так, у типового штамма 1021 S. meliloti размер мегаплазмид 1 (pSymA) и 2 (pSymB) составляет соответственно 1354 т.п.н. и 1683 т.п.н. (2, 3). К настоящему времени геном штамма 1021полностью секвенирован, однако функции многих генов остаются неизвестными (2-4).
Ген SMb20332 штамма 1021 S. meliloti расположен на мегаплазмиде 2 и по результатам секвенирования предположительно кодирует фермент арилсульфатазу (3). Выбор указанного гена как объекта исследования был обусловлен следующими причинами. Рядом с геном SMb20332 расположен ген betT (SMb20333) (рис. 1, А), вовлеченный в транспорт осмопротектора глицин-бетаина (3, 5). Ген betT конститутивно экспрессируется как у свободноживущих штаммов S. meliloti, так и в условиях симбиоза с растением-хозяином (в инфекционных нитях и бактероидах) (5, 6). Функции второго смежного гена SMb20331 неизвестны, но показано, что он экспрессируется в условиях осмотического шока (7). Таким образом, изучаемый ген SMb20332 находится в окружении генов, связанных с контролем такого экологически важного признака как солеустойчивость, поэтому логично было предположить участие арилсульфатазы в контроле солеустойчивости у S. meliloti.
Для проверки этой гипотезы и выявления функций гена SMb20332 с использованием сайт-направленного мутагенеза мы получили мутант по этому гену и охарактеризована его солеустойчивость, симбиотические и культурально-биохимические свойства.
Методика. Объектами исследования служили типовой штамм 1021 клубеньковых бактерий люцерны S. meliloti (SmR, мутант природного штамма SU47) (8) и полученный на его основе мутант по гену SMb20332, обозначенный нами как ACH-5. Также в работе использовали штаммы Eshcherichacoli: DH5a [F-, recA1, endA1, gyrA96, hsdR17, deoR, D(lacZYA-argF)U169] (9) — для трансформации клеток плазмидами и S17-1 (SmR), содержащий интегрированную в хромосому модифицированную плазмиду pRP4 (Tc::Mu, Km::Tn7) (10), — в качестве мобилизующего штамма для конъюгационного переноса плазмид в клетки S.meliloti. Использованные плазмиды: вектор для А/Т клонирования pTZ57R/T (lacZ::M13mp, ApR) («MBI Fermentas», Литва), pK18mob (mob, lacZ::M13mp, ApR, KmR) (11), pMP4655 (содержит ген gfp под контролем Plac в pME6010, TcR) (12).
Клубеньковые бактерии S. meliloti культивировали при 28 °С на среде TY (13). При выращивании в жидкой среде проводили дополнительную аэрацию на лабораторной качалке в режиме 180 об/мин. В селективные среды добавляли антибиотики Sm и Km (соответственно 800 и 200 мг/л). Штаммы E. coli выращивали при 37 °С на среде LB (14) (в жидкой среде — с аэрацией 220 об/мин), в селективные среды добавляли антибиотики Ap, Sm, Km и Tc (соответственно 100, 100, 50 и 15 мг/л).
Солеустойчивость штаммов тестировали в жидкой и на агаризованной среде TY, содержащей различные концентрации NaCl, по методике, приведенной ранее (15).
Для определения изменений в синтезе полисахаридов использовали среду 79 (16) и среду 79, содержащую индикатор восстановительных эквивалентов хлорид трифенилтетразолия (TTХ, 0,01 %). Устойчивость к гербициду тестировали на среде 79 с добавлением 2-метил-4-хлорфеноксимасляной кислоты (2М-4ХМ, 0,1 %), ингибирующей дыхание бактерий (17). Также использовали стандартные среды: минимальную (18) — для анализа слизеобразования, минимальную с добавлением красителя Конго красный (0,02 %) — для оценки продукции липо- и экзополисахаридов (ЛПС и ЭПС) (19, 20), минимальную c добавлением калькофлуора (0,02 %) — для анализа продукции ЭПС1 (сукциноглюкана) (19, 21). Рост штаммов в отсутствие неорганического сульфата анализировали в жидкой и на агаризованной минимальной среде без добавления MgSO4.
Геномную ДНК S. meliloti выделяли по стандартной методике (22), плазмидную из клеток E. coli — общепринятым методом щелочного лизиса (14). Реакции рестрикции и лигирования, электрофорез в агарозном геле и выделение фрагментов ДНК из геля осуществляли с помощью стандартных приемов (14). Плазмиды вводили в штаммы E. coli методом высокоэффективной трансформации (23). ПЦР-амплификацию фрагмента ДНК, содержащего ген SMb20332, проводили с использованием сконструированных нами праймеров betS1-F (5´-GCTTCTTCTTCCTGCCATCA-3´) и betS1-R (5´-CACCTTCTTTTCGGACGGTA-3´).
