doi: 10.15389/agrobiology.2022.2.207rus
УДК 636.4:619:578:616-097:57.083.3
Работа выполнена при финансовой поддержке РНФ (грант № 20-76-10030).
ИММУНОБИОЛОГИЧЕСКИЕ И МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ СВОЙСТВА НЕГЕМАДСОРБИРУЮЩИХ ШТАММОВ ВИРУСА АФРИКАНСКОЙ ЧУМЫ СВИНЕЙ (обзор)
С.А. КРУТЬКО, С.Г. НАМСРАЙН, А.Д. СЕРЕДА✉
Вирус африканской чумы свиней (АЧС, African swine fever virus, Asfivirus, Asfarviridae) представляет собой наиболее серьезную проблему для отрасли свиноводства во всем мире. В представленном обзоре проанализированы результаты исследований негемадсорбирующих штаммов вируса АЧС, рассмотрены особенности их иммунобиологических и молекулярно-генетических свойств, применение в фундаментальных и прикладных научных исследованиях. Согласно опубликованным данным, большинство выделенных в природе или полученных в лабораторных условиях негемадсорбирующих штаммов вируса АЧС слабовирулентны или авирулентны и обладают свойством формировать иммунную защиту от последующего заражения свиней гомологичными вирулентными гемадсорбирующими изолятами или штаммами (J.D. Vigário с соавт., 1970). На Африканском континенте авирулентные негемадсорбирующие штаммы вируса АЧС, как правило, выделяли от персистентно инфицированных бородавочников (Phacochoerus spp.), кустарниковых свиней (Potomochorusporcus) и мягких клещей Оrnithodoros moubata (A. Pini, 1976; G.R. Thomson с соавт., 1979), в Европе (Португалия, Испания, Латвия) и Азии (Китай) — от персистентно инфицированных домашних свиней (Sus scrofa domesticus), диких кабанов (Sus scrofa) и от клещей Оrnithodoros erraticus (marocanus) (F.S. Boinas с соавт., 2004; C. Gallardo с соавт., 2019; E. Sun с соавт., 2021). Негемадсорбирующие штаммы применяются в исследованиях механизмов иммунной защиты от АЧС. В экспериментах со штаммом OURT88/3 была установлена важная роль CD8+ Т-клеток в иммунной защите от АЧС. Индуцированная штаммом OURT88/3 перекрестная защита от заражения вирулентными изолятами неродственных генотипов коррелировала со способностью этих изолятов специфически стимулировать продукцию IFNγ лимфоцитами иммунизированных свиней (C.C. Abrams с соавт., 2013). В экспериментах с негемадсорбирующим штаммом NH/P68 было продемонстрировано, что высокое содержание специфических антител к вирусу АЧС характерно для хронической формы болезни, в то время как низкое количество антител отмечено у свиней с бессимптомной формой инфекции после интраназальной и внутримышечной иммунизации (A. Leitão с соавт., 2001; C. Gallardo с соавт., 2019). Низкую патогенность негемадсорбирующих изолятов связывают с потерей факторов вирулентности из-за больших делеций близко к левому концу генома или меньшими делециями и заменами в генах, кодирующих факторы вирулентности в других участках генома (F.S. Boinas с соавт., 2004). Утрату гемадсорбирующих свойств вируса АЧС связывают с делециями и/или сдвигом рамки считывания в гене EP402R (R.J. Rowlands с соавт., 2009; R. Portugal с соавт., 2015; K.A. Mima с соавт., 2015). Проведены работы по уменьшению побочных клинических реакций у свиней, инокулированных делеционными мутантами штаммов OURT88/3 и NH/P68 вируса АЧС (M.L. Nogal с соавт., 2001; C. Hurtado с соавт., 2004; A.G. Granja с соавт., 2009). Природно аттенуированные негемадсорбирующие штаммы вируса АЧС применяют в исследованиях по созданию кандидатных живых вакцин. С их помощью удалось достичь до 100 % защиты от гомологичных вирулентных изолятов и штаммов вируса АЧС у домашних свиней (K. King с соавт., 2011; P.J. Sánchez-Cordón с соавт., 2017; C. Gallardo с соавт., 2018; C. Gallardo с соавт., 2019; P.J. Sanchez-Cordon с соавт., 2020) и кабанов (J.A. Barasona с соавт., 2019).
Ключевые слова: африканская чума свиней, негемадсорбирующие изоляты, негемадсорбирующие штаммы, кандидатные вакцины.
Африканская чума свиней (АЧС) — одно из самых сложных инфекционных заболеваний свиней и диких кабанов. В настоящее время АЧС представляет собой наиболее серьезную проблему для отрасли свиноводства во всем мире. Из-за отсутствия коммерческих вакцин (1-3) единственным способом борьбы с АЧС остается тотальное уничтожение домашних свиней и диких кабанов в очагах инфекции (4, 5). В связи с серьезными экономическими последствиями для международной торговли свиньями и продуктами из свинины АЧС находится в списке болезней, подлежащих уведомлению.
Болезнь вызывает единственный представитель семейства Asfarviridae (Asfar, African swine fever virus) — крупный ДНК-содержащий вирус, поражающий домашних и диких свиней (Suidae) (6-8). В юго-восточных и южных регионах Африки АЧС поддерживается в сильватическом цикле передачи между бородавочниками (Phacochoerus spp.), кустарниковыми свиньями (Potomochorus porcus) и мягкими клещами (Оrnithodoros moubata) (9). Как правило, у природных хозяев, включая клещей, вирус АЧС вызывает субклиническую хроническую или инаппарантную формы инфекции (10, 11). Изоляты и штаммы вируса АЧС различаются по патогенным, антигенным, гемадсорбирующим, генетическим свойствам (12-14). В частности, на основе 3´-концевых последовательностей гена B646L, кодирующего основной капсидный белок p72, в Африке идентифицировано 24 генотипа вируса АЧС (15-17). Вирусом АЧС генотипа I были вызваны вспышки АЧС за пределами африканского континента с 1957 по 1991 годы в Португалии, Испании, Франции, Нидерландах, на о. Мадейра, в Италии, на Кубе, Мальте, о. Сардиния (Италия), в Бразилии, Доминиканской Республике, Гаити и СССР (18-20). Все они были ликвидированы (за исключением вспышки на о. Сардиния). В 2007 году из Африки в Грузию был занесен вирус АЧС генотипа II, получивший наименование штамм Georgia 2001/1, производные которого распространились в другие страны Кавказа и Европы (21-23). Общая картина развития заболеваемости свидетельствует, что АЧС получила эпизоотическое распространение с вовлечением популяций как домашних, так диких свиней (24-26). В 2018 году Georgia 2001/1-подобный вирус АЧС генотипа II был выделен в Китае и за три года распространился на 15 других азиатских стран (27-29). Однако, наряду с появлением низковирулентных изолятов генотипа II, в середине 2021 года в Китае от больных домашних свиней были выделены два штамма вируса АЧС генотипа I (30-31). Особое внимание специалистов вызвало то обстоятельство, что оба штамма были охарактеризованы как негемадсорбирующие (31).
В представленном обзоре проанализированы результаты исследований негемадсорбирующих штаммов вируса АЧС, рассмотрены особенности их иммунобиологических и молекулярно-генетических свойств, применение в фундаментальных и прикладных научных исследованиях. В ряде работ при описании исследуемых вирусов чаще используют термин «изолят». В контексте настоящей статьи мы в основном применяли термин «штамм», исходя из того, что штамм представляет собой идентифицированную по тестам современной классификации локальную популяцию вируса с оригинальными, стабильными свойствами (признаками), а изолят — вирус, выделенный из какого-либо конкретного источника.
