doi: 10.15389/agrobiology.2019.2.386rus
УДК 636.2:619:57.088.1
ДИНАМИКА АМИНОКИСЛОТНОГО СОСТАВА МОЛОКА У BIV- И BLV-BIV-ИНФИЦИРОВАННЫХ КОРОВ ПРИ ХРАНЕНИИ
Е.С. КРАСНИКОВА, О.С. ЛАРИОНОВА, А.В. БАННИКОВА, А.В. ЕВТЕЕВ, Г.Х. УТАНОВА
Лейкоз и вирусный иммунодефицит крупного рогатого скота относятся к разряду труднодиагностируемых инфекционных заболеваний сельскохозяйственных животных, что повышает вероятности вирусоносительства. Для диагностики иммунодефицита крупного рогатого скота не существует законодательно утвержденных норм, поэтому в хозяйствах молоко от больных коров может инфицировать товарную продукцию от здоровых животных. Часто возбудитель вирусного иммунодефицита (BIV) обнаруживается у животных, инфицированных возбудителем энзоотического лейкоза (BLV). В представленной работе мы впервые выявили, что молоко инфицированных ретровирусами коров имеет измененный аминокислотный баланс казеиновой фракции, а также низкую стабильность белка при хранении в условии холодильника. Целью наших исследований был сравнительный анализ и оценка стабильности аминокислотного состава молока от коров, зараженных возбудителями иммунодефицита и иммунодефицита/лейкоза, и животных, интактных по этим вирусам. Материалом для исследования служили 6 проб цельного молока от коров (Bos taurus taurus) черно-пестрой породы (2,8-6,7 лет), зараженных вирусом бычьего иммунодефицита, и коров с BLV-BIV микст-инфекцией. Анализ аминокислотного состава белковых фракций выполняли с использованием системы капиллярного электрофореза Капель 105М (ООО «Люмэкс-Маркетинг», Россия). Молоко исследовали на 1-е, 3-и и 6-е сут хранения в холодильнике при 4 °С. Для сравнительного анализа использовали общепринятые стандартные данные по аминокислотному балансу казеиновой фракции молока здоровых коров. Результаты наших исследований свидетельствуют о том, что молоко у BIV и BLV-BIV инфицированных коров по аминокислотному составу значительно отличается от молока здоровых коров. Массовая доля таких незаменимых и условно-незаменимых аминокислот, как метионин, глутамин, гистидин и глицин, в молоке у инфицированных коров снижалась в 3-4 раза. Содержание аргинина, лизина, пролина и аспарагина было близко к норме, тирозин переходил в разряд лимитирующих на фоне недостатка фенилаланина, содержание которого было в 6 раз ниже нормы. Также наблюдался относительный избыток треонина и серина. Массовая доля лизина, валина, лейцина-изолейцина и глутамина оказалась соответственно в 9; 13; 17,5 и 22 раза ниже, чем в молоке здоровых коров. Соотношение лейцина-изолейцина и валина в молоке инфицированных коров было примерно 1:1, тогда как в молоке здоровых — 2,5:1. Молоко, полученное от BIV и BLV-BIV инфицированных животных, характеризовалось выраженной динамичностью аминокислотного состава при хранении в условиях холодильника. На 6-е сут хранения при 4 °С незаменимые аминокислоты аргинин, гистидин и метионин в нем не обнаруживались. Массовая доля тирозина возрастала на 3-и сут хранения и снижалась к 6-м сут. Причем такая динамика была более выражена в молоке у BIV-инфицированных коров. Содержание фенилаланина, лейцина-изолейцина, валина, серина и глицина в молоке у BIV- и BLV-BIV-инфицированных животных возрастало на 6-е сут соответственно на 27,1 и 2,4 %; 22,3 и 8,9 %; 43,9 и 37,2 %; 25,0 и 27,3 % и 0 и 60 %. При этом массовая доля пролина и треонина снижалась к 6-м сут на 10,6 и 13,7 %; 5,3 и 0 %. Массовая доля аланина в молоке у BIV-инфицированных коров к 6-м сут возрастала на 37,5 %, а в молоке BLV-BIV-инфицированных животных, напротив, снижалась на 17,8 %. Дисбаланс и нестабильность совокупности аминокислот казеиновой фракции молока, полученного от BIV-BLV- и BIV-инфицированных коров, свидетельствуют о развитии в нем неконтролируемых процессов.
Ключевые слова: коровье молоко, молочный белок, аминокислоты, вирусный иммунодефицит, лейкоз.