При конъюгации ночные культуры мобилизующего штамма-донора S17-1 E. coli и штамма-реципиента 1021 S. meliloti смешивали в соотношении 1:5 и переносили каплями на поверхность твердой среды TY. После впитывания капель конъюгационную смесь инкубировали в течение 1 сут при 28 °С. Выросшие бактериальные клетки суспендировали в 1 мл стерильной воды, готовили 10- и 100-кратные разведения и высевали по 0,1 мл суспензии на среду 79, содержащую Km (200 мг/л). Отбор и анализ трансконъюгантов S. meliloti проводили на 4-5-е сут.
Симбиотические свойства штаммов изучали в условиях стерильного микровегетационного опыта (24), в качестве растения-хозяина использовали люцерну посевную Medicagosativa (сорт Вега). Инокуляцию проростков люцерны проводили 1 мл суспензии клеток клубеньковых бактерий (105-106 кл/мл). Растения выращивали в теплице при температуре 20-22 °С, дополнительном освещении 20 000 лк и 16-часовом фотопериоде в течение 28-30 сут, повторность опытов 6-кратная. Скорость клубенькообразования (25) определяли
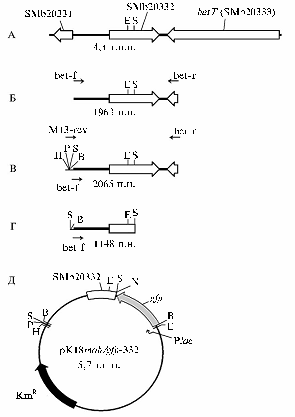 |
Рис. 1. Этапы конструирования плазмиды pK18mob/gfp-332 (описание см. в тексте): А — локус SMb20332 штамма 1021 Sinorhizobiummeliloti, Б — ПЦР-амплифицированный фрагмент ДНК, содержащий ген SMb20332, В — ПЦР-амплифицированный фрагмент ДНК, содержащий ген SMb20332 и часть полилинкера вектора pTZ57R/T, Г — SalI-фрагмент, содержащий первые 404 п.н. кодирующей области гена SMb20332, Д — плазмида pK18mob/gfp-332, несущая ген gfp и фрагмент гена SMb20332 в векторе pK18mob; сайты рестрикции В, Е, Н, Р, S и Х — соответственно BamHI, EcoRI, HindIII, PstI, SalI и XbaI. |
при подсчете клубеньков на корнях растений на 6-е, 14-е и 19-е сут после инокуляции. Симбиотическую эффективность штаммов оценивали по сухой массе надземных частей инокулированных растений относительно контроля без инокуляции на 30-е сут вегетации. Нодуляционную конкурентоспособность определяли по соотношению числа клубеньков, образованных мутантом при совместной инокуляции с родительским штаммом 1021 S. meliloti. Штаммы смешивали в растворе микроэлементов при равной оптической плотности суспензий, полученную смесь (»108 кл/мл) использовали для инокуляции 2-суточных проростков люцерны. Через 30 сут после инокуляции бактерии выделяли из клубеньков и анализировали их рост на среде TY и среде TY с антибиотиками. Долю клубеньков, содержащих Kmr клоны (мутант), сопоставляли с долей Kmr клонов в исходном инокулюме, которую определяли после высева на среду TY с антибиотиками (25, 26).
Для статистической обработки данных использовали t-критерий Стьюдента.
Результаты. П о л у ч е-
н и е м у т а н т а ACH-5.Для мутагенеза гена SMb20332 был применен широко известный метод сайт-направленной интеграции плазмиды pK18mob в геном родительского штамма 1021 S.meli-loti. Плазмида pK18mob (размер 3,8 т.п.н.) является мультикопийным вектором для клонирования фрагментов ДНК, несет ген устойчивости к канамицину и содержит полилинкер плазмиды pUC18 (11). Плазмида pK18mob как мобилизуемый вектор при конъюгации с использованием плазмиды-помощника или мобилизующего штамма может передаваться из клеток E. coli в широкий круг грамотрицательных и грамположительных бактерий (27). Однако pK18mob способна реплицироваться только в бактериях, относящихся к родам Escherichia, Salmonella и Serratia (так как имеет ориджин репликации плазмиды pMB1) (28). Таким образом, сохранение вектора pK18mob в клетках клубеньковых бактерий может происходить лишь в результате его интеграции в геном. При правильном подборе области гомологии можно добиться интеграции вектора в строго определенную часть генома, в том числе в кодирующую область изучаемого гена, что приведет к нарушению его функции.
