








УДК 636.082.13+575.17
ГЕНЕТИЧЕСКОЕ МАРКИРОВАНИЕ, СОХРАНЕНИЕ БИОРАЗНООБРАЗИЯ И ПРОБЛЕМЫ РАЗВЕДЕНИЯ ЖИВОТНЫХ
Н.С. МАРЗАНОВ1, Д.А. ДЕВРИШОВ2, С.Н. МАРЗАНОВА2, Е.А. КОМКОВА1, М.Ю. ОЗЕРОВ3, Ю. КАНТАНЕН3
Обсуждаются вопросы сохранения генофонда сельскохозяйственных животных в России и за рубежом. Показана важность разработки новых маркирующих систем для оценки степени биоразнообразия животных и выявления наследственных молекулярно-генетических болезней. Установлено влияние однонаправленности селекционного процесса и индекса общего качества (total merit index — TMI) на жизнеспособность животного и качество получаемой продукции.
Ключевые слова: эволюция, порода, генофонд, сохранение биоразнообразия, микросателлиты, аллель, генотип, мутация, острова цивилизации.
С биологической точки зрения основой полноценной жизнедеятельности у сельскохозяйственных животных служит внутривидовая дифференциация на породы, отродья, типы, линии и семейства. Человек осознанно создает условия, ведущие к определенному неравенству части особей вида внутри любого его сообщества. Подобная регуляция формообразовательного процесса приводит к тому, что многие наследственные модификации не проявляются, но, накапливаясь скрыто, образуют в стаде своего рода резерв наследственной изменчивости (или пластичности), который при изменениях во внешней среде может быть мобилизован очень быстро (1, 2). У высших позвоночных скорость эволюции постоянно варьирует и, несмотря на то, что размножение у них происходит медленно, временами достигает значений, которые совершенно необъяснимы с точки зрения теории Ч. Дарвина в ее первоначальном виде. Изменение темпов эволюции связано с определенными причинами.
Наиболее ярко это проявляется в группах узкоспециализированных форм — пород, где эволюционная пластичность действительно резко падает при отсутствии грамотного селекционного надзора. Теорией стабилизирующего отбора объясняется наследственное фиксирование в ряду поколений тех признаков, которые организм приобретает в индивидуальной жизни. Это ведет к ускорению эволюции у высших животных. Стабилизирующим отбором, конечно, не подменяется движущий эффект дарвиновского, они всегда действуют совместно, составляя, в сущности, стороны единого эволюционного процесса.
Согласно додарвиновским взглядам, отдельные организмы представляют собой несовершенные образования, а их вариабельность — лишь внешняя видимость при неизменности вида, к которому они принадлежат. С современных позиций эволюция рассматривается как изменение генетического состава популяций. Чрезвычайно важным достоинством совершенного Ч. Дарвином переворота была замена типологического мышления популяционным (3, 4), что способствовало пониманию механизмов эволюции вообще и естественного отбора в частности. При этом на популяционном уровне постоянно проявляется влияние основных уровней организации биологических объектов — молекулярно-генетического и организменного.
Таким образом, формирование пород сельскохозяйственных животных представляет собой результат сложного взаимодействия между смежными популяциями вида, человеком и внешней средой. Межпопуляционные различия всегда носят групповой характер и касаются частоты и географического распространения морфологических, физиологических и генетических признаков, в том числе вредных мутаций, что, в частности, очень важно для изучения наследственных заболеваний. Диагностика этих болезней стала необходима в результате разведения так называемых коммерческих высокоспециализированных пород. К ним, например, относятся крупная белая порода свиней, которую выращивают в 117 странах, зааненская порода коз — в 81 стране, порода овец суффольк — в 40 странах (5). Социально-экономические условия оказывают мощное воздействие на все факторы микроэволюции: частоту мутаций, периодические колебания численности индивидов, характер изоляции и обусловленные ею границы между популяциями и, наконец, на естественный отбор, роль которого непрерывно снижается в процессе формирования и использования популяций, из которых состоит порода. На смену географической изоляции, игравшей главную роль на ранних этапах истории популяции животных, приходит изоляция производственная, государственная, религиозная и др.
Безусловно, дифференциация форм внутри вида происходит благодаря изменчивости под влиянием факторов внешней среды. Модификации могут иметь разный характер (обратимые и необратимые, наследственные и ненаследственные) и неодинаковое значение в эволюционном процессе. Мы не отрицаем также наличия определенных направленных изменений и их роли, как и не отрицаем значения ненаследственных приспособительных изменений. Однако в естественных условиях они не могут рассматриваться в качестве основы для эволюции, поскольку определенность, направленность и относительная целесообразность этих изменений не представляются изначальными свойствами живых существ. Свободное накопление первично ненаправленных наследственных изменений (мутаций) в популяции приводит к разрушению существующей генетической организации вида. Такое всегда наблюдается при ослаблении действия естественного отбора или других форм контроля.