По сведениям Института питания РАМН, недостаток пищевого белка в нашей стране ежегодно превышает 1,6 млн т. Общий дефицит протеина на Земле оценивают в 10-25 млн т в год (1). При этом на каждого жителя планеты приходится около 60 г белка в сутки. Исследования баланса азота в организме выявили, что оптимальная норма потребления протеина для взрослого здорового человека составляет 0,8 г на 1 кг массы тела в сутки. Этот показатель может варьировать в зависимости от умственной и физической нагрузки, пола, возраста и физиологического состояния. Так, для молодого здорового мужчины при минимальной физической нагрузке нормальной протеиновой диетой считается 88 г белка в сутки (2). Наряду с количеством, качество белка, в том числе сбалансированное соотношение аминокислот, также определяет питательную ценность протеина для человеческого организма. Белки животного происхождения (в отличие от растительных) содержат в сбалансированном соотношении все необходимые аминокислоты. Незаменимых аминокислот в растительной продукции существенно меньше, чем в продукции животного происхождения. Даже оптимальный по аминокислотному составу соевый белок не содержит незаменимую аминокислоту метионин в достаточном количестве (3).
Коровье молоко в большинстве случаев присутствует в питании человека любого возраста, его аминокислотный состав полноценен и сбалансирован. Протеины коровьего молока усваиваются организмом на 96-98 %. Установлено, что молочные протеины стабилизируют кровяное давление у людей, страдающих гипертонией. Около 80 % молочных белков составляет казеиновая фракция, ее содержание в коровьем молоке в среднем колеблется от 2,1 до 2,9 % (4).
Ферментативный гидролиз молочного белка начинается в процессе хранения сырого молока. Его осуществляют протеолитические ферменты молочной микрофлоры и внесенной закваски, а также фермент плазмин (5, 6). Среди продуктов ферментативного гидролиза молока превалируют пептиды, при этом фракция свободных аминокислот относительно невелика. Пептиды молока стимулируют секрецию инсулина и гормона роста, оказывают положительное влияние на усвоение эссенциальных микронутриентов, в том числе ионов кальция (7). Формирующиеся в результате молочного гидролиза пептиды, помимо высокой усвояемости, задерживают десквамацию клеток слизистой оболочки кишечника и стимулируют выработку эндогенных энзимов (8). Низкомолекулярные молочные пептиды быстро проникают в мышцы и вызывают сильный инсулинотропный эффект. Восполняя энергетические запасы организма, они насыщают мышцы аминокислотами, что приводит к стимуляции синтеза мышечного белка. Микрофракции низкомолекулярных сывороточных беков — гликомакропептидов снижают риск развития вирусных заболеваний, благоприятно влияют на пищеварение, усвоение кальция и белка, стимулируют развитие нормальной микрофлоры кишечника (9).
Неуправляемый и непредсказуемый протеолиз под действием протеолитических ферментов вторичной микрофлоры, в том числе гнилостной, может стать причиной формирования в сыром молоке различных соединений с посторонним запахом и вкусом. Свободные аминокислоты при этом подвергаются ферментации, в результате которой происходят реакции их переаминирования, дезаминирования и декарбоксилирования с образованием кетокислот, оксикислот, карбоновых кислот и альдегидов (10). Следствием глубокого распада молочных белков, индуцируемого посторонними микроорганизмами, становится образование свободных аминокислот, которые, в свою очередь, влияют на органолептические свойства молока. Например, аланин, глицин, пролин и серин имеют сладкий вкус, аспарагиновая и глутаминовая кислоты — кислый, аргинин, лейцин, гистидин и триптофан — горький, метионин и цистин — сернистый, треонин, валин и фенилаланин — сладко-горький (11). Помимо этого, свободные аминокислоты конкурируют за аминокислотные каналы между собой, а большое количество свободных аминокислот вызывают расстройства желудочно-кишечного тракта (12).
Качество коровьего молока, его биологическая и питательная ценность зависят не только породы, условий содержания и кормления животных, но и от состояния их здоровья. Многие заболевания, в том числе хронические инфекционные, могут приводить к стойкому снижению удоев у коров и ухудшению качества получаемой от них продукции. Установлено, что у молочных коров, инфицированных вирусом энзоотического лейкоза крупного рогатого скота (ВЭЛ КРС, bovine leukemia virus, BLV), количество соматических клеток в молоке может составлять (4,9- 5,2)×105/см3, а удои снижаются на 13,3-15,5 % (13), что наносит животноводству значительный экономический ущерб (14). Количество ферритина и железа в молоке у BLV-инфицированных коров снижается, а сывороточного ферритина — возрастает, что служит маркером воспалительных и злокачественных процессов (15).
