doi: 10.15389/agrobiology.2022.2.328rus
УДК 636.2:636.084.1:636.085.57
Исследования выполнены при поддержке РНФ (проект № 20-16-00088).
АДАПТАЦИОННЫЕ ПРОЦЕССЫ В ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЕ ПРИ ВВЕДЕНИИ УЛЬТРАДИСПЕРСНЫХ ЧАСТИЦ ЖЕЛЕЗА В ЖИРОВЫЕ РАЦИОНЫ КРУПНОГО РОГАТОГО СКОТА
Е.В. ШЕЙДА1, 2 ✉, С.В. ЛЕБЕДЕВ2, С.А. МИРОШНИКОВ1, 2, В.В. ГРЕЧКИНА2, 3, О.В. ШОШИНА2
Жиры как кормовое средство служат концентрированными источниками энергии. Включение жировых компонентов в рационы сельскохозяйственных животных экономически целесообразно и эффективно. Однако некоторые исследования указывают на снижение переваримости питательных веществ рационов в присутствии жиров. Для повышения доступности питательных веществ рационов необходимо включение в комбикорма дополнительных компонентов, в частности ультрадисперсных частиц. В настоящей работе мы впервые установили влияние ультрадисперсного препарата железа на панкреатическую секрецию при дополнительном включении в рацион телят подсолнечного и соевого масел. Отмечено повышение ферментативной активности поджелудочного сока, а также переваримости питательных компонентов корма. Цель исследования — оценить возможность использования ультрадисперсных частиц железа в качестве модуляторов активности обменных процессов при введении растительных жиров (подсолнечное и соевое масла) в рацион телят. Опыты in vivo проводили с октября 2019 года по октябрь 2020 года в ФНЦ биологических систем и агротехнологий РАНна телятах (Bostaurustaurus) казахской белоголовой породы (по 4 гол. в группе, возраст 8 мес, средняя живая масса 120-130 кг). Применяли метод латинского квадрата 4×4, повторность эксперимента 5-кратная. Телята контрольной группы получали стандартный рацион (СР). В СР животных I группы дополнительно вводили ультрадисперсные частицы (УДЧ) Fe, II группы — подсолнечное масло, III группы — подсолнечное масло + УДЧ Fe, IV группы — соевое масло, V группы — соевое масло + УДЧ Fe. Масла вводили в расчете 3 % от сухого вещества рациона посредством замены на них концентратной части рациона. УДЧ (d = 90 нм, Z-потенциал 7,7±0,5 мВ) содержали 99,8 % Fe. Перед включением в рацион их диспергировали в физиологическом растворе, после чего замешивали с концентрированной смесью рациона в дозе 2,2 мг/гол. Для изучения внешнесекреторной функции поджелудочной железы проводили операцию по наложению дуоденального анастомоза. Панкреатический сок и химус собирали в течение 8 ч с интервалом 60 мин. Определяли активность амилазы, протеаз, липазы. Учитывали содержание NO-метаболитов в плазме крови, а также активность трипсина. Переваримость корма оценивали в течении 7 сут в балансовых опытах по количеству потребленного животными корма, несъеденных остатков и выделенного кала. Коэффициент переваримости (КП) рассчитывали, как отношение количества переваренных питательных веществ к поступившим в организм. В кале и кормах анализировали содержание питательных веществ: сухого вещества, сырого протеина, жира и золы. Кровь для оценки морфологических и биохимических показателей брали утром натощак на 7-е сут эксперимента. Включение в жировые рационы телят УДЧ Fe способствовало достоверному (р ≤ 0,05) увеличению переваримости сырого жира, органического вещества и безазотистых экстрактивных веществ, тогда как переваримость сырой клетчатки и сырого протеина снижалась. Дополнительное обогащение рационов УДЧ Fe и жировыми компонентами оказывало стимулирующее влияние на панкреатическую секрецию, приводя к увеличению количества продуцируемого сока. Под влиянием УДЧ Fe избирательно изменялась активность пищеварительных ферментов поджелудочной железы. При включении в контрольный рацион УДЧ Fe наблюдалось повышение активности липазы на 35,7 %, кишечных протеаз — на 43,1 % на фоне уменьшения амилолитической активности на 28,8 %. Введение УДЧ Fe в рационы, включающие подсолнечное и соевое масла, снижало ферментативную активность поджелудочной железы относительно контроля: в III группе усилилась активность липазы и кишечных протеаз соответственно на 12,1 и 16,7 % (р ≤ 0,05), в V группе — на 133,2 и 38,4 % (р ≤ 0,05). Дополнительное введение в рацион телят жировых компонентов самостоятельно и в комплексе с УДЧ привело к повышению содержания NO-метаболитов во всех опытных группах относительно контрольного показателя. При замене контрольного рациона на жировые отмечали повышение содержания трипсина во II группе на 106,6 % (р ≤ 0,05), в IV груп-пе — на 130,9 % (р ≤ 0,05), ышвведение УДЧ Fe способствовало снижению этого показателя. При морфологическом анализе было зафиксировано статистически значимое (р ≤ 0,05) повышение содержания гемоглобина в крови телят из опытных групп: в I группе — на 9,7 %, во II — на 31,2 %, в III — на 41,9 %, в IV — на 28,0 %, в V — на 30,1 %. Биохимический анализ крови показал, что все изучаемые параметры находились в пределах допустимых физиологических норм, однако следует отметить, что УДЧ Fe оказывали стимулирующее воздействие на белковый, жировой и угле-водный обмены в организме. Отмечено достоверное повышение коэффициента де Ритиса в группах, получавших УДЧ Fe на фоне жировых рационов: в III группе он составил 3,98, в V группе — 4,1 (р ≤ 0,05). Билирубиновый индекс относительно контроля повышался на 17,8 % (р ≤ 0,05) в I группе и на 5,5 % (р ≤ 0,05) — в IV группе, во всех других группах значения БИ были ниже, чем в контроле.
Ключевые слова: ультрадисперсные частицы, железо, морфология крови, биохимия крови, поджелудочная железа, ферменты, панкреатический сок, химус, крупный рогатый скот, жиры, подсолнечное масло, соевое масло.
В настоящее время при производстве высококачественной говядины жир превратился из простой добавки в ценный высокоэнергетический заменитель злаков, источник энергии и модификатор клеточного метаболизма (1). Жиры как кормовое средство служат концентрированными источниками энергии, содержат и транспортируют жирорастворимые витамины, обеспечивают организм незаменимыми жирными кислотами, а также придают корму определенные ароматические, вкусовые качества и структуру. Добавление жировых компонентов в состав полнокомпонентных рационов для сельскохозяйственных животных экономически целесообразно и эффективно (2, 3).
Включение растительных масел (кокосовое, пальмовое, соевое, подсолнечное, льняное и масло канолы) в рационы жвачных снижало продукцию кишечного метана in vitro на 40,55-48,58 %. При этом оно не влияло на pH рубца, количество микробного белка, усвояемость сухого и органического вещества (4). Добавление в рацион крупного рогатого скота (КРС) подсолнечного масла привело к снижению численности простейших, уменьшению метаногенеза и концентрации аммиачного азота, улучшению производства микробной биомассы и пропионовой кислоты в рубце (5). Однако некоторые исследователи указывают на снижение переваримости питательных веществ в присутствии жиров (6, 7). Отмечено снижение перевариваемости нейтрально-детергентных волокон в рубце из-за добавления жира, при этом эффективность синтеза микробного белка увеличивалась, а обилие простейших имело тенденцию к уменьшению (8). Жировые добавки приводили к статистически значимому (р ≤ 0,01) снижению переваримости органического вещества и нейтрально-детергентной клетчатки в кишечнике молодняка КРС на откорме (9).
Для повышения эффективности кормовых средств рассматриваются и изучаются такие компоненты рационов, как добавки минеральных веществ, в частности нанопорошки металлов (10-12). В настоящее время достаточно активно изучается влияние ультрадисперсных частиц металлов как самостоятельных добавок, а также в комплексе с другими компонентами корма на обменные процессы в желудочно-кишечном тракте, ферментативную активность пищеварительных желез и состав микробиома. Показано эффективное использование таких добавок для уменьшения побочных эффектов, улучшения биодоступности питательных веществ и увеличения продуктивных качеств (13-15).