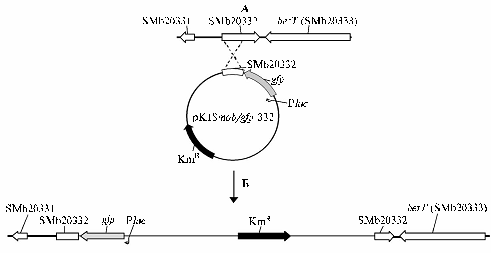
Конструирование плазмиды pK18mob/gfp-332, содержащей область гомологии, включало несколько этапов. Сначала был получен вариант вектора pK18mob, несущий ген gfp (green fluorescent protein) под контролем промотора P(lac), что дало дополнительный маркер при наблюдениях за мутантным штаммом в вегетационных опытах. Для этого BamHI-XbaI фрагмент (753 п.н.), несущий ген gfp, был вырезан из плазмиды pMP4655 (12) и клонирован в соответствующие сайты полилинкера pK18mob. Далее с помощью праймеров betS1-F и betS1-R был амплифицирован фрагмент генома штамма 1021 S.meliloti (1963 п.н.), содержащий генSMb20332 (см. рис. 1, Б). ПЦР-фрагмент был клонирован в векторе pTZ57R/T, затем из полученной плазмиды с использованием праймеров M13-rev (26) и betS1-R был реамплифицирован фрагмент с прилегающей частью полилинкера вектора pTZ57R/T (см. рис. 1, В). Из этого ПЦР-фрагмента был вырезан SalI-фрагмент (1148 п.н.), содержащий первые 404 п.н. кодирующей области гена SMb20332 (см. рис. 1, Г), и клонирован в SalI сайт полилинкера pK18mob-gfp. Далее при BamHI-рестрикци-онном анализе были отобраны плазмиды, несущие клонированный фрагмент ризобиальной ДНК в определенной ориентации относительно полилинкера pK18mob и гена gfp (см. рис. 1, Д). Сконструированная плазмида (pK18mob/gfp-332) методом трансформации была перенесена в мобилизующий штамм S17-1 E. coli. Затем был проведен конъюгационный перенос pK18mob/gfp-332 из штамма S17-1 в штамм S. meliloti 1021 и отбор KmRGFP+ трансконъюгантов. Появление KmRGFP+ клонов S. meliloti указывает на интеграцию pK18mob/gfp-332 в геном штамма 1021 по участку гомологии (рис. 2, А), что нарушает целостность открытой рамки считывания гена SMb20332 (см. рис. 2, Б).
|
Рис. 2. Схема сайт-направленного мутагенеза гена SMb20332: А — гомологичная рекомбинация участка генома штамма 1021 Sinorhizobiummeliloti и фрагмента плазмиды pK18mob/gfp-332, Б — интеграция плазмиды pK18mob/gfp-332 в ризобиальный геном по участку гомологии. |
С о л е у с т о й ч и в о с т ь. Поскольку ген SMb20332 расположен в одном локусе с генами, участвующими в контроле солеустойчивости у штаммов S. meliloti, мы изучили способность мутанта ACH-5 и его родительского штамма 1021 к росту в жидкой и на агаризованной среде TY с различным содержанием NaCl. В жидкой среде с 2,0 % NaCl у мутанта ACH-5 и штамма 1021 рост был нормальный, а при 4,5 % NaCl — полностью отсутствовал. При содержании 3,5 % NaCl у штамма 1021 наблюдался нестабильный рост, а у мутанта ACH-5 — отсутствие роста. На агаризованной среде у мутанта и его родительского штамма рост не различался: при добавлении 2,0 % NaCl рост был нормальным, а при 3,0 и 3,5 % NaCl — отсутствовал. Таким образом, мутация в гене SMb20332 у штамма 1021 S. meliloti привела только к утрате его способности к неустойчивому росту в жидкой среде TY с 3,5 % NaCl.