Иммунобиологические свойства негемадсорбирующих штаммов вируса АЧС. Подавляющее большинство изолятов вируса АЧС охарактеризованы как вирулентные и гемадсорбирующие. В 1968 году L. Coggins сообщил о выделении негемадсорбирующих субпопуляций вируса АЧС (32). Дальнейшие исследования показали, что утрата свойства индуцировать при репродукции в культуре клеток гемадсорбцию для вируса АЧС явление распространенное (33-35). Ряд исследователей отмечали у выделенных в природе или полученных в лабораторных условиях негемадсорбирующих штаммов вируса АЧС низкую вирулентность и способность формировать иммунную защиту от последующего заражения свиней гомологичными вирулентными гемадсорбирующими изолятами (36, 37). На Африканском континенте низковирулентные или авирулентные негемадсорбирующие изоляты вируса АЧС, как правило, выделяли от персистентно инфицированных вирусом бородавочников, кустарниковых и домашних свиней, клещей рода Оrnithodoros (38-40). По данным комиссии ЕС, 1 % образцов, полученных с 1968 по 1976 годы от домашних свиней на Пиренейском полуострове, содержали негемадсорбирующие вирусы АЧС (41). Исследователи, проводившие в лабораторных условиях аттенуацию вируса АЧС с целью получения кандидатных живых вакцин, эмпирически пришли к способу селекции авирулентных штаммов по признаку снижения их способности индуцировать при репродукции in vitro гемадсорбцию (42). Следует отметить, что в природе иногда выделяли и вирулентные негемадсорбирующие изоляты. Например, из двух исследованных негемадсорбирующих изолятов один (Lillie-148) был вирулентным, второй (Zaire) вызывал болезнь и гибель только 33 % свиней (40).
Первые природные негемадсорбирующие изоляты вируса АЧС получили от свиней на юге Португалии, где в большей части стад свиньи были серопозитивные (43, 44). Этому предшествовали вспышки АЧС на Пиренейском полуострове в 1957 и 1960 годах, которые послужили началом эпизоотии, продолжавшейся до 1990-х годов и до сих пор сохраняющейся на о. Сардиния в Италии. Из выделенного во время второй вспышки вирулентного гемадсорбирующего штамма Lisbon 60 в результате 150 пассажей в первичной культуре клеток костного мозга свиней был получен гемадсорбирующий «вакцинный» штамм 1455. В процессе широкомасштабной вакцинации против АЧС в Португалии в 1960-х годах отмечали гибель 3-4 % привитых животных, у 7 % свиней развились неприемлемые поствакцинальные реакции, включающие пневмонию, двигательные нарушения, язвы в области кожи, аборты и гибель животных (45, 46). Позднее в Португалии от свиньи с хронической формой АЧС выделили непатогенный негемадсорбирующий штамм вируса АЧС NH/P68 (NHV, NHA2). Также и в Испании в период с 1965 по 1974 год получено 206 негемадсорбирующих изолятов (47). Отмечалось, что выделить негемадсорбирующие изоляты вируса АЧС было труднее, чем гемадсорбирующие, поскольку вызываемая ими виремия носила спорадический характер, а вирус в органах свиней накапливался в небольших количествах. Эксперименты показали, что непатогенные негемадсорбирующие изоляты передавались контактно менее эффективно (~40-50 %), чем вирулентные гемадсорбирующие изоляты (100 %). Заражение свиней непатогенными негемадсорбирующими изолятами могло быть причиной серопозитивности некоторых стад при отсутствии у свиней клинических симптомов. На основании того, что инокуляция штамма NH/P68 приводила к защите от последующего летального заражения вирулентным штаммом Lisbon 60, был сделан вывод об их антигенном родстве (48).
Двадцать лет спустя от клещей O. erraticus (marocanus), собранных на свиньях и в свинарниках на юге Португалии, были выделены два различающихся по патогенности типа изолятов вируса АЧС. Изоляты первого типа вызывали 100 % гибель свиней от острой формы АЧС, второго, в частности OURT88/3, — не вызывали клинических признаков заболевания и гибели животных, хотя антитела против вируса АЧС у всех инфицированных свиней были обнаружены (49). Предполагают, что прямым предком штамма OURT88/3 был штамм NH/P68, который также реплицируется в клещах и хорошо адаптирован к естественному циклу передачи (50). Касательно происхождения непатогенных негемадсорбирующих изолятов на Пиренейском полуострове выдвинуто два предположения: они получены либо из аттенуированного вакцинного штамма, либо от исходного вирулентного изолята, способного персистировать в цикле домашняя свинья—клещ—дикий кабан. Свиньи, инокулированные негемадсорбирующим штаммом OURT88/3, были защищены от гибели после заражения родственным патогенным гемадсорбирующим штаммом OURT88/1. Менее эффективная защита достигалась тогда, когда выздоровевшим свиньям вводили более отдаленные по родству изоляты или штаммы вируса АЧС. Отмечалось, что после заражения иммунных свиней штаммом Lisbon 57, выделенным во время первой вспышки АЧС в Португалии в 1957 году, и африканским изолятом Malawi LIL20/1 животные погибали, хотя начало клинического проявления болезни по времени было отложено (51-53).
Было проведено сравнение внутримышечного и интраназального путей иммунизации свиней различными дозами негемадсорбирующего штамма OURT88/3 вируса АЧС (54). При интраназальном введении сформировались две клинические группы: свиньи, у которых развивались периодические клинические проявления (дозы 103 и 104 TCID50, 100 % защиты от OURT88/1), и животные, у которых развилась хроническая форма АЧС (доза 105 TCID50, 66 % защиты). У свиней, иммунизированных внутримышечно низкими и средними дозами (103 и 104 TCID50), установлен более низкий процент защиты (50 и 66 %). В образцах крови на протяжении всего периода исследования обнаруживали низкое содержание генома вируса. Интересно, что внутримышечная иммунизация не приводила к появлению у защищенных свиней признаков хронической формы АЧС. Виремию не обнаруживали уже через 7 сут после инокуляции вируса. Эти результаты показали, что способ введения и доза вируса определяют результат иммунизации естественно ослабленным штаммом OURT88/3. В исследованиях с низковирулентным штаммом NH/P68 интраназальная иммунизация также индуцировала более высокую степень защиты, чем внутримышечная (53). Была установлена корреляция между поздней виремией после иммунизации NH/P68 (через 14 сут после инокуляции вируса) и появлением свиней с хронической формой АЧС. Такого соотношения не наблюдали у защищенных свиней, иммунизированных интраназально штаммом OURT88/3, где поздняя виремия была описана у животных без хронической формы АЧС (48, 55).
Установлено, что штамм OURT88/3 индуцирует высокую степень защиты от летального заражения родственными вирулентными изолятами вируса АЧС (49, 56, 57). Экспериментальная иммунизация свиней штаммом OURT88/3 с последующим заражением близкородственным вирулентным штаммом OURT88/1 индуцировала защитный иммунитет у европейских домашних свиней против заражения двумя вирулентными африканскими изолятами вируса АЧС: Benin 97/1 генотипа I из Западной Африки (85,7 %) и Uganda 1965 генотипа X из Восточной Африки (100 %). Более 78 % свиней, зараженных Benin 97/1, и 50 %, зараженных Uganda 1965, не проявляли признаков заболевания или развития виремии (57).
В Демократической Республике Конго в 1979 году от домашних свиней был выделен негемадсорбирующй штамм Mfuati-79 (иммунотип II, генотип I). Через 15 сут после внутримышечной иммунизации штаммом Mfuati-79 в дозах 103,0-104,0 ТЦД50 развивалась устойчивость свиней к внутримышечному заражению гомологичным вирулентным гемадсорбирующим штаммом Congo-49 (иммунотип II, генотип I) в дозах 105,5-107,5 ГАЕ50 (58).
В 2017 году в Латвии был выделен негемадсорбирующий изолят вируса АЧС — Lv/17/WB/Rie1 генотипа II (59). У свиней, инокулированных внутримышечно изолятом Lv/17/WB/Rie1, наблюдали бессимптомную форму инфекции, проявлялась периодическая и слабая виремия, в сыворотках крови отмечали высокое содержание вирусоспецифических антител. Кроме того, через 2 мес после первичного инфицирования изолятом Lv17/WB/Rie1 две свиньи, зараженные вирулентным гемадсорбирующим латвийским изолятом, выжили. Несмотря на то, что число животных было небольшим, эти результаты, по мнению авторов, открывают перспективу использования изолята Lv17/WB/Rie1 в качестве объекта для разработки живых аттенуированных вакцин, как это происходит со штаммами NH/P68 и OURT88/3. Важно, что исследование иллюстрирует естественную эволюцию вируса АЧС, включая появление со временем менее авирулентных негемадсорбирующих изолятов в отсутствие клещей рода Ornithodoros.