BLV-инфекция сопровождается снижением общей резистентности организма животного и угнетает иммунитет в молочной железе. Это выражается в уменьшении эффективности фагоцитоза, следствием чего становится значительная бактериальная обсемененность молока инфицированных коров — до 2×107±4×102 КОЕ/мл (16). В молоке у BLV-инфицированных коров доля апоптических В-клеток и экспрессия нейтрофилами CD44 значительно снижены, особенно у животных с персистирующим лимфоцитозом (17). Также наблюдается уменьшение количества общего белка и аминокислот, в том числе пяти незаменимых (изолейцина, триптофана, метионина, лейцина и фенилаланина). Электрофоретический анализ белков молока показывает увеличение фракций протеозо-пептонов, лактальбуминов и иммуноглобулинов на фоне уменьшения фракции альбумина сыворотки крови (18).
Лейкоз часто протекает сочетано с другой хронической инфекцией КРС — вирусным иммунодефицитом (ВИ КРС). Вирус бычьего иммунодефицита (bovine immunodeficiency virus, BIV), как и вирус лейкоза, тропен к лимфоцитам и вызывает стойкие нарушения в иммунной системе. ВИ КРС — относительно новое заболевание, однако его распространение в неблагополучных стадах иногда выше, чем ЭЛ КРС (19, 20).
Наши предыдущие исследования белкового состава молока коров при BIV- и BLV-инфекциях показали, что он отличается от белкового состава у здоровых животных (16). У коров с дефектной иммунной системой может изменяться профиль молочных протеинов и пептидов, соотношение свободных и связанных аминокислот. В настоящее время в литературе нет данных о том, как меняется аминокислотный состав молока коров при BIV-BLV коинфекции и BIV-инфекции в условиях его хранения в холодильнике. Эта информация представляет значительный теоретический интерес и практическую значимость, поскольку позволит обосновать биохимические преобразования в молоке и судить о его биологической ценности. Наиболее информативны, на наш взгляд, данные по казеиновой фракции молока, поскольку аминокислотный состав этого белка не меняется даже при гидролизе.
В представленной работе мы впервые выявили, что молоко инфицированных ретровирусами коров имеет измененный аминокислотный баланс казеиновой фракции, а также низкую стабильность белка при хранении в холодильнике при 4 °С.
Нашей целью было сравнение и оценка стабильности аминокислотного состава казеиновой фракции молока от коров, инфицированных вирусом бычьего иммунодефицита и коинфицированных вирусами бычьего лейкоза и бычьего иммунодефицита, и молока здоровых животных.
Методика. Материалом служили 6 проб цельного молока, полученного от молочных коров (Bos taurus taurus) черно-пестрой породы (2,8-6,7 лет) с BLV-BIV коинфекцией (n = 3) и коров, инфицированных вирусом бычьего иммунодефицита (n = 3).
Выделение и очистку нуклеиновых кислот выполняли с применением набора ДНК Сорб В («ИнтерЛабСервис», Россия). Присутствие или отсутствие провирусов BLV и BIV в стабилизированной крови коров устанавливали методом классической ПЦР с применением набора ЛЕЙКОЗ («ИнтерЛабСервис», Россия) и собственным разработанным методом мультиплексной ПЦР (патент РФ № 2615465) с использованием оригинальных праймеров (синтезированы в ЗАО «Синтол», Россия), набора ПЦР-Микс и буфера для нанесения (НПФ «Литех», Россия). Амплификацию проводили в термоциклере Т100 («Bio-Rad», США). Детекцию продуктов амплификации осуществляли методом горизонтального электрофореза в 2 % агарозном геле с добавлением 0,5 мкг/л бромистого этидия с применением набора ЭФ («ИнтерЛабСервис», Россия) при стандартных условиях с фоторегистрацией полученных результатов. Учет результатов выполняли на оборудовании GelDoc XP PLUS («Bio-Rad», США). Коммерческие наборы и оборудование использовали согласно рекомендациям разработчиков и производителей.
У коров не отмечали клинически выраженного мастита, а органолептические качества молока оставались в пределах нормы.