Наглядным примером последствий подобного ослабления служит отсутствие реализации генетического потенциала у породы. Как известно, при утверждении породы или породного типа на первое место выходят продуктивные признаки, но, к сожалению, из-за действия человеческого фактора многие отечественные породы часто не могут реализовать имеющийся потенциал продуктивности. Например, костромская порода в месте создания показывает высокие продуктивные качества, тогда как в других зонах разведения возможностей для этого нет. Аналогичная ситуация с большинством отечественных пород и в других регионах России. Современное животноводство зиждется на использовании небольшого числа жестко отобранных производителей, что зачастую ведет не только к увеличению продуктивности, но и к накоплению «генетического мусора» в виде наследственных болезней, гены которых выступают своеобразным «шлейфом» высокопродуктивных пород. Таким образом, внедрение селекционных схем, построенных на одном или нескольких признаках, неизбежно вызывает сужение генетической базы и повышение уровня инбридинга у высокоспециализированных пород.
Широкое использование описанной модели усугубляется тем, что происходит разведение родственных животных, соответственно имеет место однонаправленность селекции. Мировая популяция голштинской породы исчисляется миллионами особей, ее разводят в 128 странах. Известно, что эффективный размер популяции голштинских быков в США примерно 40 животных, аналогичная ситуация в России. Около 25 % генов молодых бычков в датской популяции голштинов происходит от двух производителей (6). Это драматичное уменьшение эффективного размера популяции элитных быков приводит к снижению результативности селекции и распространению летальных болезней, вызываемых рецессивными аллелями (bovine leukocyte adhesion deficiency, BLAD — дефицит лейкоцитарной адгезии; complex vertebral malformation, CVM — комплексная аномалия позвоночника; deficiency of uridine monophosphate synthase, DUMPS — дефицит уридинмонофосфатсинтетазы и др.) (7, 8). Аллель BLAD-синдрома (дефицит лейкоцитарной адгезии), будучи «шлейфом» высокой молочности, распространился по всему миру, в том числе попал в отечественные стада голштинизированного скота при приобретении племенных животных, замороженного семени или эмбрионов из Германии, Канады, США, Голландии до разработки методов диагностики носительства. В настоящее время он передается внутри или между популяциями уже от выращенного в России племенного материала.
| 1. Геногеография распространения мутации дефицита лейкоцитарной адгезии BLAD в популяциях черно-пестрого, голштинского, черно- и красно-пест-рого голштинизированного крупного рогатого скота в регионах Российской Федерации | ||||||||
Область, край |
Число животных |
Распределение |
Генотип BLAD |
Частота аллеля |
χ2 |
|||
NN |
Nn |
nn |
N |
n |
||||
Московская, ОАО Центральная стан-ция искусственного осеменения |
377 |
Н |
367 |
10 |
0 |
0,985 |
0,014 |
1,723 |
Ленинградская, ПП «Невское» (цит. по 9) |
127 |
Н |
121 |
6 |
0 |
0,976 |
0,024 |
0,161 |
Кировская, ОАО «Кировплем» |
149 |
Н |
148 |
1 |
0 |
0,997 |
0,003 |
0,036 |
Воронежская |
31 |
Н |
30 |
1 |
0 |
0,984 |
0,016 |
0,011 |
Вологодская |
2 |
Н |
1 |
1 |
0 |
0,750 |
0,250 |
0,222 |
Тамбовская |
98 |
Н |
98 |
0 |
0 |
1 |
0 |
0 |
Хабаровский |
234 |
Н |
233 |
1 |
0 |
0,998 |
0,002 |
0,020 |
Всего |
1018 |
Н |
998 |
20 |
0 |
0,990 |
0,010 |
0,141 |
П р и м е ч а н и е. Н и О — соответственно наблюдаемое и ожидаемое число генотипов; N и n — нормальный и мутантный аллель. |
||||||||
|
Схема распространения аллеля BLAD (bovine leukocyte adhesion deficiency) в генеалогических линиях черно-пестрого крупного рогатого скота в Российской Федерации. |
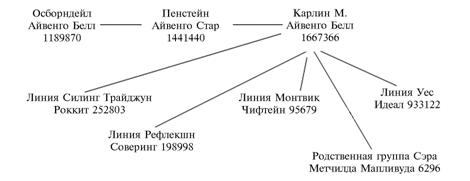
Частота аллеля BLAD (табл. 1) выше в Московской и Ленинградской областях. Он также выявляется в Вологодской, Воронежской, Кировской и Новосибирской областях, в Хабаровском крае у некоторых быков-производителей, коров и молодняка разного возраста. В СНГ аллель BLAD обнаружен в Украине и Беларуси, но отсутствует в Казахстане и Молдове. В популяциях черно- и красно-пестрого скота 998 животных из 1018 обследованных оказались гомозиготами NN (D128/D128) по нормальному аллелю D128 (неносители), 20, или 1,96 %, — гетерозиготами (Nn) и имели генотип D128/D128G (носители). По всем исследованным популяциям черно-пестрого, голштинского, черно- и красно-пестрого голштинизированного скота (n = 1018) частота нормального аллеля (N) составила 0,990, тогда как мутантного (n) — 0,010, или 1,0 % (см. табл. 1).