Впервые за 14 лет мировой панзоотии АЧС, вызванной штаммами, аналогичными Georgia 2007/1 генотипа II, за пределами Африки и о. Сардиния в провинциях Henan и Shandong КНР от домашних свиней выделили два штамма генотипа I (HeN/ZZ-P1/21 и SD/DY-I/21), которые были негемадсорбирующими и вызывали хроническую форму заболевания АЧС (31). Филогенетический анализ показал некоторые различия между штаммами HeN/ZZ-P1/21 и SD/DY-I/21 и их общность с португальскими штаммами NH/P68 и OURT88/3.
Уже много лет известно, что свиньи, выздоровевшие после заражения вирусом АЧС, могут быть защищены от болезни и/или гибели при последующем заражении родственными вирулентными изолятами вируса (60-62). Кроме того, свиньи, инокулированные естественно аттенуированными или селекционированными в лабораторных условиях в результате пассажей в культурах клеток вирусами АЧС, также могут быть защищены от заражения гомологичными вирулентными изолятами (49, 63). Негемадсорбирующие штаммы NH/P68 и OURT88/3 вируса АЧС эффективно используются при исследовании механизмов формирования иммунной защиты и для определения значения различных генов в патогенности вируса. В экспериментах со штаммом OURT88/3 была установлена важная роль CD8+ Т-клеток в иммунной защите от АЧС. Истощение моноклональными антителами этой субпопуляции клеток отменяло индуцированную штаммом OURT88/3 защиту от заражения вирулентным изолятом OURT88/1 (56). Возможная роль антител в защите от АЧС была показана в экспериментах по пассивному переносу антител от иммунизированных к интактным свиньям (64-66). Установлено, что нейтрализующие антитела недостаточно эффективны, но возможны другие, опосредованные антителами защитные функции, в частности антителозависимая цитотоксичность (64, 67).
В экспериментах со штаммом NH/P68 было продемонстрировано, что высокое содержание специфических антител к вирусу АЧС характерно для хронической формы болезни, в то время как низкое — отмечено у бессимптомных свиней после интраназальной и внутримышечной иммунизации (46, 57). Независимо от доз и способов иммунизации штаммом OURT88/3, высокое содержание антител против структурного белка р72 наблюдали у свиней как с признаками хронической формы АЧС, так и без них, и даже у 50 % иммунизированных свиней, которые не были защищены от заражения гомологичным вирулентным изолятом (68). Перекрестная защита, индуцированная штаммом OURT88/3 против заражения вирулентными изолятами вируса АЧС из неродственных генотипов, коррелировала со способностью этих изолятов специфически стимулировать продукцию IFNγ лимфоцитами иммунизированных свиней (57, 69).
Исследования по измерению активности NK-клеток у свиней, инокулированных штаммом NH/P68, позволили установить функциональную роль этого подмножества лимфоцитов в противовирусной защите. У животных, которые остались здоровыми после введения штамма NH/P68 и стали устойчивыми к заражению штаммом Lisbon 60, на 7-е сут после инокуляции было отмечено повышенное количество NK-клеток. У некоторых свиней высокая активность NK-клеток наблюдалась на протяжении всего эксперимента. В противоположность этому у животных, у которых после инокуляции NH/P68 развилась хроническая форма АЧС, активность NK-клеток была близкой или незначительно превышала показатели у контрольных животных. Вирулентные изоляты вируса АЧС подавляли активность NK-клеток у свиней (70). In vitro NK-активность мононуклеарных клеток свиней подавлялась как низко-, так и высоковирулентными изолятами вируса АЧС (71). Эти данные свидетельствуют в пользу представления о том, что иммунитет к вирусу АЧС зависит, по крайней мере частично, от клеточных механизмов, в частности от NK-клеток (48).
Структурная, функциональная, сравнительная геномика негемадсорбирующих штаммов вируса АЧС. В стратегии создания средств защиты от АЧС применение живой аттенуированной вакцины считают предпочтительным, поскольку она вызывает иммунные ответы против всех вирусных антигенов, с которыми обычно сталкивается хозяин во время инфекции (72, 73). С учетом этого в исследованиях преимущественно используют природно аттенуированные негемадсорбирующие штаммы OURT88/3 и NH/P68. В том числе изучают возможность уменьшить побочные клинические реакции у свиней, инокулированных делеционными мутантами штаммов OURT88/3 и NH/P68, при сохранении высокой защиты против гомологичных вирулентных изолятов вируса АЧС (74-76).
Известно, что делеция гена DP96R в ДНК вирулентного штамма E70 не влияла на характеристики роста вируса АЧС в культурах клеток макрофагов in vitro, но степень виремии у свиней, инокулированных делеционным мутантом вируса, были снижены в 100-1000 раз (77, 78). Подобныеделеции генов DP71L и DP96R из ДНК штамма OURT88/3 также не снижали репликацию делеционного мутанта OURT88/3ΔDP2 в первичных свиных макрофагах in vitro по сравнению с родительским штаммом OURT88/3. Однако две из шести свиней, инокулированных делеционным вирусом OURT88/3ΔDP2, не были защищены от последующего заражения вирулентным щтаммом OURT88/1, тогда как все шесть свиней, инокулированных родительским штаммом OURT88/3, оказались защищены (69). Удаление кодирующего ингибитор апоптоза гена A224L из генома штамма NH/P68 не повлияло на способность делеционного мутанта NH/P68DA224L защищать свиней от гомологичного вирулентного штамма Lisbon 60. Кандидатная рекомбинантная вакцина полностью защищала от гомологичного заражения Lisbon 60, но не от генотипа II Arm07 (за исключением одной свиньи, иммунизированной NH/P68DA224L). Напротив, родительский вирус NH/P68 полностью защищал животных после заражения Arm07. У свиней не только отсутствовали какие-либо заметные клинические признаки после заражения Arm07, но и не был обнаружен вирус в крови или тканях. Интересно, что от свиней, иммунизированных родительским негемадсорбирующим штаммом NH/P68, вирус эффективно передавался контрольным животным в течение 3-4 нед после первоначального заражения. Вместе с тем передачи вирулентного вируса Arm07 контрольным свиньям не наблюдали (72).
Очевиден вопрос: чем отличаются геномы вирулентных гемадсорбирующих и аттенуированных негемадсорбирующих штаммов вируса АЧС? Хорошо исследованы геномы двух негемадсобирующих штаммов вируса АЧС — NH/P68 и OURT88/3. У первого геном состоит из 172051 п.н., у второго — из 171719 п.н. В их геномах закодировано по 158 открытых рамок считывания (open reading frame, ORF), схожесть составляет 99,98 % (69, 79).
Большинство вариантов геномов вируса АЧС — результат наличия разного количества генов мультигенных семейств (multigene families, MGF) слева и справа вариабельных областей (left and right variable regions LVR и RVR). MGFs характерны для вируса АЧС и не имеют очевидной гомологии с другими известными генами. В зависимости от размера белков они разделены на 5 семейств: MGT-100,MGT-110,MGT-300,MGT-360 иMGF-505 (79, 80). Известно, что белки MGF играют важную роль на разных стадиях вирусной инфекции, модулируют транскрипцию и трансляцию в клетках хозяев. Например, было показано, что гены MGF-360 иMGF-505 важны для размножения штамма BA71V вируса АЧС в макрофагах (80, 81). Однако свойства большинства белков MGF до сих пор остаются не изученными.
Низкая патогенность негемадсорбирующих изолятов может быть связана с потерей факторов вирулентности из-за больших делеций близко к левому концу генома или меньшими делециями или заменами в генах, кодирующих факторы вирулентности в других участках генома (62). При сравнивании штаммов NH/P68 и OURT88/3 со штаммом Lisbon 60 основные различия обнаружены в левой части генома (рис.) (82). Расхождения отмечены также в центральной и в правой части генома. Основные различия установлены в генах белков MGF.
