doi: 10.15389/agrobiology.2019.2.337rus
УДК 619:616.98:578.2:616-078:577.2
Работа выполнена в рамках Государственного задания № 0615-2019-0003.
ПОЛУЧЕНИЕ РЕКОМБИНАНТНОГО НУКЛЕОКАПСИДНОГО БЕЛКА ВИРУСА ЧУМЫ МЕЛКИХ ЖВАЧНЫХ ДЛЯ ПРИМЕНЕНИЯ В СЕРОДИАГНОСТИКЕ
Д.Ю. МОРОЗОВА, А.Р. ИМАТДИНОВ, С.П. ЖИВОДЕРОВ, И.А. ТИТОВ, В.М. ЛЫСКА, А.В. ЛУНИЦИН, А.Д. СЕРЕДА
Чума мелких жвачных (ЧМЖ, peste des petits ruminants) — высококонтагиозная, остро или подостро протекающая вирусная болезнь овец и коз, характеризующаяся язвенными поражениями слизистой оболочки ротовой полости, лихорадкой, поражением лимфоидной системы, развитием пневмонии, геморрагическим гастроэнтеритом (A. Diallo с соавт., 1989; T.M. Ismail с соавт., 1995). Из-за высокой заболеваемости (50-100 %) и смертности (50-90 %) ЧМЖ относится к числу опасных заболеваний, наносящих значительный экономический ущерб (R.A. Kock с соавт., 2015). Возбудитель болезни — вирус ЧМЖ (род Morbillivirus, сем. Paramyxoviridae) антигенно связан с другими морбилливирусами (G. Libeau с соавт., 2014). Степень проявления клинических признаков заболевания зависит от линии вируса ЧМЖ, вида, породы, иммунного статуса животных. Поэтому окончательный диагноз должен быть подтвержден в лаборатории. Для диагностических и мониторинговых серологических исследований предпочтение отдается технологичному и чувствительному методу иммуноферментного анализа (ИФА). Современные средства серодиагностики ЧМЖ разрабатываются на основе вирусоспецифичных рекомбинантных белков, в первую очередь нуклеокапсидного (N) белка (M. Munir с соавт., 2013; Н.В. Вавилова с соавт., 2006.), который по антигенным и иммуногенным характеристикам превосходит остальные белки морбилливирусов (M. Yunus с соавт., 2012). Белок N — наиболее консервативный антиген из протеинов вируса ЧМЖ (M. Muhammad, 2013). Целью представленной работы было получение рекомбинантного нуклеокапсидного белка вируса чумы мелких жвачных для использования в качестве антигена и вирусоспецифичных антисывороток свиней как источника антител для серодиагностики заболевания методом конкурентного ИФА. Была получена конструкция, несущая последовательность фрагмента гена белка N длиной 1530 п.н. в составе экспрессирующего плазмидного вектора pET32a. Скрининг полипептидов методом SDS-PAGE и в иммуноблоттинге выявил клоны Escherichia coli pET32а/N/10, экспрессирующие вирусоспецифичный мажорный полипептид с молекулярной массой 70 кДа. Проведенные нами исследования показали, что оптимальная доза очищенного методом металлохелатной хроматографии рекомбинантного белка N в конкурентном ИФА составила 0,25 мкг на лунку. Соотношение результатов реакции с отрицательной и положительной контрольными сыворотками коз при OD450 составило 11,52. Таким образом, очищенный электрофоретически и иммунохимически чистый рекомбинантный белок N пригоден для применения в конкурентном ИФА для серодиагностики ЧМЖ. Для получения специфичных антисывороток свиньям вводили очищенный вирус ЧМЖ. Титры антител в пробах антисывороток крови свиней в реакции нейтрализации (РН) с вирусом ЧМЖ составили 1:64-1:128. Эти значения были сопоставимы с титрами антител в сыворотках крови у вакцинированных против ЧМЖ овец и коз (А.В. Константинов с соавт., 2017). Однако при конкурентном ИФА антисыворотки свиней оказались менее эффективными по сравнению с положительной козьей сывороткой.
Ключевые слова: чума мелких жвачных животных, морбивирусы, диагностика, рекомбинантные белки, иммуноферментный анализ.
Чума мелких жвачных (ЧМЖ, peste des petits ruminants) — высококонтагиозная, остро или подостро протекающая вирусная болезнь овец и коз, характеризующаяся язвенными поражениями слизистой оболочки ротовой полости, лихорадкой, поражением лимфоидной системы, развитием пневмонии, геморрагическим гастроэнтеритом (1-3). Болезнь была впервые описана на побережье Кот-д’Ивуара в Западной Африке (4), впоследствии ее регистрировали в Африке к югу от Сахары, на Аравийском полуострове и в юго-западной Азии (5). Из-за высокой заболеваемости (50-100 %) и смертности (50-90 %) ЧМЖ считают опасным заболеванием, наносящим значительный экономический ущерб животноводству стран, где оно регистрируется (6, 7).