В представленной работе мы впервые установили влияние ультрадисперсного препарата железа на панкреатическую секрецию при дополнительном включении в рацион телят подсолнечного и соевого масел. Отмечено повышение ферментативной активности поджелудочного сока, а также переваримости питательных компонентов корма.
Цель исследования — оценить возможность использования ультрадисперсных частиц железа в качестве модуляторов активности обменных процессов при введении растительных жиров (подсолнечное и соевое масла) в рацион телят.
Методика. Опыты in vivo проводили с октября 2019 года по октябрь 2020 года в ФНЦ биологических систем и агротехнологий РАНна телятах (Bos taurus taurus) казахской белоголовой породы. Формировали группы по четыре животных (возраст 8 мес, средняя живая массой 120-130 кг). Эксперимент выполняли по схеме латинского квадрата 4×4 в пяти повторностях.
Обслуживание животных и исследования проводили в соответствии с инструкциями и рекомендациями Russian Regulations, 1987 (Order No. 755 on 12.08.1977 the USSR Ministry of Health) и The Guide for Care and Use of Laboratory Animals (National Academy Press Washington, D.C. 1996). Были предприняты все меры, чтобы свести к минимуму страдания животных и сократить число отбираемых образцов. Животные содержались в отдельных метаболических клетках (1,0×2,2 м) в помещении с оптимальной температурой и влажностью (в течение эксперимента температура окружающей среды поддерживалась между 23 и 25 °C), доступ к воде был свободным.
Телята контрольной группы получали стандартный рацион (СР), включавший сено разнотравное (2 кг), смесь концентратов (1,5 кг), силос кукурузный (5 кг), солому пшеничную (1 кг), патоку кормовую (0,1 кг), соль поваренную (0,04 кг), витаминно-минеральный премикс. В стандартный рацион животных I группы дополнительно вводили ультрадисперсные частицы (УДЧ) Fe, II группы — подсолнечное масло, III группы — подсолнечное масло + УДЧ Fe, IV группы — соевое масло, V группы — соевое масло + УДЧ Fe. Масла вводили в расчете 3 % от сухого вещества рациона посредством замены концентратной части рациона. Животных кормили два раза в сутки (утром и вечером) в равных долях. Рационы были сформированы по потребности в питательных веществах и энергии, но различались по жирнокислотному составу вводимых растительных жиров (16, 17).
Ультрадисперсные частицы железа были получены методом электрического взрыва проводника в атмосфере аргона («Передовые порошковые технологии», Россия). УДЧ (d = 90 нм, Z-потенциал 7,7±0,5 мВ) содержали 99,8 % Fe. Перед включением в рацион их диспергировали в физиологическом растворе с помощью УЗДН-2Т («НПП Академприбор», Россия) (35 кГц, 300 Вт, 10 мкА, 30 мин). Животные получали УДЧ Fe после замешивания с концентрированной смесью рациона в дозе 2,2 мг/гол.
Для изучения внешнесекреторной функции поджелудочной железы выполняли операцию по наложению дуоденального анастомоза (18).
Исследования проводили после 16-часовой выдержки натощак. Панкреатический сок и химус собирали в течение 8 ч с интервалом 60 мин. После взятия первой пробы животных кормили и продолжали собирать сок и химус, количество сока и ферментативную активность сока и химуса определяли in cito.
Активность амилазы измеряли по Smith-Roe в модификации для высоких значений показателя (19), протеаз — по гидролизу казеина, очищенного по Гаммерстену, при калориметрическом контроле (λ = 450 нм) (20), активность липазы, концентрацию общего белка, фосфора, кальция и a-аммилазы — на автоматическом биохимическом анализаторе СS-T240 («DIRUI Industrial Co., Ltd», Китай) с использованием коммерческих биохимических наборов для ветеринарии (ЗАО «ДИАКОН-ДС», Россия) (21).
Содержание NO-метаболитов в плазме крови определяли спектрофотометрическим методом с реактивом Грисса, используя микропланшетный анализатор Infinite PRO F200 («Tecan Austria GmbH», Австрия) (при λ = 540 нм) (22).
Активность трипсина в плазме крови определяли на автоматическом биохимическом анализаторе СS-T240 («DIRUI Industrial Co., Ltd», Китай), в качестве субстрата использовали гидрохлорид натрий-бензоил-DL-аргинин-4(р)-нитроанилида (БАПНА).
Переваримость корма оценивали в течение 7 сут в балансовых опытах, учитывали количество потребленного животными корма, несъеденные остатки, количество выделенного кала. Коэффициент переваримости (КП) рассчитывали, как отношение переваренных питательных веществ к принятым. В кале и кормах после замораживания, высушивания и гомогенизации анализировали содержание сухого вещества, органического вещества, сырого протеина, сырого жира, безазотистых экстрактивных веществ (БЭВ) и золы в соответствии с рекомендациями Association of Official Agricultural Chemists (23). Переваримость оценивали по S. Hashemi с соавт. (24).
Кровь для оценки морфологических и биохимических показателей брали утром натощак на 7-е сут эксперимента из яремной вены в вакуумные пробирки с активатором свертывания (тромбин). Исследования проводили на автоматическом анализаторе СS-T240 («DIRUI Industrial Co., Ltd», Китай) с использованием коммерческих наборов для ветеринарии («ДиаВетТест», Россия).
Статистический анализ выполняли с использованием методик ANOVA в программном пакете Statistica 10.0 («StatSoft, Inc.», США) и программе Microsoft Excel. Представлены средние значения (M) и стандартные ошибки средних (±SEM). Достоверность различий сравниваемых показателей определяли по t-критерию Стьюдента. Различия считали статистически значимыми при p < 0,05.
Результаты. Состав и показатели качества рационов, которые скармливали подопытным телятам, представлены в таблице 1. Разница по содержанию сырого жира, сырого протеина и обменной энергии была достаточно значимой — соответственно 45,5; 8,3 и 12,9 %.
Основная задача при производстве высококачественной говядины — обеспечение животных необходимыми питательными веществами для удовлетворения метаболических потребностей и повышения продуктивности. Однако зерновые культуры, традиционные для рационов КРС, отрицательно влияют на содержание сухого вещества и подавляют переваривание клетчатки (25, 26). Использование жиров в рационах имеет важное значение в кормлении сельскохозяйственных животных. Недостаток жиров приводит к задержке роста, нарушению воспроизводительной функции, снижению продуктивности и ухудшению качества продукции. При этом наличие в рационах большого количества жиров создает нагрузку на систему пищеварения в целом, особенно у КРС. При насыщении рационов жирами изменяется активность пищеварительных ферментов, в результате чего сложные компоненты пищи недостаточно хорошо расщепляются и, как следствие, плохо усваиваются (27). Пищевой жир, который не поддается биолизу и биогидрированию с участием микроорганизмов рубца, но переваривается в нижних отделах пищеварительного тракта, известен как обходной жир, или жир, защищенный рубцом (инертный жир) (28). Введение жиров в рацион КРС, выпасаемого на пастбищах, увеличивает производство мясной и молочной продукции. Однако повышение количества жиров и жирных кислот тормозит процесс переваривания клетчатки в рубце и снижает переваривание органических веществ в передней части желудка (29).
В нашей работе при введении в рационы подсолнечного и соевого масла переваримость сырого жира снижалась соответственно на 38,2 и 10,9 % (р ≤ 0,05) относительно контроля (табл. 2). Включение в контрольный рацион телят УДЧ Fe способствовало повышению переваримости органического вещества на 9,6 % (р ≤ 0,05), сырого жира на 2,2 % (р ≤ 0,05), безазотистых экстрактивных веществ — на 9 % (р ≤ 0,05). Z. Khan с соавт. (30) установили, что при кормлении телят рационом с высоким содержанием железа снижались среднесуточный прирост живой массы, потребление сухого вещества и переваримость питательных веществ корма.
Присутствие в рационах УДЧ Fe способствовало статистически значимому увеличению переваримости сырого жира: в III группе относительно II — на 39,6 % (р ≤ 0,05), в V группе относительно IV — на 17,9 % (р ≤ 0,05). Похожая тенденция наблюдалась и в отношении переваримости органического вещества и БЭВ. Также следует отметить, что включение УДЧ Fe снижало переваримость сырой клетчатки и сырого протеина в I, III и V группах.