LVR геномов штаммов NH/P68 и OURT88/3 короче LVR генома штамма Lisbon 60 на 10 т.п.н. Примечательно, что кроме делеции присутствует вставка длиной 4458 п.н., которая расположена между MGF110-2Lи 110-13L. Вставка содержит гены белков MGF 110-4L, 110-5L, 110-9L, 100-1R, ORFs 285L и 86R. К сожалению, функции белков, кодируемых этими генами, неизвестны. Следующее различие — это отсутствие нуклеотидов в позициях 7244-8632 в геноме штамма NH/P68 в сравнении со штаммом Lisbon 60. Указанная делеция приводит к отсутствию генов MGF110-11L, 110-12Lи 110-13L. В результате еще одной делеции длиной 2173 п.н. штамм NH/P68 утратил ген MGF360-6L. Стоит отметить, что этот ген также отсутствует в непатогенном штамме BA71V. Третий участок, который отсутствует в геноме штамма NH/P68, — фрагмент 19809-29877 генома штамма Lisbon 60. Эта делеция приводит к потере или повреждению генов MGF 360-9L, 360-10L, 360-11L, 360-12L, 360-13L, 360-14L, 505-1R, 505-2R, 505-3R. Некоторые из них участвуют в репликации в макрофагах, в проявлении вирулентности, в инфицировании клещей и в иммунном ответе IFN типа I (82-84) (табл.). RVR области NH/P68 и Lisbon 60 также имеют различия. В основном это короткие вставки или делеции в генах, кодирующих белки MGF. Белок вирулентного штамма, кодируемый геном MGF 360-16R, на две аминокислоты короче своего гомолога негемадсорбирующего фенотипа. Белок MGF 505-11L у штаммов NH/P68 и OURT88/3 имеет вставку (ее длина — четыре аминокислоты). Также негемадсорбирующие штаммы имеют мутацию в гене MGF 100-2L, которая приводит к сдвигу рамки считывания и появлению стоп-кодона (85, 86).
Ответственным за феномен гемадсорбции при репродукции вируса АЧС является гликопротеин СD2v, который кодируется геном EP402R (87, 88). Этот гликопротеин гомологичен адгезивному рецептору Т-клеток мышей, человека, свиньи CD2. Удаление гена EP402R из генома вируса АЧС лишало его способности индуцировать адсорбцию свиных эритроцитов на поверхности инфицированных клеток, но не влияло на скорость его размножения in vitro. Экспрессия CD2v на поверхности вирусных частиц коррелирует с ассоциацией вирионов с эритроцитами в крови инфицированных свиней (88). Расчетная молекулярная масса полипептида гликопротеина СD2v около 45 кДа, зрелого гликопротеина с учетом углеводных цепей — 105-110 кДа (89, 90). По гидрофильному профилю это типичный трансмембранный гликопротеин, состоящий из четырех дифференцированных областей: гидрофобной лидирующей области на N-конце из 20 аминокислот, гидрофильной экстрацеллюлярной части из 183 аминокислот с 15-16 сайтами N-гликозилирования, трансмебранной области из 25 аминокислот и богатой пролином цитоплазматической С-терминальной части из 174 аминокислот. Сравнительная геномика показала, что открытая рамка считывания EP402R — одна из самых изменчивых в геноме вируса АЧС (91). Генотипирование по генетическому локусу, кодирующему CD2v, совпадает с группировкой штаммов вируса АЧС по сероиммуногруппам (92, 93). У штаммов вируса АЧС 26544/OG10, Benin 97/1 и BA71V полипептид, определяемый геном EP402R, состоит из 428 аминокислот, у штамма E75 — из 420, у негемадсорбирующих штаммов OURT/88 и NHV/P68 — из 330 (82).
Установлено, что взаимодействие между CD2v и его лигандом на эритроцитах стабилизируются посредством экспрессии вирусоспецифического лектиноподобного гликопротеина С-типа, кодируемого ORFEP153R, поскольку делеция гена EP153R приводила к снижению гемадсорбции вокруг зараженных вирусом АЧС клеток (85, 94).
Существует мнение, что гемадсорбция напрямую не связана с вирулентностью, поскольку известны негемадсорбирующие вирулентные изоляты (95, 96). Удаление генов EP402R или EP153R из генома изолята Malawi LIL20/1 не снижало его вирулентности для домашних свиней (85). Интересно, что гемадсорбирующий фенотип способствует усилению репликации вируса АЧС в клещах. Это наблюдали после восстановления гемадсорбирующей активности у штамма NHV/P68, при этом вирулентность для свиней не восстанавливалась (55). Оба гликопротеина выполняют и другие функции: CD2v обладает иммуносупрессивной активностью (28), С-лектиноподобный гликопротеин подавляет апоптоз и экспрессию на плазматической мембране антигена гистосовместимости SLA I (92).
Закономерен вопрос о молекулярных механизмах происхождения негемадсорбирующих штаммов вируса АЧС. У выделенных в природе штаммов, не способных к гемадсорбции, были обнаружены изменения в последовательности гена EP402R. Утрату гемадсорбирующих свойств у некоторых штаммов вируса АЧС связывают с делециями и/или сдвигами рамки считывания в гене EP402R (86, 97). Можно предположить, что, наряду с лишением способности индуцировать гемадсорбцию, должно утрачиваться свойство негемадсорбирующих изолятов формировать иммунотипоспецифическую защиту. Фактически это не так (98). Большинство негемадсорбирующих изолятов и лабораторных штаммов сохраняют способность индуцировать иммунотипоспецифическую защиту (99-101).
Таким образом, популяции вируса африканской чумы свиней (АЧС) и восприимчивых животных в Африке представляют собой коэволюционную биологическую систему. Изоляты вируса различаются по генетическим и иммунобиологическим характеристикам. На сегодня установлено 24 генотипа и 9 иммуносеротипов вируса АЧС, разнообразие изолятов по вирулентности, а также способности индуцировать гемадсорбцию. Негемадсорбирующие изоляты и штаммы представляют собой естественный элемент фенотипической гетерогенности вируса АЧС. Отсутствие гемадсорбирующих свойств объясняют делециями и/или сдвигами рамки считывания в гене EP402R, кодирующем оболочечный гликопротеин СD2v. Как правило, выделенные в природе изоляты или полученные в лабораторных условиях негемадсорбирующие штаммы вируса АЧС характеризуются низкой вирулентностью вплоть до отсутствия клинических симптомов после инокуляции свиньям, а также способностью формировать иммунную защиту от последующего заражения гомологичными вирулентными гемадсорбирующими изолятами. Поэтому негемадсорбирующие природные, лабораторно селекционированные и рекомбинаниные штаммы вируса АЧС используют как для получения фундаментальных знаний о механизмах формирования защитного иммунитета, так и для разработки перспективных живых вакцин против АЧС.