Возбудитель болезни — вирус ЧМЖ (род Morbillivirus, семейство Paramyxoviridae) антигенно связан с другими морбилливирусами (8, 9). Передача вируса осуществляется при тесном контакте восприимчивых животных (несколько видов газелей, ориксы и белохвостые олени) через секреты и экскреты больных особей. Крупный рогатый скот, буйволы и свиньи могут быть заражены вирусом ЧМЖ естественным образом или экспериментально, но они становятся тупиковыми хозяевами, поскольку у них отсутствуют клинические признаки заболевания и вирус не передается другим животным (5). Из четырех известных в настоящее время линий вируса ЧМЖ одна (линия 4) ограничена Азией, а другие распространены в Африке (10, 11). В основе разделения на линии лежат различия по гену нуклеокапсидного белка, и это более четко отражает географическое происхождение, чем классификация, основанная на изменениях в гене гликопротеина F (12, 13). Степень проявления клинических признаков заболевания зависит от линии вируса ЧМЖ, вида, породы и иммунного статуса животных. Поэтому окончательный диагноз должен подтверждаться лабораторными исследованиями.
Наряду с прямым выявлением генома вируса методом полимеразной цепной реакции в реальном времени (ПЦР-РВ), в лабораторной диагностике для обнаружения вирусоспецифичных антигенов и антител применяют серологические тесты — реакцию нейтрализации (РН) вируса (14), реакцию диффузионной преципитации (15), встречный иммуноэлектроосмофорез, реакцию непрямой иммунофлуоресценции (16, 17), прямой и непрямой варианты иммуноферментного анализа (ИФА) (18), а также конкурентный ИФА на основе моноклональных антител (19-23). Для изучения проб сывороток крови чаще всего используют РН и ИФА. РН — наиболее трудоемкий метод и не подходит для исследования большого количества проб, поскольку предполагает манипуляции с культурами клеток. Поэтому для диагностических и мониторинговых исследований предпочтение отдается технологичному и чувствительному методу ИФА.
Первоначально для обнаружения вирусоспецифичных антител в ИФА в качестве антигена использовали очищенный из лизатов зараженных клеток инактивированный вирус ЧМЖ. Однако специфичность метода была низкой, поскольку в препаратах антигена присутствовали белки культур клеток. Технология конструирования рекомбинантных белков с аффинными метками позволяет очищать их методом хроматографии на металл-хелатных сорбентах, уйти от использования в работе живого вируса ЧМЖ и одновременно повысить специфичность ИФА за счет получения высокоочищенных антигенов. Современные средства серодиагностики ЧМЖ разрабатывают на основе вирусоспецифичных рекомбинантных белков, в первую очередь белка N (24-27), который по антигенным и иммуногенным характеристикам превосходит остальные белки морбилливирусов (28, 29). Другое преимущество белка N состоит в том, что из протеинов вируса ЧМЖ он как антиген наиболее консервативен (30). Следует отметить, что иммунные реакции на белок N, генерируемые восприимчивыми животными в процессе вакцинации или при переболевании, не являются протективными из-за его внутривирионной локализации.
В настоящей работе представлена технология получения продуцента рекомбинантного полноразмерного нуклеокапсидного (N) белка вируса ЧМЖ. Продемонстрирована пригодность очищенного рекомбинантного белка N для выявления антител к вирусу ЧМЖ в конкурентном ИФА. Мы впервые исследовали возможность использования положительных сывороток свиней, иммунизированных вакцинным штаммом вируса ЧМЖ, в качестве контрольных в тест-системах для серодиагностики заболевания. Установлено, что по результатам конкурентного ИФА они уступают положительным сывороткам от коз.
Целью нашей работы было получение рекомбинантного белка N вируса чумы мелких жвачных для использования в качестве антигена и вирусоспецифных антисывороток свиней как источника антител для серодиагностики заболевания методом конкурентного иммуноферментного анализа.
Методика. В работе использовали домашних свиней (Sus scrofa domesticus) породы крупная белая массой 30 кг (n = 2, сектор подготовки животных Федерального исследовательского центра вирусологии и микробиологии), перевиваемую линию клеток почки африканской зеленой мартышки (VERO) и вакцинный штамм 45G37/35-К вируса чумы мелких жвачных (коллекции Федерального исследовательского центра вирусологии и микробиологии).
Накопление вируса ЧМЖ и определение его инфекционной активности осуществляли в культуре клеток VERO в поддерживающей среде Игла МЕМ («ПанЭко», Россия) с 2,5 % сыворотки крови плода крупного рогатого скота соответственно в полистироловых матрасах и 96-луночных планшетах («Costar», Франция). При титровании вирусосодержащий материал последовательно разводили 10-кратно в четырех повторах. Зараженную и контрольную культуры клеток VERO выдерживали при температуре 37 °С в воздушной атмосфере с 5 % СО2, заменяя поддерживающую среду каждые 2-3 сут. Результаты учитывали по цитопатическому действию (ЦПД) в течение 10 сут. Титр вируса рассчитывали по методу Б.А. Кербера в модификации И.П. Ашмарина и выражали в ТЦЦ50/см3 (31).