С и н т е з п о л и с а х а р и д о в и у с т о й ч и в о с т ь к г е р б и ц и д у 2М-4ХМ.При тестировании способности к слизеобразованию, а также росту и окрашиванию клонов на диагностических средах, используемых для анализа состава поверхностных полисахаридов (табл. 1), оказалось, что по росту на минимальной среде мутант ACH-5 не отличался от своего родительского штамма. На минимальной среде c добавлением калькофлуора у колоний, сформированных как мутантным, так и родительским штаммами, наблюдалось характерное свечение в УФ-свете, указывающее на продукцию сукциноглюкана (ЭПС-1). Однако при добавлении в минимальную среду красителя Конго красного было обнаружено различие в интенсивности окраски: родительский штамм формировал колонии яркого красно-розового цвета, а мутант ACH-5 — бледно-розового. Изменение окраски штаммов на среде с Конго красным определяется главным образом адсорбционными свойствами их клеточных стенок и указывает на различия в строении поверхностных полисахаридов (19).
Кроме того, полученный мутант характризовался сниженным слизеобразованием при росте на среде 79, что также указывает на изменение способности к синтезу полисахаридов. На среде 79 с TTХ колонии различались: штамм 1021 формировал ярко-красные колонии с узким белым ободком (окрашено около 90 % площади колонии), а мутант ACH-5 — белые колонии с окрашенной центральной частью (менее 20 % площади). Окрашивание колоний на среде с ТТХ зависит от окислительно-восстанови-тельного потенциала клеток и наиболее интенсивно проявляется при ограниченном доступе кислорода (29). Родительский штамм 1021 продуцирует большое количество внеклеточных полисахаридов (слизи), в связи с чем его клетки находятся в микроаэробных условиях и интенсивно окрашиваются на среде с ТТХ. Мутант ACH-5 формирует малослизистые колонии, поэтому окрашивание наблюдается лишь внутри глубинной центральной части колоний, где клетки могут испытывать недостаток кислорода.
| 1. Показатели роста у родительского (1021) и полученного мутантного (АСН-5) штаммов Sinоrhizobiummeliloti на диагностических средах | ||||||
| Штамм< | Минимальная среда |
Среда 79 |
||||
без добавок |
с калько-флуором |
с Конго |
без добавок |
с ТТХ |
с ТТХ и |
|
1021 |
Белые малослизистые колонии |
Свечение в УФ-свете |
Колонии красно-розового цвета |
Нормальное слизе- |
Колонии красного цвета с узким белым ободком, слизистые |
Колонии белого цвета |
ACH-5 |
Белые малослизистые колонии |
Свечение в УФ-свете |
Колонии розового цвета |
Сниженное слизеоб-разование |
Колонии белого цвета с красным цент-ром, менее слизистые |
Нет роста |
П р и м е ч а н и е. TTХ — хлорид трифенилтетразолия, 2М-4ХМ — 2-метил-4-хлорфеноксимасляная кислота. |
||||||
Родительский штамм 1021 был способен к росту на среде 79, содержащей ТТХ и гербицид 2М-4ХМ, однако при этом его клетки утрачивали способность к окрашиванию. Вероятно, гербицид 2М-4ХМ вызывает у штамма 1021 снижение активности функционирования дыхательной цепи транспорта электронов и, как следствие, изменение окислительно-восстановительного потенциала клеток. В отличие от родительского штамма у мутанта ACH-5 наблюдалось полное подавление роста на среде с 2М-4ХМ (см. табл. 1).
Р о с т в о т с у т с т в и е н е о р г а н и ч е с к о г о с у л ь ф а т а. Известно, что арилсульфатазы морских бактерий способны осуществлять гидролиз агаропектина (компонента агара) с высвобождением сульфата (31-33). В связи с этим мы тестировали аналогичные свойства изучаемых штаммов. В жидкой бессульфатной среде наблюдалось полное отсутствие роста как родительского, так и мутантного штаммов. Однако на агаризованной бессульфатной среде рост отсутствовал только у мутанта ACH-5, а родительский штамм формировал нормальные колонии. Поскольку единственным источником сульфата в используемой среде был агар, мы сделали заключение, что продукт гена SMb20332 способен десульфатировать агаропектин. Освобождающийся при этом сульфат полностью покрывал его дефицит в исходной среде, что позволяло штамму 1021 S. meliloti осуществлять нормальный рост в условиях сульфатного голодания.