Дополнительное обогащение рационов минеральными компонентами и изменение количественного и качественного состава корма существенно влияют на панкреатическую секрецию и активность пищеварительных ферментов (31-34). Структура и состав рациона, а также объем и кратность кормления оказывают регулирующее воздействие на пищеварительные функции, что обеспечивается рефлекторно и гуморально.
В первый час до кормления животных панкреатическая секреция во всех группах была значительно ниже, чем после кормления. Секреция увеличивалась в рефлекторную и желудочную фазы, а затем снижалась в кишечную фазу, в период 360-480 мин после начала измерений.
При замене контрольного комбикорма на опытные образцы с УДЧ Fe увеличивалось количество продуцируемого поджелудочного сока, что свидетельствует о повышении нагрузки на поджелудочную железу при таких рационах (табл. 3). Так, при использовании контрольного рациона с УДЧ Fe количество панкреатического сока за все время опыта увеличивалось на 40,1 % (р ≤ 0,05). При добавлении к рациону подсолнечного масла продукция панкреатического сока уменьшалась на 19,8 % (р ≤ 0,05), соевого масла — на 8,8 % (р ≤ 0,05) относительно контроля. Включение в жировые рационы УДЧ Fe стимулировало секрецию сока поджелудочной железы в III группе на 149,0 % относительно показателя во II, а в V группе — на 127,6 % относительно IV (р ≤ 0,05).
Введение дополнительных ингредиентов и изменение качественного состава рациона приводят к избирательному изменению активности пищеварительных ферментов (35-38). При включении в контрольный рацион УДЧ Fe наблюдали достоверное повышение активности липазы на 35,7 % (р ≤ 0,05), кишечных протеаз — на 43,1 % на фоне снижения амилолитической активности на 35,0 % (р ≤ 0,05). В I группе содержание фосфора в панкреатическом соке снижалось на 14,3 % (р ≤ 0,05) при увеличении количества Са на 5,6 % (р ≤ 0,05) относительно контроля (табл. 4).
Жировые рационы способствовали стимуляции активности фермента липазы и кишечных протеаз во II группе соответственно в 8,5 (р ≤ 0,05) и 1,9 раза (р ≤ 0,05), в IV — в 7,3 и 1,5 раза (р ≤ 0,05) относительно контроля. В присутствии жировых компонентов в рационах активность фермента амилазы снижалась.
Дополнительное введение УДЧ Fe в рационы, включающие подсолнечное и соевое масла, снижало ферментативную активность поджелудочной железы, то есть нагрузка на нее уменьшалась. Относительно контрольных животных, находящихся на стандартном рационе, в III группе достоверно повысилась активность фермента липазы и протеаз соответственно на 12,1 и 16,7 % (р ≤ 0,05), в V группе — на 133,2 и 38,4 % (р ≤ 0,05). Присутствие в рационе жировых компонентов увеличивало секрецию липазы, но уменьшало секрецию амилазы. Очевидно, что увеличение количества любого питательного вещества приводит к повышению выработки в поджелудочной железе пищеварительных ферментов, предназначенных для его переваривания.
Введение в рационы УДЧ Fe значительно снижало активность амилазы панкреатического сока в составе дуоденального химуса: при включении в рацион подсолнечного масла — на 16,0 %, соевого масла — на 66,0 % (р ≤ 0,05) (рис. 1).
Тенденция к снижению активности наблюдалась и в отношении кишечных протеаз. При введении УДЧ Fe на фоне контрольного рациона протеолитическая активность снижалась на 52,0 %, однако различия оказались недостоверными. Было отмечено статистически значимое снижение активности протеаз во II группе на 28,8 % (р ≤ 0,05), в III — на 62,9 % (р ≤ 0,05), в V — на 50,0 % (р ≤ 0,05), в IV группе показатель снизился на 3,4 %. Активность липазы в дуоденальном химусе повышалась у животных, получавших жировые рационы, однако введение в них УДЧ Fe приводило к снижению показателя. Так, в I группе липолитическая активность снижалась в 4,3 раза (р ≤ 0,05), в III — в 4,6 рааз (р ≤ 0,05), а в V — в 4,5 раза (р ≤ 0,05).
Активность метаболитов оксида азота в сыворотке крови опосредует целый каскад физиологических процессов, в том числе регуляцию сосуди-стого тонуса, плазменного и тромбоцитарного звеньев гемостаза, нейротрансмиссию и формирование иммунного ответа, торможение пролиферации гладкомышечных клеток и оказывает значительное влияние на процессы обмена в пищеварительном тракте (39). В нашем исследовании наблюдалось повышение содержания NO-метаболитов во всех опытных группах относительно контрольных значений (рис. 2).
Дополнительное вве-дение УДЧ Fe статистически значимо увеличивало содержание метаболитов оксида азота в I группе в 5,1 раза (р ≤ 0,05), в III — в 7,2 раза (р ≤ 0,05), в V — в 5,6 раза (р ≤ 0,05). Недостаточная продукция NO у животных из контрольной группы сопряжена с развитием нарушений в сердечно-сосудистой и других системах организма. При этом S.V. Rama Rao с соавт. (40) установили, что избыточная продукция NO, за счет которой обеспечивается антимикробный эффект при воспалении, может превратиться из звена адаптации в звено патогенеза и стать не менее опасным повреждающим фактором для организма, чем дефицит NO. Нарастание количества NO-метаболитов у телят в III опытной группе при включении в рацион УДЧ Fe свидетельствовало о компенсаторной реакции организма на изменяющийся липидный профиль рациона.
Показатели активности амилазы и липазы в сыворотке крови не всегда могут свидетельствовать о физиологическом напряжении функции поджелудочной железы при смене рационов, поскольку существует экстрапанкреатическая продукция этих ферментов. Трипсин — оптимальный маркер для выявления изменения в физиологическом состоянии поджелудочной железы, поскольку он специфичен для этого органа. Установлено, что поступление трипсина в кровь уменьшает выход ферментов с панкреатическим соком, а введение ингибитора трипсина, напротив, сопровождается увеличением секреции ферментов (41).
В наших опытах при введении в рацион УДЧ Fe во всех опытных группах в сыворотке крови повышалась активность трипсина, который играет критическую роль в инициации каскада активации пищеварительных ферментов в кишечнике (рис. 3). При введении УДЧ Fe на фоне контрольного рациона активность трипсина увеличивалась на 31,2 % (р ≤ 0,05). При замене контрольного рациона на жировые этот показатель повышался во II группе на 106,6 % (р ≤ 0,05), в IV группе на 130,9 % (р ≤ 0,05) относительно контроля. При включении в жировые рационы УДЧ Fe активность трипсина снижалась на 48,8 % (р ≤ 0,05) в III группе относительно II группы, в V группе — на 50,5 % (р ≤ 0,05) относительно IV группы.
Активация трипсина при протеолитическом расщеплении трипсиногена в поджелудочной железе может привести к ряду событий, которые вызывают панкреатическое самовосприятие. Одно из последствий аутосомно-рецессивного заболевания муковисцидоза — недостаточный транспорт трипсина и других пищеварительных ферментов из поджелудочной железы (42).
В результате анализа морфологических показателей крови телят установлено, что при замене контрольного рациона на опытный с введением соевого масла число лейкоцитов статистически значимо повышалось на 57,3 % (р ≤ 0,05) относительно контроля, но включение УДЧ Fe приводило к снижению показателя на 30,5 % относительно IV группы (р ≤ 0,05) (табл. 5).
Введение УДЧ Fe приводило к усилению всасывания железа из желудочно-кишечного тракта, улучшению синтеза железосодержащих метаболитов (в том числе гемоглобина), стимуляции эритропоэза (32). В экспериментах на 4-месячных телочках, получавших нанопорошок железа, число эритроцитов увеличилось на 19,6 %, а количество гемоглобина — на 17,1 % по отношению к контролю.
Мы установили, что включение как в контрольный, так и в жировые рационы УДЧ Fe снижало числа лимфоцитов и моноцитов. Число эритроцитов снижалось в I группе на 2,0 % относительно контроля, в III — на 9,0 % относительно II группы, в V группе — на 36,2 % (р ≤ 0,05) относительно IV группы.