Семейства MGF, присутствующие в LVR и RVR геномов вирулентного гемадсорбирующего штамма вируса африканской чумы свиней Lisbon 60 и авирулентного негемадсорбирующего штамма NH/P68 (NHV) (86) |
|||
Регион генома |
Семейство MGF |
Штаммы |
|
Lisbon 60 |
NH/P68 |
||
LVR (левый) |
MGF 100 |
- |
1R |
MGF 110 |
1L, 13L+2L, 11L, 12L, 13L, 14L |
1L, 2L, 4L, 5L, 9L, 13L§, 14L |
|
MGF 300 |
1L, 2R, 4L |
1L, 2R, 4L |
|
MGF 360 |
1L, 2L, 3L, 4L, 6L, 8L, 9L, 10L, 11L, 12L, 13L, 14L |
1L, 2L, 3L, 4L, 8L, 9L‡ |
|
MGF 505 |
1R, 2R, 3R, 4R, 5R, 7R, 8R, 9R, 10R |
3R‡, 4R, 5R, 7R, 8R, 9R, 10R |
|
RVR (правый) |
MGF 100 |
2L |
2L‡ |
MGF 360 |
16R, 18R |
16R, 17R‡, 18R |
|
MGF 505 |
11L |
11L |
|
Примечание. Прочерк означает отсутствие членов семейства MGF указанного типа. ‡ — усеченная версия гена, § — отличается от того же гена в штамме OURT88/3. |
|||
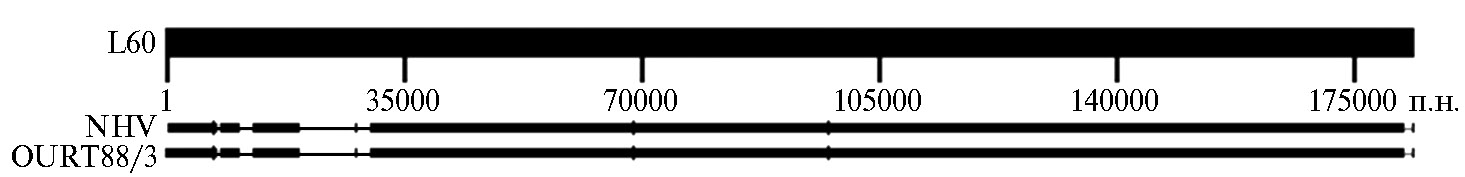
Сравнение генома штамма вируса африканской чумы свиней Lisbon 60 (L60) с геномами штаммов NH/P68 (NHV) и OURT88/3. Широкая черная линия — участки гомологии геномов NHV и OURT88/3 с геномом Lisbon 60. Тонкая черная линия — делеции в геномах NHV и OURT88/3, вертикальные черные линии — вставки, которые присутствуют у NHV и OURT88/3.
ЛИТЕРАТУРА
- Arias M., De la Torre A., Dixon L., Gallardo C., Jori F., Laddomada A., Martins C., Parkhouse R.M., Revilla Y., Rodriguez F., Sanchez-Vizcaino J.M. Approaches and perspectives for development of African swine fever virus vaccines. Vaccines, 2017, 5(4): 35 (doi: 10.3390/vaccines5040035).
- Forman A.J., Wardley R.C., Wilkinson P.J. The immunological response of pigs and guinea pigs to antigens of African swine fever virus. Archives of Virology, 1982, 74(2-3): 91-100 (doi: 10.1007/BF01314703).
- Stone S.S., Hess W.R. Antibody response to inactivated preparations of African swine fever virus in pigs. American Journal of Veterinary Research, 1967, 28(123): 475-481.
- Rock D.L. Challenges for African swine fever vaccine development—«... perhaps the end of the beginning». Veterinary Microbiology, 2017, 206: 52-58 (doi: 10.1016/j.vetmic.2016.10.003).
- Blome S., Gabriel C., Beer M. Modern adjuvants do not enhance the efficacy of an inactivated African swine fever virus vaccine preparation. Vaccine, 2014, 32(31): 3879-3882 (doi: 10.1016/j.vaccine.2014.05.051).
- OIE (World Organization for Animal Health). Animal diseases. Режим доступа: https://www.oie.int/en/what-we-do/animal-health-and-welfare/animal-diseases/. Без даты.
- Alonso C., Borca M., Dixon L., Revilla Y., Rodriguez F., Escribano J.M. Ictv report consortium. ICTV virus taxonomy profile: Asfarviridae. Journal of General Virology, 2018, 99(5): 613-614 (doi: 10.1099/jgv.0.001049).
- Galindo I., Alonso C. African swine fever virus: a review. Viruses, 2017, 9(5): 103 (doi: 10.3390/v9050103).
- Sánchez-Vizcaíno J.M., Mur L., Martínez-López B. African swine fever: an epidemiological update. Transboundary and Emerging Disease, 2012, 59(S1): 27-35 (doi: 10.1111/j.1865-1682.2011.01293.x).
- Global African Swine Fever Research Alliance (GARA). African Swine Fever. Gap Analysis Report. 2018. Режим доступа: https://go.usa.gov/xPfWr. Без даты.
- Sereda A.D., Balyshev V.M., Kazakova A.S., Imatdinov A.R., Kolbasov D.V. Protective properties of attenuated strains of African swine fever virus belonging to seroimmunotypes I-VIII. Pathogens, 2020, 9(4): 274 (doi: 10.3390/pathogens9040274).
- Malogolovkin A., Kolbasov D. Genetic and antigenic diversity of African swine fever virus. Virus Research, 2019, 271: 197673 (doi: 10.1016/j.virusres.2019.197673).
- Sereda A.D., Imatdinov A.R., Makarov V.V. The haemadsorbation at African swine fever (review). Sel’skokhozyaistvennaya Biologiya [Agricultural Biology], 2016, 51(6): 763-774 (doi: 10.15389/agrobiology.2016.6.763eng).
- Quembo C.J., Jori F., Vosloo W., Heath L. Genetic characterization of African swine fever virus isolates from soft ticks at the wildlife/domestic interface in Mozambique and identification of a novel genotype. Transboundary and Emerging Disease, 2018, 65(2): 420-431 (doi: 10.1111/tbed.12700).
- Achenbach J.E., Gallardo C., Nieto-Pelegrín E., Rivera-Arroyo B., Degefa-Negi T., Arias M., Jenberie S., Mulisa D.D., Gizaw D., Gelaye E., Chibssa T.R., Belaye A., Loitsch A., Forsa M., Yami M., Diallo A., Soler A., Lamien C.E., Sánchez-Vizcaíno J.M. Identification of a new genotype of African swine fever virus in domestic pigs from Ethiopia. Transboundary and Emerging Disease, 2017, 64(5): 1393-1404 (doi: 10.1111/tbed.12511).
- Muangkram Y., Sukmak M., Wajjwalku W. Phylogeographic analysis of African swine fever virus based on the p72 gene sequence. Genetics and Molecular Research, 2015, 14(2): 4566-4574 (doi: 10.4238/2015.May.4.15).
- Gao L., Sun X., Yang H., Xu Q., Li J., Kang J., Liu P., Zhang Y., Wang Y., Huang B. Epidemic situation and control measures of African Swine Fever Outbreaks in China 2018-2020. Transboundary and Emerging Disease, 2021, 68(5): 2676-2686 (doi: 10.1111/tbed.13968).
- Revilla Y., Pérez-Núñez D., Richt J.A. Chapter Three - African swine fever virus biology and vaccine approaches. Advances in Virus Research, 2018, 100: 41-74 (doi: 10.1016/bs.aivir.2017.10.002).
- Cisek AA., Dąbrowska I., Gregorczyk K.P., Wyżewski Z. African swine fever virus: a new old enemy of Europe. Annals of Parasitology, 2016, 62(3): 161-167 (doi: 10.17420/ap6203.49).
- Sánchez-Vizcaíno J.M., Mur L., Gomez-Villamandos J.C., Carrasco L. An update on the epidemiology and pathology of African swine fever. Journal of Comparative Pathology, 2015, 152(1): 9-21 (doi: 10.1016/j.jcpa.2014.09.003).
- Rowlands R.J., Michaud V., Heath L., Hutchings G., Oura C., Vosloo W., Dwarka R., Onashvili T., Albina E., Dixon L.K. African swine fever virus isolate, Georgia, 2007. Emerging Infectious Diseases, 2008, 14(12): 1870-1874 (doi: 10.3201/eid1412.080591).
- Sánchez-Vizcaíno J.M., Mur L., Martínez-Lópeza B. African swine fever (ASF): five years around Europe. Veterinary Microbiology, 2013, 165(1-2): 45-50 (doi: 10.1016/J.VETMIC.2012.11.030).
- Beltran-Alcrudo D., Lubroth J., Depner K., Rocque, La S.D., Beltran-Alcrudo D., Lubroth J., De La Rocque S. African swine fever in the Caucasus. FAO, EmpresWatch, 2008.