2. Динамика клубенькообразования и симбиотическая эффективность у исходного (1021) и полученного мутантного (АСН-5) штаммов Sinоrhizobiummelilotiна корнях люцерны посевной Medicagosativa сорта Вега (Х±х) |
||||
| Штамм | Число клубеньков, шт. |
Масса растений, мг |
||
6-е сут |
14-е сут |
19-е сут |
||
1021 |
0 |
0,5±0,3 |
1,7±0,3 |
15,4±2,0 |
ACH-5 |
0 |
0,5±0,3 |
1,3±0,6 |
14,0±2,0 |
Контроль (без инокуляции) |
0 |
0 |
0 |
7,8±2,0 |
С и м б и о т и ч е -
с к и е с в о й с т в а у
и з у
ч а е м ы х ш т а м м о в. По скорости клубенькообразования и симбиотической эф-фективности мутант ACH-5 и его родительский штамм достоверно не различались (табл. 2). Также достоверно не различалась нодуляционная конкурентоспособность мутанта ACH-5 и его родительского штамма при совместной инокуляции (в соотношении близком 1:1). В исходном инокулюме доля клеток ACH-5 и штамма 1021 составила соответственно 43,6±6,15 и 56,4±6,15 %, а из образовавшихся клубеньков изучаемые штаммы были выделены в соотношении соответственно 33,3±7,36 и 67,7±7,36 %.
Внесение в среду для выращивания растений повышенной концентрации NaCl (0,3 %) не приводило к достоверному изменению конкурентоспособности штаммов: в исходном инокулюме клетки мутанта ACH-5 составляли 56,8±7,50 %, в клубеньках — 58,1±5,12 %. Таким образом, результаты проведенных микровегетационных опытов показали, что мутация в гене SMb20332 не влияет на основные симбиотические свойства штамма 1021 S. meliloti при симбиозе с M. sativa (сорт Вега).
Таким образом, мы представили анализ некоторых культурально-биохимиче-ских и симбиотических свойств у мутанта S. meliloti по гену SMb20332. Ген SMb20332 предположительно кодирует арилсульфатазу, однако его функции до конца не известны (3). Арилсульфатазы (арилсульфат-сульфогидролазы, EC 3.1.6.l) — ферменты класса гидролаз, катализирующие гидролиз ароматических сульфокислот по связи О–S: H2O + Aryl-O-SO3- ↔ Aryl-OH + SO42- + H+. Арилсульфатазы обнаружены во всех основных группах организмов (животные, растения, грибы, бактерии) и представляют собой высококонсервативное семейство ферментов (34). Отсутствие некоторых арилсульфатаз у человека вызывает тяжелое заболевание — метахроматическую лейкодистрофию. Среди прокариотов арилсульфртазы наиболее изучены у патогенных и условно патогенных бактерий, например у Klebsiellapneumoniae (35), Salmonellatyphimurium (36), Pseudomonasaeruginosa (37), E.coli (38). При этом показано, что арилсульфатазы могут играть важную роль в преодолении гематоэнцефалического барьера и, как следствие, влиять на патогенность штаммов (38). При изучении арилсульфатаз у морских бактерий, таких как Alteromonas carrageenovora, A. atlantica, Sphingomonas sp. была обнаружена способность осуществлять десульфатирование агаропектина, представляющего собой составную часть агара — структурного полисахарида клеточных стенок красных водорослей (31-33).
У почвенных бактерий арилсульфатазы почти не изучены, однако предполагают, что в бедных почвах при недостатке неорганического сульфата этот фермент может принимать участие в снабжении клеток сульфатом, гидролизуя некоторые компоненты гуминовых кислот, например 2-гидрокси-5-нитрофенилсульфат и фенилсульфат (39). У клубеньковых бактерий арилсульфатазы не изучались, поэтому об их функциях (как у свободноживущих клеток, так и в условиях симбиоза с растениями) ничего не известно. Однако при полном секвенировании геномов у некоторых представителей семейства Rhizobiaceae, например S.meliloti (2-4), S. medicae, Rhizobium leguminosarum bv. viciae (40), R. leguminosarum bv. trifolii, R. etli, Agrobacteriumradiobacter (41), были обнаружены гены, сходные с генами арилсульфатаз других бактерий.
Ген SMb20332 у штамма 1021 клубеньковых бактерий люцерны, предположительно кодирующий арилсульфатазу, как уже отмечалось, расположен в окружении генов, участвующих в контроле солеустойчивости (3, 5-7). Кроме того, известно, что фермент из семейства сульфатаз — холинсульфатаза кодируется геном betC, который входит в состав betICBA-оперона, ответственного за синтез глицин-бетаина и относящегося к одной из систем защиты клеток от осмотического шока (42). В связи с этим мы выдвинули предположение об участии арилсульфатазы в контроле солеустойчивости у ризобиальных штаммов.
Однако мутация в гене SMb20332 существенно не повлияла на солеустойчивость штамма 1021 S. meliloti, а также на его основные симбиотические свойства — эффективность, скорость образования клубеньков и нодуляционную конкурентоспособность при симбиозе с M. sativa (сорт Вега).