Непосредственно участие железа в образовании гемоглобина способствовало повышению содержания гемоглобина в опытных группах при включении УДЧ Fe в рацион. Количество гемоглобина в крови у животных опытных групп статистически значимо (р ≤ 0,05) повышалось в I группе на 9,7 %, во II — на 31,2 %, в III — на 41,9 %, в IV — на 28,0 %, в V — на 30,1 % относительно показателя в контроле. Высокое содержание железа в геме делает гемоглобин идеальной молекулой для целевого извлечения железа во время эндогенного воздействия УДЧ. Значительное число эритроцитов и количества гемоглобина в крови животных свидетельствует о более интенсивных окислительно-восстановительных процессах в организме и соответствует более высоким показателям продуктивности (43).
Наибольшее количество тромбоцитов отмечали во II опытной группе (224×109/л), что оказалось выше контрольного показателя на 11,4 % (р ≤ 0,05). Конкуренция за гемовое железо между клетками крови и пулом экзогенных бактерий на фоне напряжения эритропоэза, а также низкая всасываемость железа эндотелием и увеличение моторики желудочно-кишечного тракта могли стать причиной некоторого роста показателя.
Количество форменных элементов лейкоцитарного звена крови телят в контрольной и опытной группах находилось в пределах физиологической нормы. В структуре самой лейкограммы, отражающей процентное соотношение разных популяций лейкоцитов, отклонений от нормы не регистрировали. Индекс соотношения лимфоцитов и моноцитов (ИСЛМ), который характеризует взаимоотношение аффекторного и эффекторного звеньев иммунного ответа, показал, что ИСЛМ преобладал во II группе и далее уменьшался в ряду III группа > V группа > IV группа > контроль > I группа (см. табл. 5).
Значения морфологических показателей крови у телят из опытных групп соответствовали более высокой метаболической активности.
Изменение биохимических показателей отражает адаптацию всех систем организма, в том числе пищеварения и общего гомеостаза, к смене условий кормления (43). В наших опытах введение в рацион телят УДЧ Fe стимулировало белковый обмен (табл. 6). Увеличение количества общего белка в сыворотке крови свидетельствовало о лучшем усвоении азота корма, чему способствовало и повышение ферментативной активности (29). При замене контрольного рациона на жировые этот показатель возрастал во II группе на 20,6 % (р ≤ 0,05), в IV группе — на 38,0 %. Дополнительное введение УДЧ Fe повышало содержание общего белка в I группе на 5,6 %, в III на 100,8 % и в V группе на 114,6 % относительно контроля.
Тенденция к повышению наблюдалась и в отношении альбумина (см. табл. 6). Его количество статистически значимо (р ≤ 0,05) повышалось на 24,1 % во II и на 44,8 % в IV группе. Содержание мочевины в опытных группах превышало контроль на значения от 31,2 % (в I группе) до 59,4 % (р ≤ 0,05) (во II группе).
Несвязанное железо также служит индуктором перекисного окисления липидов и перекисной деструкции белков. Металлы в микрочастицах имеют низкую степень высвобождения и скорость усвоения, тем самым исключают токсическое воздействие на организм и кишечную микрофлору (12). Изменение жирового обмена оценивали по содержанию триглицеридов и холестерина в сыворотке крови. При включении в рацион УДЧ Fe количество триглицеридов повышалось на 17,2 % (р ≤ 0,05) (см. табл. 6).
Растительные масла в рационе животных оказывают значительное влияние на липидный профиль крови (45). В нашем опыте при замене контрольного рациона на жировые содержание триглицеридов повышалось во II группе на 27,6 %, в IV группе — на 55,2 % (р ≤ 0,05). Дополнительное введение в жировые рационы УДЧ Fe способствовало достоверному снижению количества триглицеридов в сыворотке крови телят из III группы на 68,9 % (р ≤ 0,05), в V группе на 75,9 % (р ≤ 0,05) относительно контрольных значений. Аналогичную тенденцию отмечали и в отношении содержания холестерина (см. табл. 6).
Воздействие УДЧ Fe на углеводный обмен у животных оценивали по содержанию глюкозы в сыворотке крови. Показатель статистически значимо (р ≤ 0,05) увеличивался в I и III группах на 44,9 %, в V — на 53,1 % относительно контроля (см. табл. 6).
Включение в рационы УДЧ Fe способствовало увеличению содержания железа (р ≤ 0,05), однако количество магния, кальция и фосфора недостоверно снижалось, за исключением III группы. При замене контрольного рациона на жировые содержание P повышалось во II группе на 31,8 %, в IV — на 9,1 %, содержание Са во II группе — на 22,9 %, в IV — на 10,6 %, содержание Mg во II группе — на 41,8 %, в IV — на 37,7 % относительно контроля.
Включение в контрольный и жировые рационы УДЧ Fe способствовало как стимуляции, так и торможению некоторых процессов. Увеличение соотношения АсАТ/АлАТ (коэффициент де Ритиса) могло свидетельствовать о хронических процессах, связанных с паренхиматозным поражением печени вследствие интоксикации организма тяжелыми металлами. В коэффициенте де Ритиса АсАТ отражает активность реакций центрального звена метаболизма, регулирует поступление субстратов в цикл Кребса с их последующим аэробным окислением, выполняет детоксицирующую функцию по отношению к аммиаку, включая его в цикл синтеза мочевины, а также обеспечивает восстановление в тканях содержания аспартата, снижающегося при дисбалансе аминокислот и гипоксии (43, 45). Чем выше содержание АлАТ, тем ниже коэффициент де Ритиса.
Содержание АлАТ повышалось в группах, получавших жировые рационы: во II группе — на 33,6 %, в IV — на 21,0 %, однако эти изменения не были статистически значимы. Дополнительное введение УДЧ Fe способствовало снижению количества АлАТ в I группе на 5,5 % (р ≤ 0,05) относительно контроля, в III — на 16,4 % (р ≤ 0,05) относительно II группы, находящейся на том же рационе, но без добавления железа. Коэффициент де Ритиса был достоверно (р ≤ 0,05) выше в группах, получавших УДЧ Fe на фоне жировых рационов (см. табл. 6).
Также был рассчитан билирубиновый индекс (БИ) крови, который характеризует выделительную функцию печени и показывает степень токсичности УДЧ железа. БИ повышался на 17,8 % (р ≤ 0,05) в I группе и на 5,5 % (р ≤ 0,05) в IV группе относительно контроля, во всех других опытных группах значения БИ были ниже, чем в контроле.
Таким образом, включение в жировые рационы телят казахской белоголовой породы ультрадисперсных железа способствовало достоверному увеличению переваримости сырого жира, органического вещества и безазотистых экстрактивных веществ корма, тогда как переваримость сырой клетчатки и сырого протеина снижалась. Дополнительное обогащение рационов УДЧ Fe и жировыми компонентами оказывало стимулирующее влияние на панкреатическую секрецию, приводя к увеличению количества продуцируемого сока. Под влиянием УДЧ Fe избирательно изменялась активность пищеварительных ферментов поджелудочной железы. При включении в контрольный рацион УДЧ Fe наблюдалось повышение активности липазы на 35,7 %, кишечных протеаз — на 43,1 % на фоне снижения амилолитической активности на 28,8 %. Введение УДЧ Fe в жировые рационы снижало ферментативную активность поджелудочной железы относительно контрольной группы: в варианте с подсолнечным маслом активность липазы и кишечных протеаз усиливалась соответственно на 12,1 и 16,7 % (р ≤ 0,05), с соевым маслом — на 133,2 и 38,4 % (р ≤ 0,05). Дополнительное введение в рацион телят жировых компонентов отдельно и в комплексе с УДЧ приводило к повышению содержания NO-метаболитов во всех опытных группах относительно контрольного показателя. При добавлении к рационам подсолнечного и соевого масла отмечали повышение содержания трипсина соответственно на 106,6 % (р ≤ 0,05) и 130,9 %, введение УДЧ Fe способствовало снижению этого показателя. При морфологическом анализе было выявлено статистически значимое повышение содержания гемоглобина в крови у животных опытных групп на 9,7-41,9 % (р ≤ 0,05). Биохимический анализ крови показал, что все изучаемые параметры находились в пределах допустимых физиологических норм, однако УДЧ Fe оказывали стимулирующее воздействие на белковый, жировой и углеводный обмен. Было отмечено повышение коэффициента де Ритиса в группах, получавших УДЧ Fe на фоне рационов с подсолнечным и соевым маслом. Во всех других опытных группах значения билирубинового индекса были ниже, чем в контрольной группе, что указывает на достаточно низкую токсичность препарата ультрадисперсных частиц железа.