- Gogin A., Gerasimov V., Malogolovkin A., Kolbasov D. African swine fever in the North Caucasus region and the Russian Federation in years 2007-2012. Virus Research, 2013, 173(1): 198-203 (doi: 10.1016/j.virusres.2012.12.007).
- Korennoy F.I., Gulenkin V.M., Gogin A.E., Vergne T., Karaulov A.K. Estimating the basic reproductive number for African swine fever using the Ukrainian historical epidemic of 1977. Transboundary and Emerging Disease, 2017, 64(6): 1858-1866 (doi: 10.1111/tbed.12583).
- Malogolovkin A., Yelsukova A., Gallardo C., Tsybanov S., Kolbasov D. Molecular characterization of African swine fever virus isolates originating from outbreaks in the Russian Federation between 2007 and 2011. Veterinary Microbiology, 2012, 58(3-4): 415-9 (doi: 10.1016/j.vetmic.2012.03.002).
- Zhao D., Liu R., Zhang X., Li F., Wang J., Zhang J., Liu X., Wang L., Zhang J., Wu X., Guan Y., Chen W., Wang X., He X., Bu Z. Replication and virulence in pigs of the first African swine fever virus isolated in China. Emerging Microbes and Infections, 2019, 8(1): 438-447 (doi: 10.1080/22221751.2019.1590128).
- Tran H.T.T., Truong A.D., Dang A.K., Ly D.V., Nguyen C.T., Chu N.T., Hoang T.V., Nguyen H.T., Dang H.V. Circulation of two different variants of intergenic region (IGR) located between the I73R and I329L genes of African swine fever virus strains in Vietnam. Transboundary and Emerging Disease, 2021, 68(5): 2693-2695 (doi: 10.1111/tbed.13996).
- Wen X., He X., Zhang X., Zhang X., Liu L., Guan Y., Zhang Y., Bu Z. Genome sequences derived from pig and dried blood pig feed samples provide important insights into the transmission of African swine fever virus in China in 2018. Emerging Microbes and Infections, 2019, 8(1): 303-306 (doi: 10.1080/22221751.2019.1565915).
- Sun E., Zhang Z., Wang Z., He X., Zhang X., Wang L., Wang W., Huang L., Xi F., Huangfu H., Tsegay G., Huo H., Sun J., Tian Z., Xia W., Yu X., Li F., Liu R., Guan Y., Zhao D., Bu Z. Emergence and prevalence of naturally occurring lower virulent African swine fever viruses in domestic pigs in China in 2020. Sci. China Life Sci., 2021, 64(5): 752-765 (doi: 10.1007/s11427-021-1904-4).
- Sun E., Huang L., Zhang X., Zhang J., Shen D., Zhang Z., Wang Z., Huo H., Wang W., Huangfu H., Wang W., Li F., Liu R., Sun J., Tian Z., Xia W., Guan Y., He X., Zhu Y., Zhao D., Bu Z. Genotype I African swine fever viruses emerged in domestic pigs in China and caused chronic infection. Emerging Microbes and Infections, 2021, 10(1): 1-30 (doi: 10.1080/22221751.2021.1999779).
- Coggins L. Segregation of a nonhaemadsorbing African swine fever virus in tissue culture. Cornell Veterinarian, 1968, 58: 12-20.
- Jori F., Bastos A.D. Role of wild suids in the epidemiology of African swine fever. EcoHealth, 2009, 6(2): 296-310 (doi: 10.1007/s10393-009-0248-7).
- Ravaomanana J., Michaud V., Jori F., Andriatsimahavandy A., Roger F., Albina E., Vial L. First detection of African Swine Fever Virus in Ornithodoros porcinus in Madagascar and new insights into tick distribution and taxonomy. Parasites Vectors,2010, 3: 115 (doi: 10.1186/1756-3305-3-115).
- Pan I.C., Hess W.R. Diversity of African swine fever virus. American Journal of Veterinary Research, 1985, 46(2): 314-320.
- Pan I.C. African swine fever virus: generation of subpopulations with altered immunogenicity and virulence following passage in cell cultures. The Journal of Veterinary Medical Science, 1992, 54(1): 43-52 (doi: 10.1292/jvms.54.43).
- Vigário J.D., Terrinha A.M., Bastos A.L., Moura-Nunes J.F., Marques D., Silva J.F. Serological behaviour of isolated African swine fever virus. Brief report. Archiv für die Gesamte Virusforschung, 1970, 31(3): 387-389 (doi: 10.1007/BF01253773).
- Pini A. Isolation and segregation of non-haemadsorbing strains of African swine fever virus. Veterinary Record, 1976, 99(24): 479-480 (doi: 10.1136/vr.99.24.479).
- Pini A., Wagenaar G. Isolation of a non-haemadsorbing strain of African swine fever (ASF) virus from a natural outbreak of the disease. Veterinary Record, 1974, 94(1): 2 (doi: 10.1136/vr.94.1.2).
- Thomson G.R., Gainaru M.D., van Dellen A. F. African swine fever: pathogenicity and immunogenicity of two non-haemadsorbing viruses. The Onderstepoort Journal of Veterinary Research, 1979, 46(3): 149-154.
- Commission of the European Communities. Laboratory manual for research on classical and African swine fever. The Commission, Luxembourg, 1976: 111-112.
- Makarov V., Nedosekov V., Sereda A., Matvienko N. Immunological conception of African swine fever. Zoology and Ecology, 2016, 26(3): 236-243 (doi: 10.1080/21658005.2016.1182822).
- Louzã A.C., Boinas F.S., Caiado J.M., Vigario J.D., Hess W.R. Role des vecteurs et des reservoirs animaux dans la persistence de la peste porcine africaine, au Portugal. Epidemiologie et Sante Animale, 1989, 15: 89-102 (in French).
- Vigário J.D., Terrinha A.M., Moura Nunes J.F. Antigenic relationships among strains of African swine fever virus. Archiv für die Gesamte Virusforschung, 1974, 45(3): 272-277 (doi: 10.1007/BF01249690).
- Manso-Ribeiro J., Nunes-Petisca J.L., Lopez-Frazao F., Sobral M. Vaccination against ASF. Bull. Off. Int. Epiz., 1963, 60: 921-937.
- Petuska N. Quelques aspects morphogenesis des suites de la vaccination contre la PPA (virose L) an Portugal. Bull. Off. Int. Epiz., 1965, 63: 199-237.
- Sanchez Botija C., Ordaz A., Solana A., Gonzalvo F., Olias J. Carnero M.E. Peste Porcina Africana: observaciones sobre modificacion espontanea del virus de campo. An. Inst. Invest. Vet., 1977, 24: 7-17 (in Spanish).
- Leitão A., Cartaxeiro C., Coelho R., Cruz B., Parkhouse R., Portugal F.C., Vigário J.D., Martins C. The non-haemadsorbing African swine fever virus isolate ASFV/NH/P68 provides a model for defining the protective anti-virus immune response. The Journal of General Virology, 2001, 82(3): 513-523 (doi: 10.1099/0022-1317-82-3-513).
- Boinas F.S., Hutchings G.H., Dixon L.K., Wilkinson P.J. Characterization of pathogenic and non-pathogenic African swine fever virus isolates from Ornithodoros erraticus inhabiting pig premises in Portugal. Journal of General Virology, 2004, 85(8): 2177-2187 (doi: 10.1099/vir.0.80058-0).
- Rowlands R.J., Duarte M.M., Boinas F., Hutchings G., Dixon L.K. The CD2v protein enhances African swine fever virus replication in the tick vector, Ornithodoros erraticus. Virology, 2009, 393(2): 319-328 (doi: 10.1016/j.virol.2009.07.040).
- Dixon L.K., Twigg S.R., Baylis S.A., Vydelingum S., Bristow C., Hammond J.M., Smith G.L. Nucleotide sequence of a 55 kbp region from the right end of the genome of a pathogenic African swine fever virus isolate (Malawi LIL20/1). The Journal of General Virology, 1994, 75(7): 1655-1684 (doi: 10.1099/0022-1317-75-7-1655).