Обнаруженная у родительского штамма 1021 способность к росту на агаризованной бессульфатной среде и ее утрата у мутанта по гену SMb20332 указывают на то, что арилсульфатазы клубеньковых бактерий (как и арилсульфатазы морских бактерий) способны осуществлять десульфатирование агаропектина.
У полученного мутанта ACH-5 выявлены особенности культурально-морфоло-гических свойств, свидетельствующие об изменениях в синтезе поверхностных полисахаридов по сравнению с родительским штаммом. Также обнаружено, что мутация в гене SMb20332 приводит к утрате устойчивости штамма 1021 к гербициду 2М-4ХМ. Отметим, что механизм устойчивости штамма 1021 S. meliloti к этому гербициду неизвестен. Возможно, он связан со способностью продуцировать большое количество внеклеточных полисахаридов, адсорбирующих гербицид и тем самым ослабляющих его действие. В этом случае снижение количества синтезируемых полисахаридов и/или изменение их состава, которое наблюдалось у мутанта ACH-5, может опосредованно влиять на устойчивость штамма к гербицидам.
Итак, нами получен мутант ACH-5, содержащий удобный маркер (ген gfp в составе интегрированной в геном плазмиды pK18mob), который может быть в дальнейшем использован в экспериментах по изучению механизмов взаимодействия микроорганизмов в ризосфере и почве (43, 44). Впервые показано, что арилсульфатаза влияет на синтез внеклеточных полисахаридов клубеньковых бактерий люцерны, однако конкретный механизм этого влияния требует дальнейшего изучения.
Л И Т Е Р А Т У Р А
1. G u o H., S u n S., E a r d l y B. e.a. Genome variation in the symbiotic nitrogen-fixing bacterium Sinorhizobium meliloti. Genome, 2009, 52(10): 862-75.
2. B a r n e t t M.J., F i s h e r R.F., J o n e s T. e.a. Nucleotide sequence and predicted functions of the entire Sinorhizobium meliloti pSymA megaplasmid. PNAS, 2001, 98(17): 9883-9888.
3. F i n a n T.M., W e i d n e r S., W o n g K. e.a. The complete sequence of the 1,683-kb pSymB megaplasmid from the N2-fixing endosymbiont Sinorhizobium meliloti. PNAS, 2001, 98(17): 9889-9894.
4. C a p e l a D., B a r l o y - H u b l e r F., G o u z y J. e.a. Analysis of the chromosome sequence of the legume symbiont Sinorhizobium meliloti strain 1021. PNAS, 2001, 98(17): 9877-9882.
5. B o s c a r i A., M a n d o n K., D u p o n t L. e.a. BetS is a major glycine betaine/proline betaine transporter required for early osmotic adjustment in Sinorhizobium meliloti. J. Bacteriol., 2002, 184(10): 2654-2663.
6. B o s c a r i A., V a n d e S y p e G., L e R u d u l i e r D. e.a. Overexpression of BetS, a Sinorhizobium meliloti high-affinity betaine transporter, in bacteroids from Medicago sativa nodules sustains nitrogen fixation during early salt stress adaptation. Mol. Plant Microbe Interact., 2006, 19: 896-903.
7. D o m i n g u e z - F e r r e r a s A., P e t e r - A r n e d o R., B e c k e r A. e.a. Transcriptome profiling reveals the importance of plasmid pSymB for osmoadaptation of Sinorhizobium meliloti. J. Bacteriol., 2006, 188(21): 7617-7625.
8. M e a d e H.M., L o n g S.R., R u v k u n G.B. e.a. Physical and genetic characterization of symbiotic and auxotrophic mutants of Rhizobium meliloti induced by transposon mutagenesis. J. Bacteriol., 1982, 149(1): 114-122.
9. G r a n t S.G.N., J e s s e e J., B l o o m F.R. e.a. Differential plasmid rescue from transgenic mouse DNAs into Escherichia coli methylation-restriction mutant. PNAS, 1990, 87: 4645-4649.
10. S i m o n R., P r i e f e r U., P u h l e r A. A broad host range mobilization system for in vivo genetic engenering: Transposon mutagenesis in gram negative bacteria. Biotechnology, 1983, 1(11): 784-791.
11. S c h a f e r A., T a u c h A., J a g e r W. e.a. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19 selection of defined deletions in the chromosome Corynebacterium glutamicum. Gene, 1994, 145: 69-73.