Реакцию нейтрализации проводили со штаммом 45G37/35-К, используя 2-кратные серийные разведения антисывороток в среде культивирования, смешивая 100 мкл вируса с титром 103 ТЦЦ50/см3 (с получением в каждой лунке 102 ТЦЦ50/см3) и 100 мкл исследуемых разведений образцов антисывороток (по четыре лунки на разведение). Затем из 96-луночных планшетов с культурой клеток VERO удаляли среду культивирования и вносили в лунки полученные смеси вирус—антисыворотка. В качестве контролей использовали лунки с культурой клеток без вируса и зараженные вирусом ЧМЖ (102 ТЦЦ50/см3). Планшеты инкубировали в течение 2 ч при 37 °С в воздушной атмосфере с 5 % СО2, из лунок удаляли жидкую фазу и вносили вместо нее среду культивирования. Титр антител в сыворотках крови животных рассчитывали, исходя из разбавлений, которые нейтрализовали вирус ЧМЖ в 50 % лунок.
Для клонирования ПЦР-продуктов использовали плазмидный вектор pJET1.2 («Thermo Fisher Scientific, Inc.», США), для создания экспрессирующих конструкций — плазмидный вектор pET32a («Novagen», США). Секвенирование осуществляли на генетическом анализаторе Applied Biosystems 3130 («Applied Biosystems, Inc.», США). Возможность применения рекомбинантного белка N в конкурентном ИФА оценивали с использованием части компонентов набора ID Screen® PPR Competition («IDvet», Франция) (далее — набор), предназначенного для определения антител к нуклеопротеину вируса ЧМЖ в сыворотках и плазме крови овец и коз.
Для конкурентного ИФА с полученным антигеном использовали часть компонентов набора ID Screen® PPR Competition («IDvet», Франция).
Электрофоретическое разделение полипептидов проводили в 10 % полиакриламидном геле по U.K. Laemmli (32). Электроперенос полипептидов из геля на нитроцеллюлозную мембрану («Sigma», США) осуществляли в полусухой буферной системе по методу J. Kyhse-Andersen (33). Содержание общего белка в исследуемых пробах определяли по O.H. Lowry с соавт. (34) на фотоколоритметре КФК-2 (ПГК «Гранат», Россия) при λ = 750 нм.
Результаты. Вирус ЧМЖ (штамм 45G37/35-К) накапливали при множественности заражения 10-1 ТЦД50/кл и титровали по ТЦД в культуре клеток VERO. На 4-м пассаже инфекционная активность вируса достигла 104,5-106,5 ТЦЦ50/см3.
Члены семейства парамиксовирусов, как правило, способны репродуцироваться в организме многих видов животных. Например, при инфицировании вирусом чумы собак (ВЧС) погибают или проявляют клинические признаки заболевания большинство наземных плотоядных из семейств Canidae, Мustelidae, Procyonidea, Ursidae, Hyaenidae, Ailuridae, Viverridae. Кошки (Felidae) и свиньи (Suidae) также восприимчивы к инфицированию ВЧС, однако клинические признаки болезни у них не проявляются (35, 36). Титры вируснейтрализующих антител к ВЧС в антисыворотках у экспериментально зараженных свиней сопоставимы с таковыми у переболевших собак и тхорзофреток (37). Вирус ЧМЖ также способен репродуцироваться в организме свиней (38). Мы исследовали возможность получения ЧМЖ-специфической антисыворотки свиней для использования в серодиагностике методом конкурентного ИФА, поскольку иммуноглобулины свиней могут вступать в реакцию с конъюгатами на основе протеина А.
Для снижения вероятности образования антител к компонентам клеток и растворенных в среде культивирования белков из вирусосодержащей суспензии вирионы очищали методом дифференциального центрифугирования. Суспензию вируса ЧМЖ с титром 105 ТЦЦ50/см3 объемом 30 см3 центрифугировали при 5000 g в течение 40 мин. Клеточный дебрис удаляли, надосадок повторно центрифугировали через 20 % (масса/объем) сахарозную подушку при 45000 g в течение 4,5 ч. По окончании надосадок декантировали, а осадок ресуспендировали в 10 см3 ФБР. Очищенные вирионы с титром 105 ТЦЦ50/см3 вводили однократно каждой из двух свиней внутримышечно и интраназально по 2,5 см3. Клинических проявлений заболевания не наблюдали. На 28-е сут после заражения животных обескровливали. Титры антител в пробах антисывороток крови обеих свиней в РН с вирусом ЧМЖ составили 1:64-1:128. Эти значения были сопоставимы с титрами антител в сыворотках крови у вакцинированных против ЧМЖ овец и коз (39).