Мутантный аллель BLAD выявлен у всех известных генеалогических линий черно-пестрого скота (рис.). Использование сыновей и внуков американского голштинского быка-производителя Карлина М. Айвенго Белла 1667366 из группы, родственной быку Осборндейлу Айвенго Беллу 1189870, способствовало повышению генетического потенциала отечественных стад черно-пестрого скота. В то же время Карлин М. Айвенго Белл 1667366 стал основным распространителем BLAD в мире, в России, в странах СНГ и Балтии. В Хабаровском крае носителем аллеля BLAD оказалась телочка создаваемого дальневосточного типа черно-пестрой породы. Мы предполагаем, что ей аллель BLAD передала мать, отцом которой был Оливер 2170, принадлежащий ОАО «Центральная станция искусственного осеменения». Оливер 2170 получил этот аллель через мать от бабушки — дочери Карлина М. Айвенго Белла 1667366.
Среди носителей аллеля BLAD в Воронежской области впервые в этой области и в России было обнаружено животное красно-пестрой породы. В Тамбовской и Рязанской областях носителей BLAD не зарегистрировали. В популяциях выявленных нами носителей в основном представлены потомки Карлина М. Айвенго Белла 1667366. Тот же бык оказался основным источником распространения другой наследственной болезни CVM — комплексной аномалии позвоночника, которая ему передалась от отца. Следует отметить, что в настоящее время у крупного рогатого скота описаны 40 наследственных заболеваний, которые выявляются на уровне ДНК (10).
Улучшая молочную продуктивность и форму вымени за счет скрещиваний с голштинами, мы столкнулись не только с чрезвычайно острой проблемой множества генетических болезней, но и с сужением генетического биоразнообразия. Это, в частности, подтвердили результаты, полученные независимо с использованием двух маркирующих систем (микросателлитный анализ и тест на распространение групп крови) в условиях северных регионов России на черно-пестром генеалогическом корне крупного рогатого скота (11, 12). Как уже отмечалось, в природе изменения происходят медленно, так как различия между особями невелики и отбор осуществляется за счет «выбраковки» мало приспособленных к конкретным условиям или недостаточно плодовитых особей. При селекции, то есть искусственном преобразовании животных, место естественного отбора занимает активный выбор немногих наиболее подходящих форм и их последующее размножение. Один из приемов усиления различий между особями — комбинирование признаков и направление изменений в нужное русло, что, безусловно, способствует совершенствованию методов селекции. Еще больший эффект достигается при получении животных с определенными известными наследственными изменениями. В настоящее время такой подход из чисто научного превращается в практический. Один из примеров — создание свиней с улучшенным качеством мяса, имеющих гетерозиготный генотип (Nn) по рецессивному (n) аллелю RYR1. Однако в результате использования современных селекционных программ вытесняются местные, менее продуктивные породы, утрачивается их генетическое разнообразие, что в целом снижает вероятность адаптации селекционных схем к будущим потребностям животноводческой отрасли.
Суть известной скандинавской модели (Nordic Profile) в селекции молочных коров заключается в разведении только продуктивных, но в то же время здоровых животных. Селекция основывается на индексе общего качества (total merit index — TMI), который включает оценку по комплексу показателей — высокому надою молока, репродуктивным качествам, резистентности к маститу, другим функциональным признакам, что, на наш взгляд, правильнее, чем ограничиваться только величиной удоя. TMI основывается на уникальной базе данных, содержащей материалы по продуктивности, воспроизводству, лечению животного и т.д. Широкий генетический профиль элитных быков-производителей позволяет специалистам осуществлять целенаправленный подбор родительских пар, тем самым оптимизируя генотип потомка. Следует, однако, отметить, что в фермерских хозяйствах Дании, предоставляющих материалы в эту базу более 30 лет, большинство включаемых в программы по искусственному осеменению молодых быков все еще представлены сыновьями импортированных быков, которые отбирались главным образом по надою матерей (в 1999-2000 годах — приблизительно 80 % быков, использованных в голштинской популяции Дании), и сведения о них в базе TMI неполные. Тем не менее накоплено достаточно данных, подтверждающих экономическую эффективность описанной модели (13).