1. Состав и показатели качества рационов телят (Bos taurus taurus) казахской белоголовой породы в эксперименте (ФНЦ биологических систем и агротехнологий РАН, 2019-2020 годы) |
|||
Показатель |
Рацион |
||
стандартный |
с подсолнечным маслом |
с соевым маслом |
|
Состав рациона |
|||
Сено разнотравное, кг |
7,0 |
7,0 |
7,0 |
Концентраты, кг |
2,0 |
2,0 |
2,0 |
Масло подсолнечное, кг |
|
0,3 |
|
Масло соевое, кг |
|
|
0,3 |
Патока кормовая, кг |
0,6 |
0,6 |
0,6 |
Премикс ПК-60, кг |
0,06 |
0,06 |
0,06 |
Соль, кг |
0,02 |
0,02 |
0,2 |
УДЧ Fe, мг |
|
2,2 |
2,2 |
Питательность рациона |
|||
Сухое вещество, кг |
8,42 |
8,42 |
8,42 |
Сырая клетчатка, кг |
2,56 |
2,56 |
2,56 |
Сырой жир, кг |
0,244 |
0,355 |
0,355 |
Сырой протеин, кг |
0,72 |
0,66 |
0,66 |
БЭВ, кг |
5,4 |
5,0 |
5,0 |
Кальций, г |
42,2 |
42,6 |
43,2 |
Фосфор, г |
30,0 |
29,8 |
30,4 |
ОЭ, МДж |
63,0 |
71,1 |
71,1 |
Примечание. БЭВ — безазотистые экстрактивные вещества, ОЭ — обменная энергия. Состав витаминно-минерального премикса (на 1 кг концентрата): Mn — 48 мг, Zn — 36 мг, Fe — 60 мг, Cu —10 мг, Se — 0,24 мг, Со — 0,12 мг; витамин А — 2640 МЕ, витамин Д — 302 МЕ; витамин Е — 17 мг. |
|||
2. Коэффициенты переваримости питательных веществ (%) у телят (Bos taurus taurus) казахской белоголовой породы при скармливании рационов с растительными жирами и ультрадисперсными частицами железа (n = 4, M±SEM, ФНЦ биологических систем и агротехнологий РАН, 2019-2020 годы) |
||||||
Показатель |
Группа |
|||||
контроль |
I |
II |
III |
IV |
V |
|
Сухое вещество |
74,3±0,04 |
70,8±0,03* |
76,1±0,01* |
72,1±0,02* |
60,9±1,05 |
68,4±0,03* |
Органическое вещество |
87,8±0,30 |
96,2±0,40* |
83,83±0,21* |
98,7±0,42 |
59,4±0,03* |
95,8±0,05* |
Сырой протеин |
76,3±3,60 |
71,0±4,10 |
81,4±2,30* |
74,5±2,82 |
72,8±1,02 |
70,4±2,4 |
Сырой жир |
72,7±1,23 |
74,3±1,40* |
44,9±1,88* |
62,7±1,54* |
64,8±0,75* |
76,4±1,23* |
Сырая клетчатка |
37,4±0,18 |
36,6±0,20 |
45,5±0,08* |
37,5±0,12 |
43,7±0,45* |
36,2±1,12 |
БЭВ |
80,3±0,90 |
88,2±0,70* |
82,3±1,10 |
91,8±0,93* |
75,6±0,38* |
84,3±0,24* |
Примечание. БЭВ — безазотистые экстрактивные вещества. Описание групп см. в разделе «Методика». |
||||||
3. Количество панкреатического сока (мл), выделявшегося у телят (Bos taurus taurus) казахской белоголовой породы при скармливании рационов с растительными жирами и ультрадисперсными частицами железа (n = 4, M±SEM, ФНЦ биологических систем и агротехнологий РАН, 2019-2020 годы) |
||||||
Продолжительность эксперимента, мин |
Группа |
|||||
контроль |
I |
II |
III |
IV |
V |
|
0-60 |
32,0±2,81 |
48,0±3,24 |
28,0±1,72 |
109,0±19,02 |
18,0±2,43 |
88,0±12,23 |
60-120 |
66,0±3,42 |
76,0±3,61* |
48,0±3,51 |
138,0±6,52 |
78,0±6,72* |
141,0±5,64* |
120-180 |
67,0±5,32 |
80,0±5,12 |
58,0±4,62* |
228,0±4,11* |
72,0±7,12 |
218,5±14,64 |
180-240 |
59,0±4,71 |
84,0±6,04 |
50,0±5,03 |
156,5±1,93* |
62,0±5,83 |
184,5±3,83 |
240-300 |
55,5±5,20 |
88,0±4,22 |
48,0±3,72 |
142,5±9,92 |
5,0±4,31 |
168,5±11,31* |
300-360 |
59,5±4,11 |
91,0±2,81 |
47,0±3,30* |
77,0±5,02 |
48,0±5,31 |
81,5±18,04 |
360-420 |
67,0±6,31 |
90,0±4,52 |
38,0±5,43 |
23,0±6,23 |
46,0±3,82 |
39,0±6,40 |
420-480 |
51,5±4,70 |
84,0±3,02* |
50,0±5,22 |
40,0±2,90 |
42,0±3,22 |
28,0±3,91 |
0-480 |
457,5±37,83 |
641,0±32,40* |
367,0±55,42* |
914,0±55,53* |
417,0±45,22* |
949,0±75,81* |
Примечание. В первый час эксперимента показатели фиксировали натощак. Описание групп см. в разделе «Методика». |
||||||
4. Активность ферментов панкреатического сока у телят (Bos taurus taurus) казахской белоголовой породы при введении в рационы растительных жиров и ультрадисперсных частиц железа (n = 4, M±SEM, ФНЦ биологических систем и агротехнологий РАН, 2019-2020 годы) |
||||||
Показатель |
Группа |
|||||
контроль |
I |
II |
III |
IV |
V |
|
Липаза, Ед/л |
90,9±18,2 |
123,4±19,4* |
773,0±14,8* |
101,9±12,7* |
667,0±37,0* |
212,0±11,3* |
Амилаза, мг·мл-1·мин-1 |
5137,5±450,0 |
3337,5±330,0 |
2537,0±400,0 |
1698,4±330,0 |
1931,0±69,0 |
1456,0±34,0 |
Протеазы, мг·мл-1·мин-1 |
133,5±24,3 |
191,0±22,6 |
249,0±21,1* |
155,8±14,6* |
200,0±12,6* |
184,8±13,5* |
Общий белок, г/л |
0,46±0,12 |
0,48±0,16 |
0,18±0,01 |
0,38±0,01* |
0,33±0,01 |
0,41±0,020 |
Фосфор, моль/л |
0,14±0,02 |
0,12±0,03* |
0,03±0,00 |
0,10±0,00 |
0,08±0,01 |
0,10±0,010* |
Кальций, моль/л |
2,33±0,12 |
2,46±0,15* |
2,43±0,22* |
2,26±0,18 |
2,39±0,10* |
2,41±0,12 |
α-Амилаза, Ед/л |
416,0±4,8 |
536,0±6,2 |
578,0±11,5 |
559,1±6,7 |
767,0±13,8 |
758,0±16,8 |