Из базы данных GenBank (NCBI, https://www.ncbi.nlm.nih.gov/) были взяты нуклеотидные последовательности гена белка N вируса ЧМЖ. В качестве референтной при дизайне праймеров использовали нуклеотидную последовательность штамма Nigeria 75/1 (X74443.2). Праймеры, фланкирующие полноразмерные копии вирусного гена белка N, конструировали с помощью программ Oligo 7, SnapGene v.4.1. Была рассчитана пара праймеров, содержащая на 5'- концах сайты рестрикции SalI и EcoRV. Последовательность Forward-праймера (F-PPRV_N_atc) — 5'-ATCTCGGGTT-CAGGAGGGGCCATCCGGGGG-3', Revers-праймера (R-PPRV_N_SalI) — 5'-GGGTGCGTCGACTTAGCTGAGGAGATCCTTGTCGTTGTAGATCTG-3'. Праймеры были сконструированы для амплификации фрагмента длиной 1530 п.н. (позиции с 4-й по 1518-ю в гене белка N). В олигонуклеотидные праймеры были встроены специфические гексамерные последовательности сайтов рестрикции эндонуклеаз SalI и EcoRV. РНК-экстракцию проводили с использованием TRIzol («Invitrogen Corp.», США). Для синтеза первой цепи кДНК с одноцепочечной РНК-матрицей использовали обратную транскриптазу M-MLV («a-Фермент», Россия). Перед ОТ-ПЦР проводили отжиг праймеров при 70 °С в течение 5 мин.
Нуклеотидную последовательность гена белка N синтезировали в полимеразной цепной реакции (ПЦР) с использованием Pfu-полимеразы («a-Фермент», Россия) на амплификаторе «Терцик» («ДНК-Технологии», Россия) согласно рекомендациям производителя. В качестве матрицы брали полученную ранее кДНК. Программа включала следующие этапы: горячий старт при 95 °С (предварительная денатурация); 3 мин при 95 °С (денатурация), 30 с при 95 °С (отжиг праймеров), 185 c при 68 °С (элонгация) (25 циклов); 5 мин при 72 °С (финальная элонгация). Для очистки ПЦР-продуктов от агарозного геля использовали набор GeneJET Gel Extraction Kit («Thermo Fisher Scientific, Inc.», США) согласно инструкции производителя. Клонировали очищенный от агарозного геля ПЦР-продукт по тупым концам в вектор pJET1.2 с использованием Т4 ДНК лигазы («Thermo Fisher Scientific Inc.», США).
Трансформацию компетентных клеток Escherichia coli (штамм Rosetta 2 pLysS) полученными конструкциями осуществляли по стандартному протоколу с применением теплового шока и последующим высеванием на твердую питательную среду с селективным антибиотиком (ампициллин). Секвенирование положительных в ПЦР плазмидных клонов показало целостность открытой рамки считывания рекомбинантного гена. Следующим этапом стало переклонирование гена белка N в экспрессирующий плазмидный вектор pET32a по сайтам рестрикции EcoRV и XhoI из плазмиды pJET1.2/N/4 (рис. 1). Наличие тиоредоксина в N-концевой области от сайта множественного клонирования вектора pET32a повышало растворимость рекомбинантного белка, а наличие двух полигистидиновых участков позволило проводить очистку экспрессируемого рекомбинантного белка на никелевом сорбенте. Единичные положительные клоны скринировали в ПЦР. Сиквенс положительных клонов подтвердил целостность рамки считывания гена белка N в клонах № 8 и № 10. В дальнейшей работе использовали плазмидную ДНК pET32а/N/10.
Плазмиду pET32а/N/10 трансформировали в E. coli и индуцировали единичные клоны в разных условиях (26, 30 и 37 °С в жидкой питательной среде SOB с добавлением селективного антибиотика ампициллина, исследование проб на 4-й, 8-й и 16-й ч). Установлено, что оптимальными параметрами индукции были температура 26 °С и время 16 ч, индуктор IPTG (изопропил-β-D-1-тиогалактопиранозид) в финальной концентрации 0,4 мМ.
Расчетная молекулярная масса рекомбинантного белка N, слитого с тиоредоксином А и двумя 6His, составила около 70 кДа. Скрининг полипептидов в SDS-PAGE выявил клоны E. coli pET32а/N/10 №№ 3, 5, 6, 8, экспрессирующие мажорный полипептид с молекулярной массой 70 кДа, который отсутствовал в лизате транформированных плазмидой pET32a клеток E. coli без вставки.Рекомбинантный белок N очищали методом металл-хелатной хроматографии в нативных условиях. В качестве сорбента использовали Ni Sepharose™ 6 Fast Flow («GE Healthcare Bio-Sciences Corp», США). Элюировали водными растворами имидазола (50, 100, 250 и 500 мМ), скрининг элюатов проводили методом SDS-PA-GE (рис. 2).
Во фракции элюата с 500 мМ имидазолом конечную концентрацию рекомбинантного белка N после диализа доводили до 2 мг/см3. Его антигенную активность подтверждали методом иммуноблоттинга с антителами ЧМЖ-положительных антисывороток свиней. На блоттограмме был выявлен мажорный вирусоспецифичный полипептид с молекулярной массой 70 кДа. Активность полученного рекомбинантного белка N вируса ЧМЖ и свиных антисывороток против вируса ЧМЖ определяли с использованием компонентов набора ID Screen® PPR Competition («IDvet», Франция).