Индивидуальная идентификация животных, сбор информации о племенной ценности, учет продуктивности, особенностей репродукции и роста, использование новых статистических методов и компьютерных технологий позволили выявлять выдающихся животных с высокой степенью точности. С 2008 года специалистами из США предложена геномная оценка прогноза племенной ценности животных голштинской, джерсейской, бурой швицкой и других пород крупного рогатого скота (genomic predicted transmitting abilities). Она основывается на использовании пакета из 60 000 нуклеотидных генетических маркеров, из которых около 40 000 считаются информативными, поскольку оказывают влияние на продуктивные и другие признаки. Метод одновременно решает широкий спектр задач: установление достоверности происхождения потомства, степени родства и гетерозиготности, диагностика аномалий в геноме животного, выявление возможности улучшения продуктивных признаков и устойчивости к болезням. Этот опыт по широкому использованию ДНК-технологии в разведении молочного и мясного крупного рогатого скота, других видов сельскохозяйственных животных в настоящее время перенимают Голландия, Канада, Великобритания, ФРГ, Япония, Австралия и другие страны. Идентификация хозяйственно ценных особей, внедрение современных методов воспроизводства (искусственное осеменение, трансплантация эмбрионов, а также применение генной инженерии) и международная торговля племенным материалом привели к глобальному распространению нескольких так называемых коммерческих американских и западноевропейских пород. Они обладают высоким потенциалом продуктивности, но в то же время животные этих пород требовательны к качеству кормов, при их выращивании необходим высокий уровень организации труда, жесткий контроль зоогигиенических условий (температура, влажность, световой режим) и состояния окружающей среды, четкая работа ветеринарной службы (7).
В 1928 году понятие «геногеография» (географическое распространение характерных генетических маркеров и других генетических признаков) впервые ввел российский генетик А.С. Серебровский (1892-1948). Геногеографические исследования, как правило, включают картографический анализ распространения генетических маркеров (гаплогрупп) в популяциях крупных регионов и детальное изучение нескольких конкретных популяций. Проводятся экспедиционные исследования и генотипирование ДНК- и классических маркеров: однородительских (например, митохондриальной ДНК — мтДНК, Y-хромосомы), генетико-биохимических и других систем (14, 15).
| 2. Показатели генетической структуры пород крупного рогатого скота, разводимых на территории Российской Федерации | ||||||
Порода |
n |
Ho |
He |
Ae |
Ao |
Apr |
Суксунская |
40 |
0,6813 |
0,6909 |
3,63 |
6,00 |
1 |
Истобенская |
49 |
0,6895 |
0,6776 |
3,61 |
6,13 |
– |
Ярославская |
44 |
0,6977 |
0,6895 |
3,68 |
6,47 |
3 |
Холмогорская |
42 |
0,6392 |
0,6479 |
3,23 |
5,87 |
2 |
Печорский породный тип |
33 |
0,6438 |
0,6477 |
3,22 |
5,43 |
– |
Черно-пестрая |
30 |
0,6656 |
0,6716 |
3,41 |
5,80 |
1 |
Красная степная |
37 |
0,7504 |
0,7423 |
4,50 |
7,83 |
10 |
Бестужевская |
67 |
0,6560 |
0,6772 |
3,53 |
6,53 |
2 |
Калмыцкая |
28 |
0,6875 |
0,6511 |
3,36 |
6,13 |
2 |
Казахская белоголовая |
40 |
0,7053 |
0,6901 |
3,64 |
7,20 |
3 |
Якутская |
60 |
0,5832 |
0,5842 |
2,62 |
5,00 |
5 |
П р и м е ч а н и е. n — число обследованных животных; Ho — наблюдаемая гетерозиготность; He — ожидаемая гетерозиготность; Ae — среднеe эффективное число аллелей; Ao — среднеe наблюдаемое число аллелей; Apr — число прайвит- (или породоспецифических) аллелей. Прочерк означает отсутствие породоспецифических аллелей. |
||||||
При анализе аллелофонда 11 пород крупного рогатого скота, которых разводят в разных географических зонах России (16), по 30 микросателлитным локусам выявили 339 аллелей (табл. 2). Среднее число аллелей на один локус составило 11,3 при колебаниях от 5,0 (ILSTS005) до 21,0 (TGLA122). Впервые в 9 из 11 исследованных популяций крупного рогатого скота установлено наличие прайвит-аллелей (или породоспецифических аллелей). Исключением оказались истобенская порода и печорский породный тип, у которых они не обнаружены. Наибольшее число породоспецифических аллелей выявлено у красной степной (n = 10) и якутской (n = 5) пород крупного рогатого скота (см. табл. 2). Что касается овец, то у них прайвит-аллели обнаружили только у андийской породы из Республики Дагестан (n = 1) и эдильбаевской породы из Астраханской области (n = 3) (10). Наименьшую степень наблюдаемой гетерозиготности установили у якутской (Нo = 0,5832), наибольшую — у красной степной породы (Нo = 0,7504). У якутской породы среднее эффективное число аллелей оказалось наименьшим (Ae = 2,62), у красной степной — наибольшим (Ae = 4,50). У красной степной породы из Республики Дагестан выявлены аллели, которые встречаются у зебу: по локусу BM2113 — 130 и 142 п.н., по CSSM66 — 181 п.н., ETH10-207 — 209 и 211 п.н., ETH152 — 191 п.н., HEL1 — 101, 107 и 117 п.н., HEL13 — 182 и 186 п.н., TGLA122 — 144 п.н. Мы объясняем это тем, что в Республике Дагестан довольно широко применяется гибридизация красной степной породы с зебу, поскольку зебукоровы устойчивы к кровепаразитарным болезням.