При пробоподготовке цельное молоко (10 мл) центрифугировали 30 мин при 10 тыс. об/мин и температуре 5 °С. Разделяли осадок и верхнюю фракцию липидов, после чего в обезжиренное молоко по каплям вносили соляную кислоту при 37 °С, постоянно перемешивая, до достижения рН 4,5, затем перемешивание продолжали на протяжении 30 мин. Полученную субстанцию центрифугировали в течение 10 мин при 5 тыс. об/мин и 25 °С и отделяли надосадочную фракцию; рН осадка доводили до 6,5 с помощью 1 М раствора гидроксида натрия.
Аминокислотный состав белковых фракций определяли с использованием системы капиллярного электрофореза Капель 105М (ООО «Люмэкс-Маркетинг», Россия; методика М-04-38-2009 изменениями № 1 от 01.02.2010 в соответствии с рекомендациями производителя). Молоко исследовали на 1-е, 3-и и 6-е сут хранения в холодильнике при 4 °С.
Для сравнительного анализа использовали общепринятые стандартные данные по аминокислотному балансу казеиновой фракции молока здоровых коров.
Полученные данные обрабатывали в программе Statistica 8.0 («StatSoft, Inc.», США). Достоверность различий оценивали с помощью U-критерия Манна-Уитни, за достоверные принимали различия на уровне 95 % (p < 0,05). В статье представлены средние значения массовой доли основных аминокислот казеиновой фракции молока (M) и среднеквадратические отклонения (±σ).
Результаты. Изучение стабильности аминокислотного состава молока инфицированных коров в динамике свидетельствует о достаточно высокой вариабельности содержания аминокислот (рис.). Это может быть следствием активных биохимических процессов в молоке.
Наиболее динамичными были показатели массовой доли аргинина, метионина и гистидина: к 6-м сут хранения молока в условиях холодильника она стала ниже предела детекции. Содержание остальных аминокислот либо было относительно стабильным (треонин, глутамин, аспарагин, фенилаланин, пролин, лейцин-изолейцин), либо колебалось в более значительных пределах (серин, глицин, аланин, лизин, валин, тирозин, глутамин). Изменения показателя для многих аминокислот носили случайный характер. Их количество либо возрастало на 3-и сут (тирозин) или 6-е сут (фенилаланин, валин, серин, глицин) хранения молока, либо со временем снижалось (треонин, пролин).
Известно, что аминокислотный состав казеиновой фракции молока коров наиболее стабилен. У коров черно-пестрой породы он колеблется в незначительных пределах (0,01-0,15 %) по фазам лактации в зависимости от концентрации аминокислоты (21).
Таблица иллюстрирует содержание аминокислот в молоке у BLV-BIV- и BIV-инфицированных коров в динамике относительно средних справочных значений у здоровых коров (22). Содержание аминокислот в коровьем молоке при BIV-инфекции было в среднем в 5-20 раз меньше, чем в молоке здоровых животных (см. табл.), а большинство незаменимых аминокислот, таких как метионин, глицин, гистидин, глутамин, присутствовали в очень небольшом количестве. Значительно изменялся аминокислотный баланс молока. Содержание тирозина, лейцина, серина в молоке у больных коров оказалось во много раз ниже, чем у здоровых, в то время как содержание аргинина, лизина, пролина и аспарагина было ближе к норме.
Суточная норма потребления аминокислот для человека, как правило, исчисляется в зависимости от диетических особенностей питания и индивидуальных потребностей организма. При определении баланса аминокислот лимитирующим считается содержание 11 незаменимых аминокислот (лизина, метионина, триптофана, валина, аргинина, гистидина, лейцина, изолейцина, фенилаланина треонина, и глицина). Другие 10 аминокислот считаются заменимыми (аспарагин, аланин, аспартат, глутамат, глицин, глутамин, серин, пролин, тирозин и цистеин). Однако это условное разделение (23). В частности, в исследованном нами молоке от BIV-инфицированных коров аргинин и гистидин были в дефиците, в то время как содержащийся в достаточном количестве тирозин переходил в разряд лимитирующих, поскольку отмечался недостаток фенилаланина.
У некоторых людей возникает потребность в повышении суточной нормы потребления аминокислот. В то же время избыток аминокислот нежелателен, поскольку для их утилизации в организме требуется дополнительная энергия (24). Аминокислота, представленная в избытке, может занять в обмене веществ место недостающей, вследствие чего развиваются костные патологии, токсикоз и увеличение отложения жира в организме. В молоке у BIV-инфицированных коров мы выявили относительный избыток серина и треонина. Подобную тенденцию отмечали и у коров с коинфекцией BIV-BLV (см. табл.).