Примечание. Описание групп см. в разделе «Методика». |
||||||
5. Морфологические показатели крови у телят (Bos taurus taurus) казахской белоголовой породы при введении в рационы растительных жиров и ультрадисперсных частиц железа (n = 4, M±SEM, ФНЦ биологических систем и агротехнологий РАН, 2019-2020 годы) |
||||||
Показатель |
Группа |
|||||
контроль |
I |
II |
III |
IV |
V |
|
Лейкоциты, ×109/л |
7,5±1,32 |
7,1±1,20 |
7,2±1,63 |
6,9±1,32 |
11,8±2,22* |
8,2±1,72* |
Лимфоциты, ×109/л |
2,9±0,83 |
2,6±0,63 |
4,1±1,21 |
3,2±1,13 |
5,5±1,21 |
4,3±1,32 |
Моноциты, ×109/л |
1,3±0,32 |
1,2±0,25 |
1,5±0,31 |
1,2±0,22 |
2,5±0,42 |
1,6±0,33 |
ИСЛМ |
2,21 |
2,13 |
2,72 |
2,61 |
2,22 |
2,50 |
Гранулоциты, ×109/л |
1,90±0,63 |
2,10±0,52 |
5,30±0,82* |
4,30±0,62* |
4,80±0,61 |
4,36±0,71 |
Эритроциты, ×1012/л |
5,08±2,91 |
4,98±2,22 |
5,37±3,13 |
4,88±2,50 |
7,62±1,81* |
4,86±1,61* |
Гемоглобин, г/л |
93,0±11,12 |
102,0±9,81* |
122,0±13,80* |
132,0±11,51* |
119,0±14,70* |
121,0±12,22* |
Гематокрит, % |
20,1±4,34 |
22,6±3,63 |
21,6±4,91 |
22,0±2,82 |
24,2±3,94 |
20,8±2,61 |
MCHC, г/л |
349±26,92 |
324±28,63 |
440±26,42 |
388±36,53 |
424±27,72 |
368±34,23 |
Тромбоциты, ×109/л |
201±19,81 |
212±14,32 |
224±17,71 |
216±18,22 |
220±21,70 |
206±14,63 |
Примечание. ИСЛМ — индекс соотношения лимфоцитов и моноцитов, MCHC — средняя концентрация гемоглобина в эритроците. Описание групп см. в разделе «Методика». |
||||||
6. Биохимические показатели крови у телят (Bos taurus taurus) казахской белоголовой породы при введении в рационы растительных жиров и ультрадисперсных частиц железа (n = 4, M±SEM, ФНЦ биологических систем и агротехнологий РАН, 2019-2020 годы) |
||||||
Показатель |
Группа |
|||||
контроль |
I |
II |
III |
IV |
V |
|
Общий белок, г/л |
72,05±3,98 |
76,05±2,82* |
86,85±4,51* |
144,70±4,81* |
99,43±6,98 |
154,50±4,54* |
Альбумин, г/л |
29,00±6,12 |
38,00±5,41 |
36,0±5,80* |
38,00±4,72 |
42,00±5,33* |
44,20±3,82 |
Глюкоза, ммоль/л |
3,41±0,87 |
4,94±0,55* |
3,52±0,63 |
4,94±0,46* |
4,21±0,74* |
5,22±0,41* |
Триглицериды, ммоль/л |
0,29±0,07 |
0,34±0,06* |
0,37±0,03 |
0,09±0,01* |
0,45±0,09* |
0,07±0,01* |
Холестерин, ммоль/л |
2,67±0,19 |
1,08±0,05* |
3,63±0,31* |
0,92±0,03 |
4,99±0,81* |
1,06±0,07 |
АлАТ, Ед/л |
23,80±4,31 |
22,50±3,81* |
31,80±5,12 |
26,60±2,71* |
28,80±5,11 |
26,30±2,91 |
АсАТ, Ед/л |
44,20±5,93 |
42,20±2,92* |
52,3±6,33 |
105,80±6,12* |
54,90±5,82 |
108,60±4,82 |
Коэффициент де Ритиса |
1,86 |
1,87* |
1,64 |
3,98* |
1,91* |
4,12* |
Билирубин общий, мкмоль/л |
2,43±0,07 |
2,61±0,08* |
3,25±0,08* |
2,16±0,12 |
3,67±0,09* |
1,94±0,22 |
Билирубин прямой, мкмоль/л |
1,11±0,13 |
1,01±0,11 |
1,72±0,18 |
1,05±0,02 |
1,59±0,16 |
1,84±0,06** |
Биллирубиновый индекс |
2,19 |
2,58* |
1,91* |
2,05* |
2,31* |
1,05 |
ЛДГ, Ед/л |
3049±56,05 |
3856±62,21* |
5272±64,31 |
3659±51,12* |
4098±63,70* |
3426±42,01* |
α-Амилаза, Ед/л |
415±23,11 |
712±30,22 |
471±63,12* |
358±16,21 |
423±21,91 |
346±11,62 |
Липаза, Ед/л |
17,30±3,42 |
18,00±2,20 |
16,80±1,21 |
8,00±0,63 |
28,40±3,91 |
8,60±0,52 |
Мочевина, ммоль/л |
3,20±0,72 |
4,20±0,63 |
5,10±0,91* |
5,00±0,91 |
4,60±0,92 |
4,40±0,55 |
Креатинин, мкмоль/л |
74,50±6,31 |
81,20±5,11 |
88,70±7,23* |
93,10±5,31 |
89,60±7,21 |
92,80±4,83 |
γ-ГТ, Ед/л |
18,30±2,60 |
23,20±3,21 |
32,40±4,11 |
24,00±2,12 |
23,60±3,13 |
21,00±2,32 |
Мочевая кислота, мкмоль/л |
15,50±3,22 |
16,00±2,82 |
18,90±4,32 |
21,20±3,82 |
16,10±3,92 |
19,80±3,61 |
Железо, мкмоль/л |
19,20±3,81 |
22,70±4,61* |
33,40±5,12* |
49,30±4,41* |
34,60±4,31 |
36,80±3,62* |
Магний, ммоль/л |
1,22±0,21 |
1,08±0,08 |
1,73±0,31 |
0,84±0,02 |
1,68±0,91 |
0,78±0,06 |
Кальций, ммоль/л |
2,45±1,12 |
2,68±1,21 |
3,01±1,22 |
2,60±0,58 |
2,71±0,83 |
2,32±0,12 |
Фосфор, ммоль/л |
1,54±0,04 |
2,04±0,08 |
2,03±0,06 |
1,97±0,09 |
1,68±0,62 |
1,44±0,23 |
Примечание. АлАТ — аланинаминотрансфераза, АсАТ — аспартатаминотрансфераза, ЛДГ — лактатдегидрогеназа, γ-ГТ — γ-глютамилтранспептидаза. Описание групп см. в разделе «Методика». |
||||||
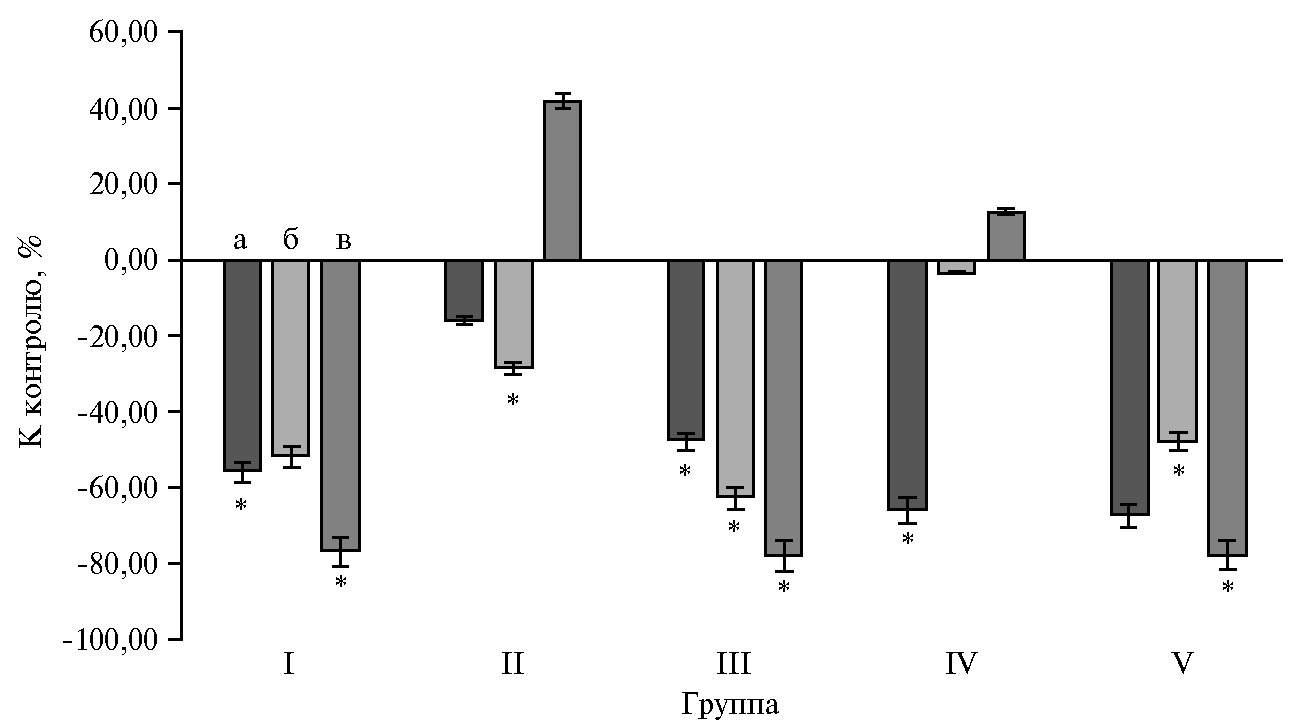
Рис. 1. Изменение активности пищеварительных ферментов панкреатического сока в составе дуоденального химуса у телят (Bos taurus taurus) казахской белоголовой породы относительно контроля при введении в рационы растительных жиров и ультрадисперсных частиц железа: а — амилаза, б — протеазы, в — липаза (n = 4, M±SEM, ФНЦ биологических систем и агротехнологий РАН, 2019-2020 годы). Описание групп см. в разделе «Методика».