При кластерном анализе коммерческих и местных пород крупного рогатого скота в Российской Федерации их чистота в среднем составляла 90,8 %. Вместе с тем по якутской и калмыцкой породам этот показатель равнялся 100 %, что свидетельствует о консолидированности животных из старинных популяций. Сравнение с данными литературы позволило сделать вывод, что суксунская порода близка к белорусской красной, а обе они, в свою очередь, — к датской красной, от которой произошли. Что касается черно-пестрой, холмогорской, печорской и истобенской пород, то они генетически близки к голштинской: в последние годы быки голштинской породы интенсивно используются в этих популяциях. Широкая колонизация местных пород европейской части севера России голштинами приводит к размыванию корней у первых и созданию единой популяции крупного рогатого скота черно-пестрого генеалогического корня. Признанная неудачной идея создания монопороды на самом деле неформально реализуется в худших традициях российского скотоводства. В итоге это приводит к естественному росту степени гомозиготности, потере породо-специфических признаков, созданию синтетических популяций, снижению оплодотворяемости, болезням репродуктивной сферы, конечностей, маститу и лейкозу. Как следствие, сокращаются сроки продуктивного использования животных, снижается качество продуктов питания и получаемого потомства. По сути, в молочном скотоводстве затраты на ветеринарные мероприятия становятся одной из основных статей расхода (17).
Что касается генетической характеристики азиатских пород крупного рогатого скота (якутской, калмыцкой и казахской белоголовой), то и здесь имеется своя специфика, которую необходимо учитывать. Так, у якутского скота низкая гетерозиготность (Нo = 0,5832) (см. табл. 2) обусловлена малочисленностью, географической изолированностью, дрейфом генов, эффектом основателя. Такой естественный рост гомозиготности проявляется в снижении оплодотворяемости, высокой яловости и необходимости многократного осеменения. Аналогичная ситуация может возникнуть у калмыцкого скота и казахской белоголовой породы, хотя степень гетерозиготности у этих животных выше (соответственно 0,6875 и 0,7053), чем у якутского скота (11).
Судя по публикациям, с 2005 года реализуются международные глобальные геногеографические проекты по исследованию изменчивости мтДНК и Y-хромосом у животных, населяющих Землю, с целью составления детального генетического атласа пород и изучения древних путей миграции видов (14, 15).
Бюджеты проектов различны. Так, международный проект «Генография» (The Genographic project, крупнейший геногеографический проект изучения человека) превышает 40 млн долларов США. Он финансируется Географическим обществом США, фирмой IBM и другими спонсорами. Благодаря подобным исследованиям стало, например, возможно выявление многочисленных прямых потомков Чингизхана (в основном на территории Монголии), аналогичные проекты осуществлены и в отношении потомков царя Соломона (они найдены в Израиле и Эфиопии).
В рамках проекта «Россия в процессе перемен», который спонсировался Финской академией наук (18), инициированы фундаментальные исследования животного мира на стыке молекулярной биологии, прикладной математики и высокопроизводительных вычислений. Роль этих исследований поддерживается по линии ФАО Международной программой по решению проблемы исчезающих пород, их оценке по продуктивным и биологическим маркерам с целью создания морфогенетического и метаболического паспорта. Данные по статусу риска пород сельскохозяйственных животных показали угрозу их биоразнообразию. По информации ФАО, в 2000-2006 годах ежемесячно в мире утрачивается одна порода, причем Россия находится в числе лидеров по потерям основных видов (крупный рогатый скот, свиньи, овцы, козы, лошади), хотя на ее долю приходится 17 % породного разнообразия (19).
Главная цель проектов по сохранению генофонда — минимизировать потери генетических вариаций в малочисленных породах и популяциях с помощью племенной и/или маркерной селекции в сочетании с современными зообиотехнологическими методами (эмбриотрансплантация, геномная дактилоскопия). Обычно подобные проекты основаны на учете селекционного потенциала животных и подборе родительских пар. Алгоритм такого подхода был реализован в программе, получившей название EVA (6). Он позволяет увеличить племенную ценность признаков в последующих поколениях с учетом степени инбридинга. Раскрытие племенной ценности — это возможность уравнять вклад родителей. Было показано, что ежегодная потеря генетического многообразия может быть сокращена на 40-50 % при правильном сборе племенной информации о предках. Снижению степени инбридинга способствовало накопление данных о генотипе родителей по маркирующим системам. В теории при неограниченном размере популяции возможно полное сохранение всех аллелей (20), на практике же редкие аллели со временем утрачиваются из-за ограниченности популяций или определенных генетических событий (эффект «бутылочного горлышка») (21-22).