12. B l o e m b e r g G.V., W i j f j e s A.H.M., L a m e r s G.E.M. e.a. Simultaneous imaging of Pseudomonas fluorescens WCS365 populations expressing three different autofluorescent proteins in the rhizosphere: new perspectives for studying microbial communities. MPMI, 2000, 13(11): 1170-1176.
13. B e r i n g e r J.E. R1 transfer in Rhizobium leguminosarum. J. Gen. Microbiol., 1974, 84: 188-198.
14. М а н и а т и с Т., Ф р и ч Э., С э м б р у к Дж. Методы генной инженерии. Молекулярное клонирование. М., 1984.
15. И б р а г и м о в а М.В., Р у м я н ц е в а М.Л., О н и щ у к О.П. и др. Симбиоз клубеньковых бактерий Sinorhizobiummeliloti с люцерной Medicagosativaв условиях засоления. Микробиология, 2006, 75(1): 94-100.
16. A l l e n O.N. Experiments in soil bacteriology. Burgess Publishing Co., Minneapolis, 1959: 54.
17. Ш а м ш у р и н А.А., К р и м е р М.З. Физико-химические свойства пестицидов: Справочник. М., 1976.
18. P o o l e P.S., S c h o f i e l d N.A., R e i d C.J. e.a. Identification of chromosomal genes located downstream of dctD that affect the requirement for calcium and the lipopolysaccharide layer of Rhizobium leguminosarum. Microbiology, 1994, 140(10): 2797-2809.
19. З а т о в с к а я Т.В., У ш а к о в К.В., Ю р г е л ь С.Н. и др. Получение Tn5-мутантов Rhizobiummeliloti с измененным составом липополисахаридов и анализ их симбиотических свойств. Генетика, 1998, 34(6): 742-748.
20. О н и щ у к О.П., Ш а р ы п о в а Л.А., К у р ч а к О.Н. и др. Выявление генов Sinorhizobiummeliloti, влияющих на синтез поверхностных полисахаридов и конкурентоспособность. Генетика, 2005, 41(12): 1617-1623.
21. C l o v e r R.H., K i e b e r J., S i g n e r E.R. Lipopolysaccharide mutants of Rhizobium meliloti are not defective in symbiosis. J. Bacteriol., 1989, 171(7): 3961-3967.
22. M e a d e H.M., L o n g S.R., R u v k u n G.B. e.a. Physical and genetic characterization of symbiotic and auxotrophic mutants of Rhizobium meliloti induced by transposon mutagenesis. J. Bacteriol., 1982, 149(1): 114-122.
23. H i r o a k i I., N o g i m a H., O k a y a m a H. High efficiency transformation of Escherichia coli with plasmids. Gene, 1990, 96: 23-28.
24. Ф е д о р о в С.Н., С и м а р о в Б.В. Получение мутантов с измененными симбиотическими свойствами у Rhizobiummeliloti под действием УФ-лучей. С.-х. биол., 1987, 9: 44-49.
25. O n i s h c h u k O.P., S h a r y p o v a L.A., S i m a r o v B.V. Isolation and characterization of the Rhizobium meliloti Tn5-mutants with impaired nodulation competitiveness. Plant and Soil, 1994, 167: 267-274.
26. О н и щ у к О.П., К у р ч а к О.Н., Ш а р ы п о в а Л.А. и др. Анализ различных типов конкурентоспособности у Tn5-мутантов клубеньковых бактерий люцерны (Sinorhizobiummeliloti). Генетика, 2001, 37(11): 1507-1512.
27. N o r r a n d e r J., K e m p e T., M e s s i n g J. Construction of improved M13 vectors using oligonucleotide-directed mutagenesis. Gene, 1983, 26: 101-106.
28. S u t c l i f f e J.G. Complete nucleotide sequence of the Escherichia coli plasmid pBR322. Cold Spring Harbor Symp. Quant. Biol., 1979, 43: 77-90.
29. Ю р г е л ь С.Н., Ш а р ы п о в а Л.А., С ы р ц о в а Л.А. и др. Дыхательная активность и симбиотическая эффективность у клубеньковых бактерий Rhizobiummeliloti. Микробиология, 1996, 86(4): 517-521.
30. A k a g a w a - M a t s u s h i t a M., M a t s u o p M., K o g a Y. e.a. Alteromonas atlantica sp. nov. and Alteromonas carrageenovora sp. nov., bacteria that decompose algal polysaccharides. Int. J. Syst. Bacteriol., 1992, 42: 621-627.
31. B a r b e y r o n T., P o t i n P., R i c h a r d C. e.a. Arylsulfatase from Alteromonas carrageenovora. Microbiology, 1995, 141: 2897-2904.