Чтобы избежать нарушений в обмене веществ, необходимо придерживаться индекса аминокислотной сбалансированности, то есть соотношения содержания аминокислот в пище. Это связано с тем, что в процессе метаболизма между сходными по структуре аминокислотами наблюдается антагонизм. В частности, между треонином и триптофаном, аргинином и лизином, лейцином и изолейцином (25). Полученные нами данные свидетельствуют о конфликте баланса между лейцином-изолейцином и валином, поскольку их массовая доля в молоке у здоровых коров должна иметь соотношение близкое к 2,5:1 (22), в то время как у инфицированных коров это соотношение со временем хранения молока стремилось к 1:1. Изменение метаболизма аминокислот, в частности их относительных пропорций, приводит к изменению гомеостаза всего организма. Среди эссенциальных аминокислот значительное внимание уделяется лейцину, который необходим для стимуляции синтеза белка и ингибирования протеолиза в клетках. Функции валина в организме менее разнообразны (4, 25).
Ряд зарубежных авторов предлагают с целью ранней диагностики субклинических маститов у коров исследовать аминокислотный состав молока методом высокоэффективной жидкостной хроматографии (26) и даже указывают индикаторные показатели при наиболее распространенном стафилококковом мастите (27). Однако подобные сообщения в литературе встречаются достаточно редко, следовательно, это направление исследований в ветеринарной медицине актуально и перспективно.
Таким образом, содержание аминокислот в молоке, полученном от коров при инфицировании вирусом иммунодефицита и коинфекции вирусами бычьего лейкоза и иммунодефицита, было в 5-20 раз меньше, чем в молоке от здоровых животных. Массовая доля таких незаменимых и условно незаменимых аминокислот, как метионин, глутамин, гистидин и глицин, снижалась наиболее значительно. Содержание тирозина, лейцина, серина в молоке у инфицированных коров было во много раз меньше, чем в молоке здоровых. Количество аргинина, лизина, пролина и аспарагина снижалось менее значительно, при этом тирозин переходил в разряд лимитирующих на фоне недостатка фенилаланина. Наблюдался избыток относительной концентрации треонина и серина. Массовая доля лейцина-изолейцина и валина в молоке у здоровых и инфицированных коров была не пропорциональна, что усугублялось со временем хранения молока. Молоко от BIV и BLV-BIV-инфицированных коров характеризовалось выраженной динамичностью аминокислотного состава при хранении в холодильнике. На 6-е сут хранения в холодильнике незаменимые