Вместо стрипов из набора ID Screen® PPR Com-petition («IDvet», Франция) использовали свободные пластиковые стрипы («Eppendorf», Германия), в лунках которых иммобилизовали полученный рекомбинантный белок N (по 50 мкл в 2-кратных нарастающих разведениях от 1:1000 до 1:16000). Положительные и отрицательные сыворотки из набора или положительные и отрицательные сыворотки свиней добавляли в лунки (по 100 мкл) и инкубировали в шейкере 1 ч при 37 °С. Последующие этапы выполняли согласно рекомендациям производителя набора.
Оптимальная концентрация рекомбинантного белка N составила 0,25 мкг на лунку (разведение 1:8000). При этой концентрации соотношение OD450 с отрицательной и положительной сыворотками из набора или с отрицательной и положительной сыворотками крови свиней было максимальным — соответственно 11,52 и 2,22 (рис. 3). Сходный результат (0,20 мкг на лунку) был получен с рекомбинантным белком с молекулярной массой 50 кДа, продуцируемым в клетках E. coli BL21(DE3) pLysS, трансформированных 5´-концевой последовательностью из 838 п.н. гена N-белка (23).
При использовании сенсибилизированных стрипов из набора производителя соотношение OD450 для отрицательной и положительной сывороток из набора составило 16,03. По объективным показателям полученный нами рекомбинантный белок N был пригоден для серодиагностики ЧМЖ методом конкурентного ИФА.
Следует отметить, что показатели оптической плотности в реакции с отрицательной сывороткой крови свиней были сопоставимы с показателями для отрицательной сыворотки из набора производителя, тогда как показатели с положительной сывороткой из набора и положительной антисывороткой от иммунизированных свиней при разведении рекомбинантного белка N 1:8000 различались в 5 раз. Вероятно, это свидетельствует о меньшей концентрации антител к белку N в сыворотках крови свиней, иммунизированных вирусом ЧМЖ, чем в положительной сыворотке из набора, полученной от переболевших коз.
Рекомбинантный белок N вируса ЧМЖ использовали в качестве антигена в непрямом или конкурентном ИФА для серодиагностики в нескольких исследованиях. Ранее этот белок в больших количествах был получен из клеток насекомых, инфицированных рекомбинантным бакуловирусом, содержащим ген нуклеопротеина вируса ЧМЖ (25). Показана активность в конкурентном ИФА укороченного и полноразмерного N-белка вируса ЧМЖ, который получали в E. coli. Оптимальной для обнаружения противовирусных антител была концентрация N-белка 0,16 мкг/на лунку (40). Следует отметить, что для белков, которые не требуют посттрансляционных модификаций, чаще всего используется прокариотическая система E. coli, так как экспрессия в ней проста и есть возможность получать белки в больших количествах. Кроме того, производство рекомбинантных антигенов в бактериях проще и экономичнее по сравнению с другими гетерологичными системами.
Таким образом, нами получен клон Escherichia coli pET32а/N/10, пригодный для использования в качестве продуцента рекомбинантного белка N вируса чумы мелких жвачных (ЧМЖ). Химерный рекомбинантный белок N содержит фрагмент белка тиоредоксина для повышения растворимости в воде и два полигистидиновых участка для обеспечения эффективной очистки на никелевом сорбенте. Очищенный электрофоретически и иммунохимически чистый рекомбинантный белок N пригоден для серодиагностики ЧМЖ с использованием конкурентного иммуноферментного анализа (ИФА). Активные в реакции нейтрализации специфичные анти-ЧМЖ антисыворотки свиней в конкурентном ИФА были менее эффективны, чем положительная козья антисыворотка из набора ID Screen® PPR Competition («IDvet», Франция). На следующем этапе мы планируем получить другие активные компоненты разрабатываемой тест-системы, пригодной для диагностики чумы мелких жвачных методом конкурентного ИФА.
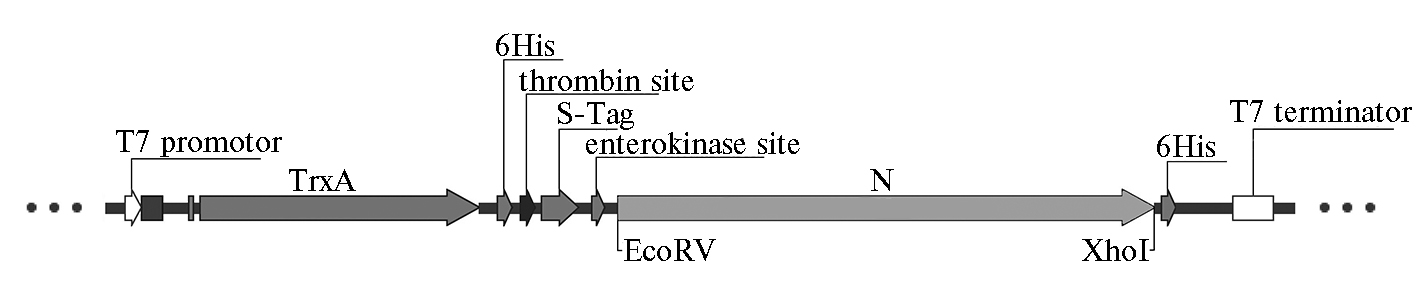
Рис. 1. Схема открытой рамки считывания рекомбинантного гена с клонированной последовательностью фрагмента, кодирующего белок N вируса чумы мелких жвачных: T7 promotor — промотор для РНК-полимеразы бактериофага T7; TrxA — ген тиоредоксина А; 6His — кодирующая последовательность полигистидина; thrombin site — нуклеотидная последовательность, кодирующая сайт расщепления тромбина; S-Tag — кодирующая последовательность олигопептида с N-конеца РНКазы А; enterokinase site — нуклеотидная последовательность, кодирующая сайт расщепления энтерокиназы; EcoRV и XhoI — сайты рестрикции; T7 terminator — терминатор транскрипции для РНК-полимеразы бактериофага T7.