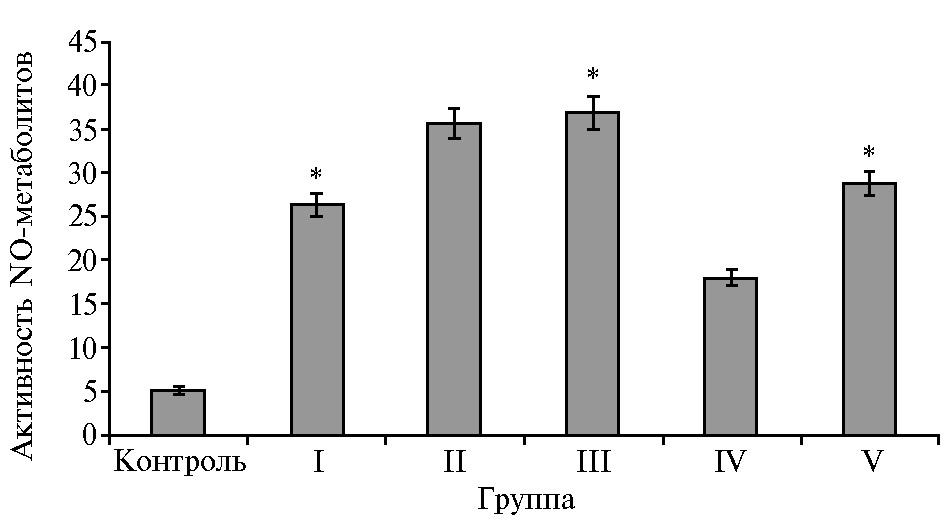
Рис. 2. Содержание NO-метаболитов в сыворотке крови у телят (Bos taurus taurus) казахской белоголовой породы при введении в рационы растительных жиров и ультрадисперсных частиц железа (n = 4, M±SEM, ФНЦ биологических систем и агротехнологий РАН, 2019-2020 годы). Описание групп см. в разделе «Методика».
* Различия с контрольной группой статистически значимы при р ≤ 0,05.
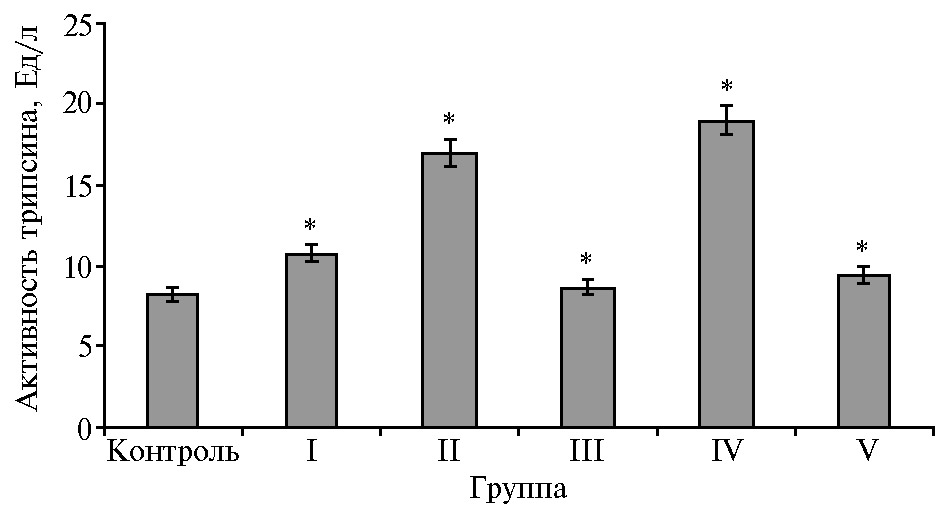
Рис. 3. Активность трипсина в сыворотке крови у телят (Bos taurus taurus) казахской белоголовой породы при введении в рационы растительных жиров и ультрадисперсных частиц железа (n = 4, M±SEM, ФНЦ биологических систем и агротехнологий РАН, 2019-2020 годы).
* Различия с контрольной группой статистически значимы при р ≤ 0,05.
ЛИТЕРАТУРА
- Зенова Н.Ю., Назарова А.А., Полищук С.Д. Влияние ультрадисперсного железа на рост и развитие крупного рогатого скота. Молочное и мясное скотоводство, 2010, 1: 30-32.
- Калагина Л.С. Клиническое значение определения показателей трипсина в сыворотке крови (обзор). Медицинский альманах, 2010, 1: 281-283.
- Вертипрахов В.Г., Егоров И.А., Андрианова Е.Н., Грозина А.А. Физиологические аспекты использования разных растительных масел в кормлении цыплят-бройлеров (Gallus gallus L.). Сельскохозяйственная биология, 2018, 53(4): 811-819 (doi: 10.15389/agrobiology.2020.6.1159rus).
- Hartanto R., Cai L., Yu J., Zhang N., Sun L., Qi D. Effects of supplementation with monensin and vegetable oils on in vitro enteric methane production and rumen fermentability of goats. Pakistan Journal of Agricultural Sciences, 2017, 54(3): 693-698 (doi: 10.21162/PAKJAS/17.4347).
- Santra A., Banerjee A., Das S.K. Еffect of vegetable oils on ciliate protozoa, methane yield, enzyme profile and rumen fermentation in vitro. Animal Nutrition and Feed Technology, 2013, 13(2): 181-193.
- Oldick B.S., Firkins J.L. Effects of degree of fat saturation on fiber digestion and microbial protein synthesis when diets are fed twelve times daily. Journal of Animal Science, 2000, 78(9): 2412-2420 (doi: 10.2527/2000.7892412x).
- Plascencia A., Mendoza G.D., Vásquez C., Zinn R.A. Relationship between body weight and level of fat supplementation on fatty acid digestion in feedlot cattle. Journal of Animal Science,2003, 81(11): 2653-2659 (doi: 10.2527/2003.81112653x).
- Левахин Ю.И., Нуржанов Б.С., Рязанов В.А., Джуламанов Е.Б. Изменения микробиоценоза рубца, крови и переваримость сухого вещества рациона при введении бычкам совместно c жировой добавкой ультрадисперсных частиц железа. Аграрный вестник Урала, 2019, 192(1): 53-59 (doi: 10.32417/1997-4868-2020-192-1-53-59).
- Калашников А.П., Фисинин В.И., Щеглов В.В., Клейменов Н.И. Нормы и рационы кормления сельскохозяйственных животных. М., 2003.
- Шейда Е.В., Русакова Е.А., Сипайлова О.Ю., Cизова Е.А., Лебедев С.В. Токсические эффекты ультрадисперсных форм металлов (Mo и MoO3) в эксперименте in vivo. Сельскохозяйственная биология, 2020, 55(6): 1171-1181 (doi: 10.15389/agrobiology.2020.6.1171rus).
- Gülşen N., Umucalilar H.D., Inal F., Hayirli A. Impacts of calcium addition and different oil types and levels on in vitro rumen fermentation and digestibility. Archives of Animal Nutrition, 2006, 60(6): 443-453 (doi: 10.1080/17450390600973634).
- Hassan S., Hassan F.U., Rehman M.S.U. Nano-particles of trace minerals in poultry nutrition: potential applications and future prospects. Biol. TraceElem. Res., 2020, 195(2): 591-612 (doi: 10.1007/s12011-019-01862-9).