аминокислоты аргинин, гистидин и метионин в молоке у инфицированных коров не обнаруживались. Массовая доля тирозина возрастала на 3-и сут хранения, затем снижалась. Массовая доля фенилаланина, лейцина, изолейцина, валина, серина и глицина возрастала на 6-е сут хранения, пролина и треонина — снижалась. Дисбаланс и нестабильность аминокислотного состава молока у BLV-BIV- и BIV-инфицированных коров свидетельствуют о развитии в нем неконтролируемых процессов. Важно своевременно выводить из стада инфицированных животных и совершенствовать систему мониторинга здоровья крупного рогатого скота.
Динамика относительного содержания аминокислот (доля от нормы, %) в молоке коров черно-пестрой породы, инфицированных BIV (bovine immunodeficiency virus) или коинфицированных BLV (bovine leukemia virus) и BIV, после хранения в холодильнике при 4 °С (M±σ) |
||||||
Аминокислота |
BIV |
BLV-BIV |
||||
1-е сут |
3-и сут |
6-е сут |
1-е сут |
3-и сут |
6-е сут |
|
Аргинин |
8,8±0,9 |
5,6±0,6* |
-* |
8,9±0,9 |
6,6±0,6* |
-* |
Лизин |
5,8±0,6 |
6,6±0,6* |
6,8±0,7* |
6,1±0,6 |
6,6±0,7 |
6,5±0,6 |
Фенилаланин |
4,2±0,4 |
4,6±0,4 |
5,8±0,6* |
4,9±0,5 |
4,6±0,4* |
5,0±0,5* |
Гистидин |
3,7±0,3 |
2,1±0,2* |
-* |
2,4±0,2 |
2,1±0,2* |
-* |
Лейцин + Изолейцин |
2,9±0,3 |
3,5±0,3* |
3,7±0,3* |
3,3±0,3 |
3,5±0,3 |
3,6±0,3 |
Метионин |
4,5±0,5 |
1,4±0,1* |
-* |
8,6±0,8 |
1,9±0,2* |
-* |
Валин |
4,3±0,4 |
4,4±0,4 |
7,7±0,7* |
3,8±0,3 |
5,9±0,6* |
6,0±0,6* |
Пролин |
7,2±0,7 |
6,9±0,7 |
6,5±0,6* |
7,7±0,7 |
6,0±0,6* |
6,7±0,7* |
Треонин |
5,3±0,5 |
4,2±0,4* |
5,1±0,5 |
5,4±0,5 |
4,4±0,4* |
4,6±0,4* |
Серин |
3,3±0,3 |
3,2±0,3 |
4,4±0,4* |
3,3±0,3 |
2,9±0,3* |
4,5±0,4* |
Аланин |
4,8±0,4 |
5,1±0,5 |
7,8±0,8* |
6,7±0,6 |
4,9±0,5* |
5,5±0,5* |
Глицин |
4,3±0,4 |
3,1±0,3* |
4,3±0,4 |
3,1±0,3 |
2,9±0,3 |
5,0±0,5* |
Глутамин |
0,6±0,1 |
0,4±0,1* |
0,6±0,1 |
0,4±0,1 |
0,5±0,1* |
0,6±0,1* |
Аспарагин |
6,1±0,6 |
6,2±0,6 |
5,4±0,5* |
5,5±0,5 |
5,9±0,6 |
5,8±0,6 |
Примечание. Прочерки означают, что содержание аминокислоты в образце было ниже предела детекции (для аргинина и гистидина менее 0,50 %, для метионина менее 0,25 %). |
||||||
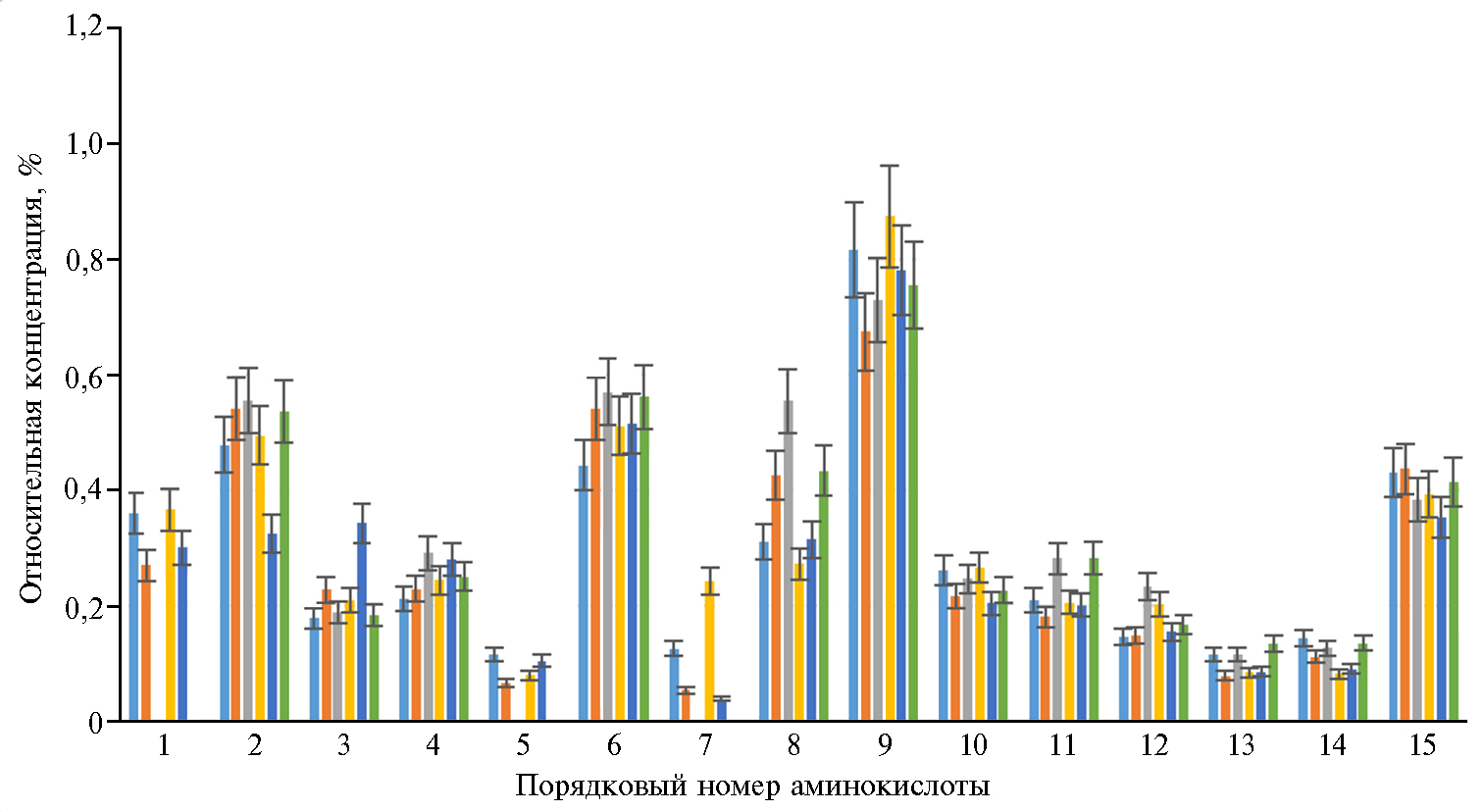
Массовая доля аминокислот (%) в молоке коров (Bos taurus taurus) черно-пестрой породы, инфицированных BIV (bovine immunodeficiency virus) или коинфицированных BLV (bovine leukemia virus) и BIV, после хранения в холодильнике при 4 °С: 1 — аргинин, 2 — лизин, 3 — тирозин, 4 — фенилаланин, 5 — гистидин, 6 — лейцин + изолейцин, 7 — метионин, 8 — валин, 9 — пролин, 10 — треонин, 11 — серин, 12 — аланин, 13 — глицин, 14 — глутамин, 15 — аспарагин; 