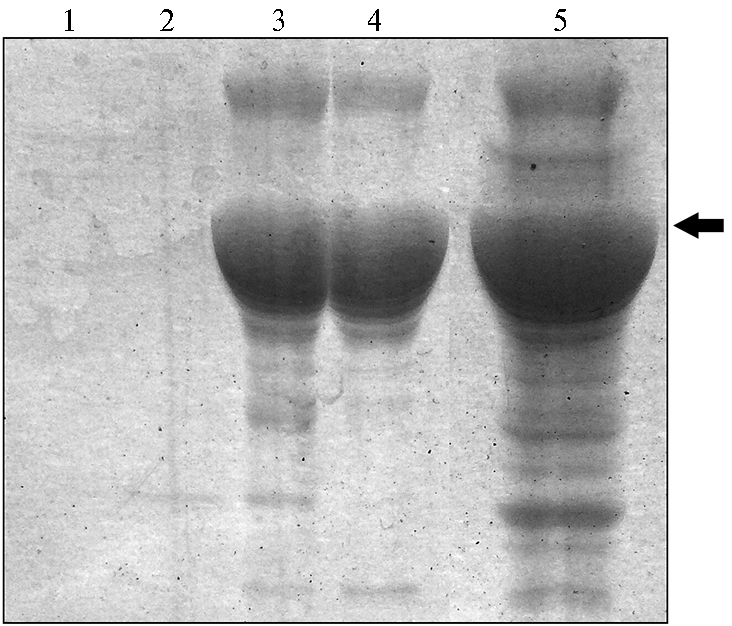
Рис. 2. Электрофореграмма фракций, полученных при хроматографической очистке рекомбинантного белка N вируса чумы мелких жвачных при дискретном повышении концентрации имидазола: 1 — 50 мМ, 2 — 100 мМ, 3 — 250 мМ, 4 — 500 мМ, 5 — исходный лизат индуцированных клеток Escherichia сoli. Стрелка указывает на положение фракции очищенного рекомбинантного белка N.
Рис. 3. Результаты конкурентного иммуноферментного анализа с иммобилизованным на лунках стрипов («Ep-pendorf», Германия) рекомбинантным белком N вируса чумы мелких жвачных: 1 и 2 — положительная и отрицательная сыворотки из набора ID Screen® PPR Competition («IDvet», Франция), 3 и 4 — сыворотки крови свиней (соответственно положительная от экспериментально иммунизированного вирусом ЧМЖ животного и отрицательная от интактного животного).
ЛИТЕРАТУРА
- Gibbs E.P.J., Taylor W.P., Lawman M.J.P., Bryant J. Classification of peste des petits ruminants virus as the fourth member of the genus Morbillivirus. Intervirology, 1979, 11: 268-274 (doi: 10.1159/000149044).
- Diallo A., Barrett T., Barbron M., Shaila M.S., Lefevre P.C. Differentiation of rinderpest and Pestedes-petits-ruminants viruses using specific cDNA clones. J. Virol. Methods, 1989, 23(2): 127-136 (doi: 10.1016/0166-0934(89)90126-2).
- Ismail T.M., Yamanaka M.K., Saliki J.T., El-Kholy A., Mebus C., Yilma T. Cloning and expression of the nucleoprotein of Peste-des-petits-ruminants virus in baculovirus for use in serological diagnosis. Virology, 1995, 208(2): 776-778 (doi: 10.1006/viro.1995.1210).
- Gargadennec L., Lalanne A. La peste des petits ruminants. Bulletin des Services Zootechniques et des Epizooties de l’Afrique Occidentale Francaise, 1942, 5(1): 16-21.
- Taylor W.P. The distribution and epidemiology of peste des petits ruminants. Prev. Vet. Med.,1984, 2(1-4): 157-166 (doi: 10.1016/0167-5877(84)90059-X).
- Kock R.A., Orynbayev M.B., Sultankulova K.T., Strochkov V.M., Omarova Z.D., Shalgynbayev E.K., Rametov N.M., Sansyzbay A.R., Parida S. Detection and genetic characterization of lineage IV peste des petits ruminant virus in Kazakhstan. Transbound. Emerg. Dis., 2015, 62(5): 470-479 (doi: 10.1111/tbed.12398).