В настоящее время в Мировом банке данных ФАО насчитывается 7616 пород (6536 локальных и 1080 транснациональных, или широко распространенных), причем 1491 относят к находящимся на стадии исчезновения. Возможно, эта цифра неточна, поскольку по 36 % пород какие-либо данные отсутствуют. К районам с высокой степенью риска относятся Европа и Кавказ (28 % пород млекопитающих и 49 % — птицы), а также Северная Америка (соответственно 20 и 79 %). Для этих же районов характерно наличие небольшого числа специализированных пород с высокой продуктивностью. Вместе с тем для Латинской Америки и Карибского бассейна по 68 % пород млекопитающих и 81 % — птицы какие-либо данные также отсутствуют, для Африки — соответственно по 59 и 60 %, что серьезно осложняет проведение эффективной политики сохранения пород. Необходимо активизировать поиск таких групп животных, усовершенствовать методы оценки размеров и оптимизации структуры популяций, выявления других факторов, влияющих на сохранение генофонда.
Потеря пород вызывает обоснованное беспокойство. В Восточной Европе, СНГ и России некоторые местные породы обладают уникальными особенностями, сложившимися в течение нескольких веков, — устойчивостью к болезням, адаптацией к экстремальным условиям среды, более высоким качеством продукции. Вместе с тем они менее продуктивны, чем западные коммерческие породы, инфраструктура, необходимая для повышения продуктивности, у них слабее, к тому же для существенного генетического улучшения местных пород требуется немало времени. Обычно для достижения быстрого генетического прогресса локальные породы скрещивают с импортными высокопродуктивными. Подобная близорукая селекционная политика может привести к тому, что некоторые местные породы и их вышеупомянутые особые качества исчезнут навсегда.
Усадебных, или поместных, животных в средние века называли ландрасами (land — усадьба, поместье; race — раса, порода). Первые породы были зарегистрированы в племенных книгах в конце XIX века, а в начале XX, когда был достигнут определенный генетический прогресс, началось систематическое накопление данных по продуктивности. Импорт животных для улучшения местного скота тогда уже был повсеместным, и многие ландрасы исчезли в начале XX века, однако несомненно, что масштабы негативных тенденций революционизированного разведения животных полностью проявились в течение второй половины XX века.
Существуют различные способы сохранения биоразнообразия животных. Метод ex situ включает получение, оценку и замораживание семени и эмбрионов, а также ветеринарный контроль. Однако наиболее важным для сохранения домашних видов остается метод in situ как основная составляющая управления генетическими ресурсами, требующего взаимодействия менеджеров, специалистов, ассоциаций, а также государственных структур. В 1998 году страны — члены ФАО приняли решение о мерах по подготовке первого Доклада о состоянии мировых генетических ресурсов животных. Были сформулированы следующие приоритетные задачи по использованию и сохранению домашних животных: определение ядра генетических ресурсов для каждого вида; установление приоритетов и действия по генетическому улучшению видов и пород животных; выявление наиболее подходящей среды для реализации потенциала видов и пород животных; мониторинг популяций, находящихся в зоне риска; помощь в разработке систем раннего оповещения для пород, находящихся в опасности. В настоящее время в ряде стран при министерствах сельского хозяйства созданы комитеты по управлению генетическими ресурсами сельскохозяйственных животных.
В скандинавских странах такие комитеты функционируют более 25 лет «… с целью охраны биологического разнообразия, культурного наследия и культурных пейзажей». Например, в Дании эта политика проводится в рамках «Стратегии по управлению генетическими ресурсами сельскохозяйственных животных», обеспечивающей сохранение старых пород. Программа включает разработку селекционных планов для старых пород Дании; расширение существующего генетического банка; накопление информации и улучшение взаимодействия между различными структурами по разведению в хозяйствах редких пород; исследования и разработку планов по эффективному управлению генетическими ресурсами животных. По всем перечисленным направлениям активность за последние годы значительно возросла. Комитет основал секретариат, который тесно взаимодействует с селекционерами старых пород (как из частных, так и из государственных структур), что привело к консолидации и увеличению численности старых датских пород скота, а также уделяет внимание коммерческим породам, взаимодействуя с датской Ассоциацией селекционеров крупного рогатого скота по вопросам систематического сбора и долговременного хранения семени от современных пород молочного скота.