 — соответственно 1-е сут (BIV), 3-и сут (BIV), 6-е сут (BIV), 1-е сут (BLV-BIV), 3-и сут (BLV-BIV), 6-е сут (BLV-BIV) (M±σ). Наблюдаемые различия статистически значимы при р < 0,05.
— соответственно 1-е сут (BIV), 3-и сут (BIV), 6-е сут (BIV), 1-е сут (BLV-BIV), 3-и сут (BLV-BIV), 6-е сут (BLV-BIV) (M±σ). Наблюдаемые различия статистически значимы при р < 0,05.
ЛИТЕРАТУРА
- Овсянникова О.В., Ксёнз М.В. Обоснование возможности получения пищевых белковых продуктов из семян подсолнечника. Сфера услуг: инновации и качество, 2012, 7: 30-43.
- Teunissen-Beekman K.F., Dopheide J., Geleijnse J.M., Bakker S.J., Brink E.J., de Leeuw P.W., van Baak M.A. Effect of increased protein intake on renal acid load and renal hemodynamic responses. Physiological Reports, 2016, 4(5): e12687 (doi: 10.14814/phy2.12687).
- Wu G. Dietary protein intake and human health. Food & Function, 2016, 7: 1251-1265 (doi: 10.1039/c5fo01530h).
- McGregor R.A., Poppitt S.D. Milk protein for improved metabolic health: a review of the evidence. Nutrition & Metabolism, 2013, 10: 46 (doi: 10.1186/1743-7075-10-46).
- Saubade F., Hemery Y.M., Guyot J.P., Humblot C. Lactic acid fermentation as a tool for increasing the folate content of foods. Critical Reviews in Food Science and Nutrition, 2017, 57(18): 3894-3910 (doi: 10.1080/10408398.2016.1192986).
- Savilahti E. Probiotics in the treatment and prevention of allergies in children. Bioscience and Microflora, 2011, 30(4): 119-128 (doi: 10.12938/bifidus.30.119).
- Ricci-Cabello I., Herrera M.O., Artacho R. Possible role of milk-derived bioactive peptides in the treatment and prevention of metabolic syndrome. Nutrition Reviews, 2012, 70(4): 241-255 (doi: 10.1111/j.1753-4887.2011.00448.x).
- Heine W.E. Protein source and microflora. In: Probiotics, other nutritional factors, and intestinal microflora. 2nd Nestle Nutrition Workshop, Beijing, China, May 1997 /N.A. Hanson, R.H. Yolken (eds.). Philadelphia, Lippincott-Raven, 1999: 175-188.
- Caldow M.K., Digby M.R., Cameron-Smith D. Short communication: Bovine-derived proteins activate STAT3 in human skeletal muscle in vitro. Journal of Dairy Science, 2015, 98(5): 3016-3019 (doi: 10.3168/jds.2014-9035).
- Hagi T., Sasaki K., Aso H., Nomura M. Adhesive properties of predominant bacteria in raw cow's milk to bovine mammary gland epithelial cells. Folia Microbiologica, 2013, 58(6): 515-522 (doi: 10.1007/s12223-013-0240-z).