- Abu-Elzein E.M.E., Hassanien M.M., Al-Afaleq A.I., Abd-Elhadi M.A., Housain F.M.I. Isolation of Peste-des-petits-ruminants virus from goats in Saudi Arabia. Vet. Rec., 1990, 127(12): 309-310.
- Desselberger U. Virus taxonomy: classification and nomenclature of viruses. Seventh report of the International Committee on Taxonomy of Viruses, edited by M.H.V. van Regenmortel, C.M. Fauquet, D.H.L. Bishop, E. Carstens, M.K. Estes, S. Lemon, J. Maniloff, M.A. Mayo, D.J. McGeoch, C.R. Pringle, R. Wickner. Virology Division, International Union of Microbiological Societies, Academic Press, Inc., San Diego, Calif., 2000. Virus Res., 2002, 83(1): 221-222 (doi: 10.1016/S0168-1702(01)00352-5).
- Libeau G., Diallo A., Parida S. Evolutionary genetics underlying the spread of peste des petits ruminants virus. Animal Frontiers, 2014, 4(1): 14-20 (doi: 10.2527/af.2014-0003).
- Shaila M.S., Shamaki D., Forsyth M.A., Diallo A., Groatley L., Kitching R.P., Barrett T. Geographic distribution and epidemiology of Peste-des-petits-ruminants viruses. Virus Res., 1996, 43(2): 149-153 (doi: 10.1016/0168-1702(96)01312-3).
- Dhar P., Sreenivasa B.P., Barrett T., Corteyn M., Singh R.P., Bandyopadhyay S.K. Recent epidemiology of Peste-des-petits-ruminants virus (PPRV). Vet. Microbiol., 2002, 88(2): 153-159 (doi: 10.1016/S0378-1135(02)00102-5).
- Diallo A., Minet C., Le Goff C., Berhe G., Albina E., Libeau G., Barrett T. The threat of peste des petits ruminants: progress in vaccine development for disease control. Vaccine, 2007, 25(30): 5591-5597 (doi: 10.1016/j.vaccine.2007.02.013).
- Kwiatek O., Minet C., Grillet C., Hurard C., Carlsson E., Karimov B., Albina E., Diallo A., Libeau G. Peste des petits ruminants (PPR) outbreak in Tajikistan. J. Comp. Pathol., 2007, 136(2-3): 111-119 (doi: 10.1016/j.jcpa.2006.12.002).
- Rossiter P.B., Jessett D.M., Taylor W.P. Microneutralisation systems for use with different strains of peste des petits ruminants virus and rinderpest virus. Trop. Anim. Health. Pro., 1985, 17(2): 75-81 (doi: 10.1007/BF02360775).
- Durojaiye O.A. Precipitating antibody in sera of goats naturally affected with peste des petits ruminants. Trop. Anim. Health. Pro., 1982, 14(2): 98-100 (doi: 10.1007/BF02282591).
- Sumption K.J., Aradom G., Libeau G., Wilsmore A.J. Detection of peste des petits ruminants virus antigen in conjunctival smears of goats by indirect immunofluorescence. Vet. Rec., 1998, 142(16): 421-424 (doi: 10.1136/vr.142.16.421).
- Durojaiye O.A., Taylor W.P. Application of countercurrent immuno-electro-osmophoresis to the serology of peste des petits ruminants. Revue d'élevage et de Médecine Vétérinaire des Pays Tropicaux, 1984, 37(3): 272-276.
- Obi T.U., McCullough K.C., Taylor W.P. The production of peste des petits ruminants hyperimmune sera in rabbits and their application in virus diagnosis. Zoonoses and Public Health, 1990, 37(1-10): 345-352 (doi: 10.1111/j.1439-0450.1990.tb01068.x).
- Anderson J., McKay J.A., Butcher R.N. The use of monoclonal antibodies in competition ELISA for detection of antibodies to rinderpest and peste des petits ruminants viruses. Proc. of a Final Research Co-ordination Meeting of the FAO/IAEA/SIDA/OAU/IBAR/PARC co-ordinated research programme organized by the joint FAO/IAEA division of nuclear techniques in food and agriculture and held in Bingerville. Côte d'Ivoire, 1990, v. 1: 43-53.
- Saliki J.T., Libeau G., House J.A., Mebus C.A., Dubovi E.J. A monoclonal antibody-based blocking enzyme-linked immunosorbent assay for specific detection and titration of peste-des-petits-ruminants virus antibody in caprine and ovine sera. J. Clin. Microbiol., 1993, 31(5): 1075-1082.
- Balamurugan V., Singh R.P., Saravanan P., Sen A., Sarkar J., Sahay B., Rasool T.J., Singh R.K. Development of an indirect ELISA for the detection of antibodies against Peste-des-petits-ruminants virus in small ruminants. Vet. Res. Commun., 2007, 31(3): 355-364 (doi: 10.1007/s11259-006-3442-x).