В начале 1990-х годов был начат совместный проект по исследованию генетического разнообразия старых и редких пород крупного рогатого скота в Дании, Финляндии, Исландии, Норвегии и Швеции (финансирование осуществлялось Скандинавским генным банком животных). Целью проекта была оценка сходства и различия среди 15 местных и двух импортных пород крупного рогатого скота, а также сравнение местных и редких популяций с тремя коммерческими породами. Для характеристики использовали разнообразные генетические маркеры (группы крови, полиморфные белки и микросателлиты). Генетический анализ выявил различия между местными и старыми породами и позволил разделить их на четыре группы: северную местного скота, включающую исландский скот; южную локальную, включающую красный датский скот 1970 и датских шортгорнов; группу, состоящую из айрширского и фризского скота (в том числе датский черно-пестрый скот 1965 и ютландская порода); импортированную джерсейскую породу. Все исследованные породы генетически отличались от известных популяций норвежского крупного рогатого скота, одновременно демонстрируя очевидное смешение. Анализ микросателлитов показал, что различия между местными и коммерческими породами по степени наблюдаемой гетерозиготности и числу аллелей в локусе не были статистически достоверными. Вместе с тем отмеченные генетические различия свидетельствовали, что локальные популяции могут выступать в качестве резервата биоразнообразия (14, 16, 23, 24).
Исходя из фундаментальной идеи французского историка XX века Фернана Броделя о существовании «островов цивилизаций, окруженных сушей» (25, 26), животноводство в России можно рассматривать как крупную социально-биологическую структуру, состоящую из разных историко-культурных самобытностей. В научной литературе подобная попытка выделения региона в качестве самостоятельного историко-культурного острова, который существенно отличается от других, еще не предпринималась. Что формирует подобные острова цивилизаций? История, проявленное неповторимое своеобразие, особая региональная самобытность. В отношении не только животноводства, садоводства, но и в целом сельскохозяйственной отрасли таких островов на территории Российской Федерации несколько (Северный Кавказ, Верхне-Волжский район, Забайкалье, зона тундры, Республика Тыва, Республика Саха—Якутия, Алтай), что целесообразно учитывать при создании пород, перспективных в производственном отношении в будущем. Это важный этап в продолжении породообразовательного процесса, для которого необходимы селекционные программы, адаптированные к местным условиям и потребностям. Наиболее рациональный и эффективный способ сохранения генетических ресурсов — использование местных адаптированных пород как неотъемлемой функциональной части системы по производству животноводческой продукции. Безусловно, для этого необходимо выявлять и развивать их экономически значимые и уникальные признаки (27, 28).
Сохранить все многообразие существующих сельскохозяйственных животных, конечно, невозможно, однако важно, чтобы породы или местные региональные группы пород, линии и типы, обладающие особыми качествами, не были утрачены. Для этого необходимо проводить их идентификацию, описание, оценку. Генетические маркеры обеспечивают описание пород по основным характеристикам, однако сбор данных по продуктивности (удой, показатели роста, функциональные признаки) обязательно должен продолжаться как неотъемлемая часть оценки продуктивного потенциала животного и породы.
Научное сообщество может и должно влиять на племенные предприятия через принятие соответствующих селекционных программ. Необходимо также, чтобы государство стимулировало ученых и животноводов, эффективно использующих старые породы. Пока что в нашей стране управлению генетическими ресурсами уделяется меньше внимания, чем требуется, однако изменения условий среды делают все более заметной необходимость переосмысления политики ведения животноводства, размещения пород, даже размеров животного.
Периодическому обобщению состояния животноводства посвящены ежегодные совещания селекционеров племенных заводов, которые проводятся Всероссийским НИИ племенного дела совместно с Министерством сельского хозяйства РФ, однако речь идет только о племенных животных. Привлечение внимания к проблеме сохранения биоразнообразия, выполнения соответствующих генетических исследований повысило бы статус этого мероприятия. По нашему мнению, такой подход позволит оценить состояние генетических ресурсов по видам и породам животных, выделить ключевые проблемы и определить возможности для их решения, консолидировать усилия по оптимальному использованию генетических ресурсов.
Итак, в настоящее время многие селекционные программы признаются непригодными для долговременного использования: наблюдается стремительное увеличение инбридинга, накопление генетических болезней, что ощутимо снижает эффект селекции и повышает прямые потери продуктивности. Следовательно, управлению генетическими ресурсами будет придаваться все большее значение. Сейчас в России выполняются генетические исследования состояния пород овец, коз, крупного рогатого скота, проводится оценка аллелофонда, генетической структуры и динамики популяций, разрабатываются системы маркирования генотипов. Конечной целью таких исследований должно быть сохранение и эффективное использование в селекции того огромного генетического (в том числе адаптивного) потенциала, который все еще сохраняется у отечественных сельскохозяйственных видов животных, у локальных пород. Важно, чтобы селекционеры, хозяйства, ассоциации и союзы, общество в целом осознавали значение этой проблемы для разработки и осуществления селекционных программ и обеспечения продовольственной безопасности. Дальновидность стратегии бережного использования генетических ресурсов отечественных видов и пород домашних, а также популяций диких животных должна обосновываться в научных публикациях и широко дискутироваться в средствах массовой информации.