- Горбатова К.К., Гунькова П.И. Химия и физика молока и молочных продуктов. М., 2012.
- Wu G. Amino acids: metabolism, functions, and nutrition. Amino Acids, 2009, 37(1): 1-17 (doi: 10.1007/s00726-009-0269-0).
- Yang Y., Fan W., Mao Y., Yang Z., Lu G., Zhang R., Zhang H., Szeto C., Wang C. Bovine leukemia virus infection in cattle of China: Association with reduced milk production and increased somatic cell score. Journal of Dairy Science, 2016, 99(5): 3688-3697 (doi: 10.3168/jds.2015-10580).
- Bauermann F.V., Ridpath J.F., Dargatz D.A. Bovine leukemia virus seroprevalence among cattle presented for slaughter in the United States. Journal of Veterinary Diagnostic Investigation, 2017, 29(5): 704-706 (doi: 10.1177/1040638717702183).
- Schnell S.A., Ohtsuka H., Kakinuma S., Yoshikawa Y., Watanabe K., Orino K. Iron and ferritin levels in the serum and milk of bovine leukemia virus-infected dairy cows. Front. Vet. Sci., 2015, 2: 12 (doi: 10.3389/fvets.2015.00012).
- Красникова Е.С., Утанова Г.Х., Федосов Н.А., Щербаков А.А. Оценка качества молока, полученного от инфицированных ретровирусами коров и определение способов его переработки. Научное обозрение, 2015, 17: 10-15.
- Della Libera A.M., de Souza F.N., Batista C.F., Santos B.P., de Azevedo L.F., Sanchez E.M., Diniz S.A., Silva M.X., Haddad J.P., Blagitz M.G. Effects of bovine leukemia virus infection on milk neutrophil function and the milk lymphocyte profile. Veterinary Research, 2015, 46: 2 (doi: 10.1186/s13567-014-0125-4).
- Закрепина Е.Н. Лейкоз крупного рогатого скота и его влияние на количественные и качественные показатели молочной продуктивности коров. Автореф. канд. дис. Вологда-Молочное, 2001.
- Brujeni G.N., Poorbazargani T.T., Nadin-Davis S., Tolooie M., Barjesteh N. Bovine immunodeficiency virus and bovine leukemia virus and their mixed infection in Iranian Holstein cattle. J. Infect. Dev. Ctries, 2010, 4(9): 576-579 (doi: 10.3855/jidc.711).
- Bhatia S., Patil S.S., Sood R. Bovine immunodeficiency virus: a lentiviral infection. Indian Journal of Virology, 2013, 24(3): 332-341 (doi: 10.1007/s13337-013-0165-9).
- Баркова А.С., Колчина А.Ф., Барашкин М.И., Лоретц О.Г., Тарасенко М.Н. Аминокислотный состав молока коров черно-пестрой породы. Мат. VII Межд. науч. практ. конф. «Образованието и науката на ХХI век — 2012». София, 2012: 95-100.
- Надточий Л.А., Орлова О.Ю. Инновации в биотехнологии. Ч. 2. Пищевая комбинаторика. СПб, 2015: 8.
- Yu X., Long Y.C. Autophagy modulates amino acid signaling network in myotubes: differential effects on mTORC1 pathway and the integrated stress response. FASEB J., 2015, 29(2): 394-407 (doi: 10.1096/fj.14-252841).
- Wu G. Dietary requirements of synthesizable amino acids by animals: a paradigm shift in protein nutrition. Journal of Animal Science and Biotechnology, 2014, 5(1): 34 (doi: 10.1186/2049-1891-5-34).
- Wu G. Functional amino acids in growth, reproduction, and health. Advances in Nutrition, 2010, 1(1): 31-37 (doi: 10.3945/an.110.1008).
- Ianni F., Sardella R., Lisanti A., Gioiello A., Cenci Goga B.T., Lindner W., Natalini B. Achiral-chiral two-dimensional chromatography of free amino acids in milk: a promising tool for detecting different levels of mastitis in cows. J. Pharm. Biomed. Anal., 2015, 116: 40-46 (doi: 10.1016/j.jpba.2014.12.041).
- Bochniarz M., Kocki T., Dąbrowski R., Szczubiał M., Wawron W., Turski W.A. Tryptophan, kynurenine, kynurenic acid concentrations and indoleamine 2,3-dioxygenase activity in serum and milk of dairy cows with subclinical mastitis caused by coagulase-negative staphylococci. Reprod. Domest. Anim., 2018, 53(6): 1491-1497 (doi: 10.1111/rda.13299).