32. K i m J.-H., B y u n D.-S., G o d b e r J.S. e.a. Purification and characterization of arylsulfatase from Sphingomonas sp. AS6330. Appl. Microbiol. Biotechnol., 2004, 63: 553-559.
33. H a n s o n S.R., B e s t M.D., W o n g C.H. Sulfatases: structure, mechanism, biological activity, inhibition, and synthetic utility. Angew. Chem. Int. Ed. Engl., 2004, 43: 5736-5763.
34. M i e c h C., D i e r k s T., S e l m e r T. e.a. Arylsulfatase from Klebsiella pneumoniae carries a formylglycine generated from a serine. J. Biol. Chem., 1998, 273: 4835-4837.
35. H e n d e r s o n M.J., M i l a z z o F.H. Arylsulfatase in Salmonella typhimurium: detection and influence of carbon source and tyramine on its synthesis. J. Bacteriol., 1979, 139: 80-87.
36. B e i l S., K e h r l i H., J a m e s P. e.a. Purification and characterization of the arylsulfatase synthesized by Pseudomonas aerogenosa PAO during growth in sulfate-free medium and cloning of the arylsulfatase gene (atsA). Eur. J. Biochem., 1995, 229: 385-394.
37. H o f f m a n J.A., B a d g e r J.L., Z h a n g Y. e.a. Escherichia coli K1 aslA contributes to invasion of brain microvascular endothelial cells in vitro and in vivo. Infect. Immun., 2000, 68: 5062-5067.
38. F i t z g e r a l d J.W. Sulfate ester formation and hydrolysis: a potentially important yet often ignored aspect of the sulfur cycle of aerobic soils. Bacteriol. Rev., 1976, 40: 698-721.
39. C r o s s m a n L.C., J o h n s t o n A.W., T h o m s o n N.R. e.a. The genome of Rhizobium leguminosarum has recognizable core and accessory components. Genome Biol., 2006, 7(4): R34.
40. S l a t e r S.C., G o l d m a n B.S., G o o d n e r B. e.a. Genome sequences of three agrobacterium biovars help elucidate the evolution of multichromosome genomes in bacteria. J. Bacteriol., 2009, 191(8): 2501-2511.
41. O s t e r a s M., B o n c o m p a g n i E., V i n c e n t N. e.a. Presence of a gene encoding choline sulfatase in Sinorhizobium meliloti bet operon: choline-O-sulfate is metabolized into glycine betaine. PNAS, 1998, 95: 11394-11399.
42. П и щ и к В.Н., П р о в о р о в Н.А., В о р о б ь е в Н.И. и др. Взаимодействие растений с ассоциативными бактериями при загрязнении почвы тяжелыми металлами. Микробиология, 2009, 78(6): 826-835.
43. А н д р о н о в Е.Е., П е т р о в а С.Н., Ч и ж е в с к а я Е.П. и др. Влияние внесения генетически модифицированного штамма Sinorhizobiummeliloti ACH-5 на структуру почвенного сообщества микроорганизмов. Микробиология, 2009, 78(4): 474-482.
USE OF SITE-DIRECTED MUTAGENESIS TO STUDY THE FUNCTIONS OF GENE SMb20332 IN THE NODULE BACTERIA Sinorhizobium meliloti
E.P. Chizhevskaya, O.P. Onishchuk, E.E. Andronov, B.V. Simarov
The mutant for gene SMb20332 derived from alfalfa nodule bacteria Sinorhizobium meliloti type strain 1021 has been constructed by the use of site-directed mutagenesis. This gene is located on the megaplasmid-2 and possibly encodes for arylsulfatease according to the results of complete genome sequence of strain 1021 S. meliloti. Symbiotic properties, salt resistance and growth on the diagnostic media for mutant ACH-5 have been tested. It was shown that arylsulfatases from nodule bacteria resemble the arylsulfatases from marine bacteria which are able for desulphation agaropectin. Mutation in gene SMb20332 does not effect significantly the salt resistance of strain 1021 S. meliloti as well as its efficiency and competitiveness in symbiosis with Medicago sativa (cv. Vega). It was firstly shown that mutation in the gene SMb20332 of strain 1021 S. meliloti causes the change of surface polysaccharides and loose of resistance to the herbicide 2M-4HM (inhibitor of the bacterial respiratory activity).
Keywords: Sinorhizobium meliloti, arylsulphatase, production of polysaccharides, herbicide resistance, salt tolerance, competitiveness, symbiotic effectiveness.
ГНУ Всероссийский НИИ сельскохозяйственной |
Поступила в редакцию |