- Singh R.P., Sreenivasa B.P., Dhar P., Shah L.C., Bandyopadhyay S.K. Development of a monoclonal antibody based competitive ELISA for detection and titration of antibodies to Peste-des-petits-ruminants (PPR) virus. Vet. Microbiol., 2004, 98(1): 3-15 (doi: 10.1016/j.vetmic.2003.07.007).
- Balamurugan V., Roy M., Sowjanyakumari S., Abraham S., Apsana R., Nagalingam M., Hemadri D., Rahman H. Development of recombinant nucleocapsid protein based indirect ELISA for serodiagnosis of Peste des Petits ruminants in sheep and goats. Adv. Anim. Vet. Sci., 2016, 4(6): 301-310 (doi: 10.14737/journal.aavs/2016/4.6.301.310).
- Diallo A., Barrett T., Barbron M., Meyer G., Lefevre P.C. Cloning of the nucleocapsid protein gene of peste-des-petits-ruminants virus: relationship to other morbilliviruses. J. Gen.Virol., 1994, 75(1): 233-237 (doi: 10.1099/0022-1317-75-1-233).
- Libeau G., Prehaud C., Lancelot R., Colas F., Guerre L., Bishop D.H., Diallo A. Development of a competitive ELISA for detecting antibodies to the peste des petits ruminants virus using a recombinant nucleoprotein. Res. Vet. Sci., 1995, 58(1): 50-55 (doi: 10.1016/0034-5288(95)90088-8).
- Munir M., Zohari S., Berg M. Current advances in molecular diagnosis and vaccines for peste des petits ruminants. Molecular Biology and Pathogenesis of Peste des Petits Ruminants Virus, 2013, 1: 105-133 (doi: 10.1007/978-3-642-31451-3_6).
- Вавилова Н.В., Щербаков А.В. Применение рекомбинантного нуклеокапсидного белка в непрямом варианте ИФА для выявления антител к вирусу чумы мелких жвачных. Ветеринарная патология, 2006, 4: 76-78.
- Lefevre P.C., Diallo A., Schenkel F., Hussein S., Staak G. Serological evidence of peste des petits ruminants in Jordan. Vet. Rec., 1991, 128: 110 (doi: 10.1136/vr.128.5.110).
- Yunus M., Shaila M.S. Establishment of an in vitro transcription system for peste des petits ruminant virus. Virol. J., 2012, 9: 302 (doi: 10.1186/1743-422X-9-302).
- Muhammad M. Peste des petits ruminants virus. In: Mononegaviruses of veterinary importance. Volume I: Pathobiology and molecular diagnosis /M. Munir (ed.). CABI Publishers, 2013: 65-98 (doi: 10.1079/9781780641799.0000).
- Ашмарин И.П., Васильев Н.Н., Амбросов В.А. Быстрые методы статистической обработки и планирование экспериментов. Л., 1974.
- Laemmli U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 1970, 227: 680-685 (doi: 10.1038/227680a0).
- Kyhse-Andersen J. Electroblotting of multiple gels: a simple apparatus without buffer tank for rapid transfer of proteins from polyacrylamide to nitrocellulose. J. Biochem. Bioph. Meth., 1984, 10(3/4): 203-209 (doi: 10.1016/0165-022X(84)90040-X).
- Lowry O.H., Rosebrough N.J., Farr A.L., Randall R.J. Protein measurement with the Folin Phenol Reagent. J. Biol. Chem., 1951, 193(1): 265-275.
- Appel M.J.G., Summers B.A. Pathogenicity of morbilliviruses for terrestrial carnivores. Vet. Microbiol., 1995, 44(2): 187-191 (doi: 10.1016/0378-1135(95)00011-X).
- Appel M.J.G., Sheffy B.E., Percy D.H., Gaskin J.M. Canine distemper virus in domesticated cats and pigs. Am. J. Vet. Res., 1974, 35: 803-806.
- Середа А.Д., Ногина И.В. Иммуносупрессия у собак и свиней, инфицированных вирусом чумы собак. Вопросы вирусологии, 2011, 5: 44-47.
- Taylor W.P. The distribution and epidemiology of peste des petits ruminants. Prev. Vet. Med., 1984, 2: 157-166 (doi: 10.1016/0167-5877(84)90059-X).
- Константинов А.В., Старов С.К., Диев В.И., Мороз Н.В., Курненкова Е.В., Басова Д.К., Кононов А.В., Федосеев К.Ю., Мельников В.П. Антигенная и протективная активность ассоциированной вирусвакцины против оспы овец, оспы коз и чумы мелких жвачных. Ветеринария сегодня, 2017, 3: 28-32.
- Yadav V., Balamurugan V., Bhanuprakash V., Sen A., Bhanot V., Venkatesan G., Riyesh T., Singh R.K. Expression of Peste des Petits ruminants virus nucleocapsid protein in prokaryotic system and its potential use as a diagnostic antigen or immunogen. J. Virol. Methods, 2009, 162(1-2): 56-63 (doi: 10.1016/j.jviromet.2009.07.014).