- Sumption K.J., Hutchings G.H., Wilkinson P.J., Dixon L.K. Variable regions on the genome of Malawi isolates of African swine fever virus. The Journal of General Virology, 1990, 71(10): 2331-2340 (doi: 10.1099/0022-1317-71-10-2331).
- Yáñez R.J., Rodríguez J.M., Nogal M.L., Yuste L., Enríquez C., Rodriguez J.F., Viñuela E. Analysis of the complete nucleotide sequence of African swine fever virus. Virology, 1995, 208(1): 249-278 (doi: 10.1006/viro.1995.1149).
- Sánchez-Cordón P.J., Chapman D., Jabbar T., Reis A.L., Goatley L., Netherton C.L., Taylor G., Montoya M., Dixon L. Different routes and doses influence protection in pigs immunised with the naturally attenuated African swine fever virus isolate OURT88/3. Antiviral Reserch, 2017, 138: 1-8 (doi: 10.1016/j.antiviral.2016.11.021).
- Gallardo C., Nieto R., Soler A., Pelayo V., Fernández-Pinero J., Markowska-Daniel I., Pridotkas G., Nurmoja I., Granta R., Simón A., Pérez C., Martín E., Fernández-Pacheco P., Arias M. Assessment of African swine fever diagnostic techniques as a response to the epidemic outbreaks in eastern european union countries: How To Improve surveillance and control programs. Journal of Clinical Microbiology, 2015, 53(8): 2555-2565 (doi: 10.1128/JCM.00857-15).
- Oura C., Denyer M.S., Takamatsu H., Parkhouse R. In vivo depletion of CD8+ T lymphocytes abrogates protective immunity to African swine fever virus. The Journal of General Virology, 2005, 86(9): 2445-2450 (doi: 10.1099/vir.0.81038-0).
- King K., Chapman D., Argilaguet J.M., Fishbourne E., Hutet E., Cariolet R., Hutchings G., Oura C.A., Netherton C.L., Moffat K., Taylor G., Le Potier M.F., Dixon L.K., Takamatsu H.H. Protection of European domestic pigs from virulent African isolates of African swine fever virus by experimental immunisation. Vaccine, 2011, 29(28): 4593-4600 (doi: 10.1016/j.vaccine.2011.04.052).
- Рудобельский Э.В. Исследование иммунобиологических свойств культурального варианта цитолитического штамма МФ-79 вируса АЧС. Тез. докл. научной конференции ВНИИВВиМ, посвященной 70-летию Великого Октября. Покров, 1988: 80-81.
- Gallardo C., Soler A., Rodze I., Nieto R., Cano-Gómez C., Fernandez-Pinero J., Arias M. Attenuated and non-haemadsorbing (non-HAD) genotype II African swine fever virus (ASFV) isolated in Europe, Latvia 2017. Transboundary and Emerging Diseases, 2019, 66(3): 1399-1404 (doi: 10.1111/tbed.13132).
- Detray D.E. Persistence of viremia and immunity in African swine fever. American Journal of Veterinary Research, 1957, 18(69): 811-816.
- Malmquist W.A. Serologic and immunologic studies with African swine fever virus. American Journal of Veterinary Research, 1963, 24: 450-459.
- Mebus C.A., Dardiri A.H. Western hemisphere isolates of African swine fever virus: asymptomatic carriers and resistance to challenge inoculation. American Journal of Veterinary Research, 1980, 41(11): 1867-1869.
- Sereda A.D., Kazakova A.S., Imatdinov A.R., Kolbasov D.V. Humoral and cell immune mechanisms under African swine fever. Agricultural Biology, 2015, 50(6): 709-718 (doi: 10.15389/agrobiology.2015.6.709eng).
- Wardley R.C., Norley S.G., Wilkinson P.J., Williams S. The role of antibody in protection against African swine fever virus. Veterinary Immunology and Immunopathology, 1985, 9(3): 201-212 (doi: 10.1016/0165-2427(85)90071-6).
- Onisk D.V., Borca M.V., Kutish G., Kramer E., Irusta P., Rock D.L. Passively transferred African swine fever virus antibodies protect swine against lethal infection. Virology, 1994, 198(1): 350-354 (doi: 10.1006/viro.1994.1040).
- Ruiz-Gonzalvo F., Carnero M.E., Bruyel V. Immunological responses of pigs to partially attenuated African swine fever virus and their resistance to virulent homologous and heterologous viruses. Proc. EUR 8466 EN CEC/FAO Research Seminar «African Swine Fever», Sardinia, Italy /P.J. Wilkinson (ed.). Luxemburg, Belgium, Commission of the European Communities, 1981: 206-216.
- Середа А.Д., Соловкин С.Л., Фугина Л.Г., Макаров В.В. Иммунные реакции на вирус африканской чумы свиней. Вопросы вирусологии, 1992, 37(3): 168-170.
- Sánchez-Cordón P.J., Jabbar T., Chapman D., Dixon L.K., Montoya M. Absence of long-term protection in domestic pigs immunized with attenuated African swine fever virus isolate OURT88/3 or BeninΔMFG correlates with increased levels of regulatory T cells and IL-10. Journal of Virology, 2020, 94(14), e00350-20 (doi: 10.1128/jvi.00350-20).
- Abrams C.C., Goatley L., Fishbourne E., Chapman D., Cooke L., Oura C.A., Netherton C.L., Takamatsu H.H., Dixon L.K. Deletion of virulence associated genes from attenuated African swine fever virus isolate OUR T88/3 decreases its ability to protect against challenge with virulent virus. Virology, 2013, 443(1): 99-105 (doi: 10.1016/j.virol.2013.04.028).
- Norley S.G., Wardley R.C. Investigation of porcine natural-killer cell activity with reference to African swine-fever virus infection. Immunology, 1983, 49(4): 593-597.
- Mendoza C., Videgain S.P., Alonso F. Inhibition of natural killer activity in porcine mononuclear cells by African swine fever virus. Research in Veterinary Science, 1991, 51(3): 317-321 (doi: 10.1016/0034-5288(91)90084-2).
- Gallardo C., Sánchez E.G., Pérez-Núñez D., Nogal M., de León P., Carrascosa Á.L., Nieto R., Soler A., Arias M.L., Revilla Y. African swine fever virus (ASFV) protection mediated by NH/P68 and NH/P68 recombinant live-attenuated viruses. Vaccine, 2018, 36(19): 2694-2704 (doi: 10.1016/j.vaccine.2018.03.040).
- Granja A.G., Sánchez E.G., Sabina P., Fresno M., Revilla Y. African swine fever virus blocks the host cell antiviral inflammatory response through a direct inhibition of PKC-theta-mediated p300 transactivation. Journal of Virology, 2009, 83(2): 969-980 (doi: 10.1128/JVI.01663-08).
- Nogal M.L., González de Buitrago G., Rodríguez C., Cubelos B., Carrascosa A.L., Salas M.L., Revilla Y. African swine fever virus IAP homologue inhibits caspase activation and promotes cell survival in mammalian cells. Journal of Virology, 2001, 75(6): 2535-2543 (doi: 10.1128/JVI.75.6.2535-2543.2001).
- Hurtado C., Granja A.G., Bustos M.J., Nogal M.L., González de Buitrago G., de Yébenes V.G., Salas M.L., Revilla Y., Carrascosa A.L. The C-type lectin homologue gene (EP153R) of African swine fever virus inhibits apoptosis both in virus infection and in heterologous expression. Virology, 2004, 326(1): 160-170 (doi: 10.1016/j.virol.2004.05.019).
- Revilla Y., Callejo M., Rodríguez J.M., Culebras E., Nogal M.L., Salas M.L., Viñuela E., Fresno M. Inhibition of nuclear factor kappaB activation by a virus-encoded IkappaB-like protein. The Journal of Biological Chemistry, 1998, 273(9): 5405-5411 (doi: 10.1074/jbc.273.9.5405).
- Zsak L., Caler E., Lu Z., Kutish G.F., Neilan J.G., Rock D L. A nonessential African swine fever virus gene UK is a significant virulence determinant in domestic swine. Journal of Virology, 1998, 72(2): 1028-1035 (doi: 10.1128/JVI.72.2.1028-1035.1998).