- Фисинин В.И., Вертипрахов В.Г., Титов В.Ю., Грозина А.А. Динамика активности пищеварительных ферментов и содержание депонированного оксида азота в плазме крови петушков после кормления. Российский физиологический журнал имени И.М. Сеченова, 2018, 104(8): 976-983 (doi: 10.7868/S0869813918070080).
- Corring T. The adaptation of digestive to the diet: Its physiological significante. Reprod. Nutr. Develop., 1980, 20(4B): 1217-1235 (doi: 10.1051/rnd:19800713).
- Clary J., Mitchell Jr. G.E., Bradley N.W. Pancreatic amylase activity from ruminants fed different rations. Canadian Journal of Physiology and Pharmacology, 1969, 47(2): 161-164 (doi: 10.1139/y69-027).
- Duthie C.A., Troy S.M., Hyslop J.J., Ross D.W., Roehe R., Rooke J.A. The effect of dietary addition of nitrate or increase in lipid concentrations, alone or in combination, on performance and methane emissions of beef cattle. Animal, 2018, 12(2): 280-287 (doi: 10.1017/S175173111700146X).
- Eastridge M.L. Major advances in applied dairy cattle nutrition. Journal of Dairy Science, 2006, 89(4): 1311-1323 (doi: 10.3168/jds.S0022-0302(06)72199-3).
- Синещеков А.Д. Процессы питания и нервная регуляция их у сельскохозяйственных животных. Тезисы докладов VIII Всесоюзного съезда физиологов, биохимиков, фармакологов. М., 1955: 736.
- Батоев Ц.Ж. Динамика сокоотделения и выделение ферментов поджелудочной железы у птиц. Физиологический журнал СССР имени И.М. Сеченова, 1972, 58 (11): 1771-1773.
- Батоев Ц.Ж. Фотометрическое определение активности протеолитических ферментов поджелудочного сока по уменьшению концентрации казеина. Вопросы физиологии и патологии животных: Сборник трудов Бурятского государственного сельскохозяйственного института, 1971, 25: 22-26.
- Батоев Ц.Ж. Физиология пищеварения птиц. Улан-Удэ, 2001.
- Мажитова М.В. Спектрофотометрическое определение уровня метаболитов монооксида азота в плазме крови и ткани мозга белых крыс. Современные проблемы науки и образования, 2011, 3.
- Association of Official Agricultural Chemists. Official methods of analysis. 16th edition. Association of Official Agricultural Chemists, Washington DC, 1995.
- Hashemi S., Loh T., Foo H., Zulkifli I., Bejo M. Small intestine morphology, growth performance and nutrient digestibility of young broilers affected by different levels of dietary putrescine. Journal of Animal and Poultry Sciences, 2014, 3(3): 95-104.
- Humer E., Kröger I., Neubauer V., Reisinger N., Zebeli Q. Supplementation of a clay mineral-based product modulates plasma metabolomic profile and liver enzymes in cattle fed grain-rich diets. Animal, 2019, 13(6): 1214-1223 (doi: 10.1017/S1751731118002665).
- Hansen S.L., Ashwell M.S., Moeser A.J., Fry R.S., Knutson M.D., Spears J.W. High dietary iron reduces transporters involved in iron and manganese metabolism and increases intestinal permeability in calves. Journal of Dairy Science, 2010, 93(2): 656-65 (doi: 10.3168/jds.2009-2341).
- Jampilek J., Kos J., Kralova K. Potential of nanomaterial applications in dietary supplements and foods for special medical purposes. Nanomaterials, 2019, 9(2): 296 (doi: 10.3390/nano9020296).
- Lebedev S.V., Sheida E., Vertiprakhov V., Gavrish I., Kvan O., Gubaidullina I., Ryazanov V., Miroshnikov I. A study of the exocrinous function of the cattle pancreas after the introduction of feed with a various protein source in rations. Bioscience Research, 2019, 16(3): 2553-2562.
- Lebedev S.V., Gavrish I.A., Shejda E.V., Miroshnikov I.S., Ryazanov V.A., Gubajdullina I.Z., Makaeva A.M. Effect of various fats on digestibility of nutrients in diet of сalves. IOP Conf. Series: Earth and Environmental Science,2019, 341: 012066 (doi: 10.1088/1755-1315/341/1/012066).
- Khan Z., Al-Thabaiti S.A. Green synthesis of zero-valent Fe-nanoparticles: catalytic degradation of rhodamine B, interactions with bovine serum albumin and their enhanced antimicrobial activities. Journal of Photochemistry and Photobiology B: Biology, 2018, 180: 259-267 (doi: 10.1016/j.jphotobiol.2018.02.017).
- Вертипрахов В.Г., Грозина А.А., Фисинин В.И. Внешнесекреторная функция поджелудочной железы кур-несушек (GallusgallusL.) при добавлении в корм различных растительных масел. Сельскохозяйственная биология, 2020, 55(4): 726-737 (doi: 10.15389/agrobiology.2020.4.726rus).
- Lieu P.T., Heiskala M., Peterson P.A., Yang Y. The roles of iron in health and disease. Molecular Aspects of Medicine, 2001, 22(1-2): 1-87 (doi: 10.1016/s0098-2997(00)00006-6).
- Mody V.V., Siwale R., Singh A., Mody H.R. Introduction to metallic nanoparticles. J. Pharm. Bioallied Sci., 2010, 2(4): 282-289 (doi: 10.4103/0975-7406.72127).
- Naik P.K. Bypass fat in dairy ration-a review. Animal Nutrition and Feed Technology, 2013, 13: 147-163.
- Вертипрахов В.Г., Грозина А.А., Долгорукова А.М. Активность ферментов поджелудочной железы у цыплят-бройлеров на разных этапах пищеварения. Сельскохозяйственная биология, 2016, 4(51): 509-515 (doi: 10.15389/agrobiology.2016.4.509rus).
- Фисинин В.И., Егоров И.А., Вертипрахов В.Г., Грозина А.А., Ленкова Т.Н., Манукян В.А., Егорова Т.А. Активность пищеварительных ферментов в дуоденальном химусе и плазме крови у исходных линий гибридов мясных кур при использовании биологически активных добавок в рационе. Сельскохозяйственная биология, 2017, 6(52): 1226-1233 (doi: 10.15389/agrobiology.2017.6.1226rus).
- Palmquist D.L., Jenkins T.C. A 100-year review: fat feeding of dairy cows. Journal of Dairy Science,2017, 100(12): 10061-10077 (doi: 10.3168/jds.2017-12924).
- Al-Qushawi A., Rassouli A., Atyabi F., Peighambari S. M., Esfandyari-Manesh M., Shams G., Yazdani A. Preparation and characterization of three tilmicosin-loaded lipid nanoparticles: physicochemical properties and in-vitro antibacterial activities. Iran. J. Pharm. Res., 2016, 15(4): 663-676.
- Подоксенов Ю.К., Каменщиков Н.О., Мандель И.А. Применение оксида азота для защиты миокарда при ишемической болезни сердца. Анестезиология и реаниматология, 2019, 2: 34-47 (doi: 10.17116/anaesthesiology201902134).
- Rama Rao S.V., Prakash B., Raju M.V.L.N., Panda A.K., Kumari R.K., Pradeep Kumar Reddy E. Effect of supplementing organic forms of zinc, selenium and chromium on performance, anti-oxidant and immune responses in broiler chicken reared in tropical summer. Biological Trace Element Research, 2016, 172(2): 511-520 (doi: 10.1007/s12011-015-0587-x).
- Экзокринная недостаточность поджелудочной железы /А.А. Нижевич, О.А. Малиевский, А.Я. Валиулина, Л.В. Яковлева, Р.М. Файзуллина (сост.). Уфа, 2017.
- Hirota M, Ohmuraya M., Baba H. The role of trypsin, trypsin inhibitor, and trypsin receptor in the onset and aggravation of pancreatitis. J. Gastroenterol., 2006, 41(9): 832-836 (doi: 10.1007/s00535-006-1874-2)
- Snook J.T. Adaptive and nonadaptive changes in digestive enzyme capacity influencing digestive function. Feder. Proc., 1974, 33(1): 88-93.
- Farouk S.N., Muhammad A., Aminu M.A. Application of nanomaterials as antimicrobial agents: a review. Archives of Nanomedicine: Open Access Journal, 2018, 1(3): 59-64 (doi: 10.32474/anoaj.2018.01.000114).