- Afonso C.L., Zsak L., Carrillo C., Borca M.V., Rock D.L. African swine fever virus NL gene is not required for virus virulence. The Journal of General Virology, 1998, 79(10): 2543-2547 (doi: 10.1099/0022-1317-79-10-2543).
- Keßler C., Forth J.H., Keil G.M., Mettenleiter T.C., Blome S., Karger A. The intracellular proteome of African swine fever virus. Scientific Reports, 2018, 8(1): 14714 (doi: 10.1038/s41598-018-32985-z).
- Zsak L., Lu Z., Burrage T.G., Neilan J.G., Kutish G.F., Moore D.M., Rock D.L. African swine fever virus multigene family 360 and 530 genes are novel macrophage host range determinants. Journal of Virology, 2001, 75(7): 3066-3076 (doi: 10.1128/JVI.75.7.3066-3076.2001).
- Tulman E.R., Rock D.L. Novel virulence and host range genes of African swine fever virus. Current Opinion in Microbiology, 2001, 4(4): 456-461 (doi: 10.1016/s1369-5274(00)00235-6).
- Bacciu D., Deligios M., Sanna G., Madrau M.P., Sanna M.L., Dei Giudici S., Oggiano A. Genomic analysis of Sardinian 26544/OG10 isolate of African swine fever virus. Virology Report, 2016, 6: 81-89 (doi: 10.1016/j.virep.2016.09.001).
- Afonso C.L., Piccone M.E., Zaffuto K.M., Neilan J., Kutish G.F., Lu Z., Balinsky C.A., Gibb T.R., Bean T.J., Zsak L., Rock D.L. African swine fever virus multigene family 360 and 530 genes affect host interferon response. Journal of Virology, 2004, 78(4): 1858-1864 (doi: 10.1128/jvi.78.4.1858-1864.2004).
- Burrage T.G., Lu Z., Neilan J.G., Rock D.L., Zsak L. African swine fever virus multigene family 360 genes affect virus replication and generalization of infection in Ornithodoros porcinus ticks. Journal of Virology, 2004, 78(5): 2445-2453 (doi: 10.1128/jvi.78.5.2445-2453.2004).
- Neilan J.G., Borca M.V., Lu Z., Kutish G.F., Kleiboeker S.B., Carrillo C., Zsak L., Rock D. L. An African swine fever virus ORF with similarity to C-type lectins is non-essential for growth in swine macrophages in vitro and for virus virulence in domestic swine. The Journal of General Virology, 1999, 80(10): 2693-2697 (doi: 10.1099/0022-1317-80-10-2693).
- Portugal R., Coelho J., Höper D., Little N.S., Smithson C., Upton C., Martins C., Leitão A., Keil G.M. Related strains of African swine fever virus with different virulence: genome comparison and analysis. The Journal of General Virology, 2015, 96(2): 408-419 (doi: 10.1099/vir.0.070508-0).
- Rodríguez J.M., Yáñez R.J., Almazán F., Viñuela E., Rodriguez J.F. African swine fever virus encodes a CD2 homolog responsible for the adhesion of erythrocytes to infected cells. Journal of Virology, 1993, 67(9): 5312-5320 (doi: 10.1128/JVI.67.9.5312-5320.1993).
- Borca M.V., Kutish G.F., Afonso C.L., Irusta P., Carrillo C., Brun A., Sussman M., Rock D. L. An African swine fever virus gene with similarity to the T-lymphocyte surface antigen CD2 mediates hemadsorption. Virology, 1994, 199(2): 463-468 (doi: 10.1006/viro.1994.1146).
- Kay-Jackson P.C., Goatley L.C., Cox L., Miskin J.E., Parkhouse R., Wienands J., Dixon L.K. The CD2v protein of African swine fever virus interacts with the actin-binding adaptor protein SH3P7. The Journal of General Virology, 2004, 85(1): 119-130 (doi: 10.1099/vir.0.19435-0).
- Goatley L.C., Dixon L.K. Processing and localization of the African swine fever virus CD2v transmembrane protein. Journal of Virology, 2011, 85(7): 3294-3305 (doi: 10.1128/JVI.01994-10).
- Chapman D., Tcherepanov V., Upton C., Dixon L.K. Comparison of the genome sequences of non-pathogenic and pathogenic African swine fever virus isolates. The Journal of General Virology, 2008, 89(2): 397-408 (doi: 10.1099/vir.0.83343-0).
- Tulman E.R., Delhon G.A., Ku B.K., Rock D.L. African swine fever virus. In: Lesser known large dsDNA viruses. Current topics in microbiology and immunology, vol. 328/J.L. Van Etten (ed.). Springer, Berlin, Heidelberg, 2009: 43-87 (doi: 10.1007/978-3-540-68618-7_2).
- Malogolovkin A., Burmakina G., Tulman E.R., Delhon G., Diel D.G., Salnikov N., Kutish G.F., Kolbasov D., Rock D.L. African swine fever virus CD2v and C-type lectin gene loci mediate serological specificity. The Journal of General Virology, 2015, 96(4), 866-873 (doi: 10.1099/jgv.0.000024).
- Galindo I., Almazán F., Bustos M.J., Viñuela E., Carrascosa A.L. African swine fever virus EP153R open reading frame encodes a glycoprotein involved in the hemadsorption of infected cells. Virology, 2000, 266(2): 340-351 (doi: 10.1006/viro.1999.0080).
- Gonzague M., Roger F., Bastos A., Burger C., Randriamparany T., Smondack S., Cruciere C. Isolation of a non-haemadsorbing, non-cytopathic strain of African swine fever virus in Madagascar. Epidemiology and Infection, 2001, 126(3): 453-459 (doi: 10.1017/s0950268801005465).
- Pan I.C., Hess W.R. Virulence in African swine fever: its measurement and implications. American Journal of Veterinary Research, 1984, 45(2): 361-366.
- Mima K.A., Burmakina G.S., Titov I.A., Malogolovkin A.S. African swine fever virus glycoproteins p54 and CD2v in the context of immune response modulation: bioinformatic analysis of genetic variability and heterogeneity. Agricultural Вiology, 2015, 50(6): 785-793 (doi: 10.15389/agrobiology.2015.6.785eng).
- Barasona J.A., Gallardo C., Cadenas-Fernández E., Jurado C., Rivera B., Rodríguez-Bertos A., Arias M., Sánchez-Vizcaíno J.M. First oral vaccination of Eurasian wild boar against African swine fever virus genotype II. Frontiers in Veterinary Science, 2019, 6: 137 (doi: 10.3389/fvets.2019.00137).
- Monteagudo P.L., Lacasta A., López E., Bosch L., Collado J., Pina-Pedrero S., Correa-Fiz F., Accensi F., Navas M.J., Vidal E., Bustos M.J., Rodríguez J.M., Gallei A., Nikolin V., Salas M.L., Rodríguez F. BA71ΔCD2: a new recombinant live attenuated African swine fever virus with cross-protective capabilities. Journal of Virology,2017, 191(21): e01058-17 (doi: 10.1128/JVI.01058-17).
- Lopez E., van Heerden J., Bosch-Camós L., Accensi F., Navas M.J., López-Monteagudo P., Argilaguet J., Gallardo C., Pina-Pedrero S., Salas M.L., Salt J., Rodriguez F. Live attenuated African swine fever viruses as ideal tools to dissect the mechanisms involved in cross-protection. Viruses, 2020, 12(12): 1474 (doi: 10.3390/v12121474).
- Lopez E., Bosch-Camós L., Ramirez-Medina E., Vuono E., Navas M.J., Muñoz M., Accensi F., Zhang J., Alonso U., Argilaguet J., Salas M.L., Anachkov N., Gladue D.P., Borca M.V., Pina-Pedrero S., Rodriguez F. Deletion mutants of the attenuated recombinant ASF virus, BA71ΔCD2, show decreased vaccine efficacy. Viruses,2021, 13(9): 1678 (doi: 10.3390/v13091678).