Л И Т Е Р А Т У Р А
1. Ш м а л ь г а у з е н И.И. Вопросы дарвинизма. Неопубликованные работы. М., 1990.
2. М а р з а н о в Н.С. Генетические ресурсы животных — это национальная ответственность. Изв. ТСХА, 2007, 5: 135-141.
3. М а й р Э. Популяции, виды и эволюция. М., 1974.
4. M a y r E. Speciation and macroevolution. Evolution, 1982, 36(6): 1119-1133.
5. http://dad.fao.org
6. V i g h - L a r s e n F., L i b o r i u s s e n T., B e r g P., H o l m L.-E. Sustainable use and conservation of farm animal genetic resources. Dias Report, 2002, 38: 219-229.
7. Т у р б и н а И.С. Характеристика быков-производителей по различным генетическим маркерам. Канд. дис. М., 2006.
8. К и й к о Е.И. Геногеография вариантов каппа-казеина и дефицита лейкоцитарной адгезии у различных пород крупного рогатого скота. Канд. дис. Дубровицы, 2009.
9. Я к о в л е в А., Т е р л е ц к и й В., М и т р о ф а н о в а О., Д е м е н т ь е в а Н. Определение носителей генетических дефектов среди быков-производителей. Молочное и мясное скотоводство, 2004, 7: 31-32.
10. S c h ü t z E., S c h a r f e n s t e i n M., B r e n i g B. Implication of complex vertebral malformation and bovine leukocyte adhesion deficiency DNA-based testing on disease frequency in the holstein population. J. Dairy Sci., 2008, 91: 4854-4859.
11. М а р з а н о в Н.С. Изучение полиморфизма ядерной и митохондриальной ДНК для оценки биоразнообразия и хозяйственно-биологических качеств сельскохозяйственных животных (отчет по теме). Дубровицы, 2008.
12. П о п о в Н.А. Генетические и фенотипические аспекты оценки гетерозиготности при селекции в популяциях крупного рогатого скота молочных пород (отчет по теме). Дубровицы, 2009.
13. S o r e n s e n M.K. Stochastic stimulation of breeding schemes for dairy cattle. Ph. D. thesis. Royal Veterinary and Agricultural University, Copenhagen, 1999.
14. T a p i o M., M a r z a n o v N., O z e r o v M., C i n k u l o v M., G o n z a r e n k o G., K i s e l y o v a T., M u r a w s k i M., V i i n a l a s s H., K a n t a n e n J. Sheep mitochondrial DNA variation in European, Caucasian and Central Asian areas. Mol. Biol. Evol., 2006, 23(9): 1776-1783.
15. M e a d o w s J.R.S., H a n o t t e O., D r o g e m ü l l e r C., C a l v o J., G o d f r e y R., C o l t m a n D., M a d d o x J.F., M a r z a n o v N., K a n t a n e n J., K i j a s J. Globally dispersed Y chromosomal haplotypes in wild and domestic sheep. Animal Genetics, 2006, 37(5): 444-453.
16. M e n g - H u a L i, T a p i o I., V i l k k i J., I v a n o v a Z., K i s e l y o v a T., M a r-
z a n o v N., СCi n k u l o v M., S t o j a n o v i c S., A m m o s o v I., P o p o v R., K a n t a n e n J. The genetic structure of cattle populations (Bos taurus) in northern Eurasia and the neighboring Near Eastern regions: implications for breeding strategies and conservation. Mol. Ecol., 2007, 16(18): 3839-3853.
17. С а м о р у к о в Ю., К а л я з и н а Т., М а р з а н о в Н. О породах в молочном скотоводстве. Молочное и мясное скотоводство, 2009, 6: 3-5.
18. A s k o M.-T. Genetic resources of Russian farm animals — the state of endangerment and ethno-ecological, technical and social opportunities for conservation. In: Russian in Flux — research program 1.1.2004-31.12.2007. Jokioinen, Finland, 2004: 11.
19. С т о л п о в с к и й Ю.А. Консервация генетических ресурсов сельскохозяйственных животных: проблемы и принципы их решения. М., 1997.
20. W a n g J., H i l l W.G. Marker-assisted selection to increase effective population size by reducing Mendelian segregation variance. Genetics, 2000, 154: 475-489.
21. О з е р о в М.Ю. Характеристика аллелофонда у различных пород овец по микросателлитам. Канд. дис. М., 2004.
22. О з е р о в М.Ю., Т а п и о М., М а р з а н о в Н.С., Н а с и б о в Н.Г., Ш а й д у л-
л и н И.Н., К а н т а н е н Ю. Микросателлитный анализ эволюционно-генетических связей у различных пород овец. Докл. РАСХН, 2006, 2: 30-33.
23. K a n t a n e n J., O l s a k e r I., H o l m L.-E., L i e n S., V i l k k i J., B r u s g a-
a r d K., E y t h o r s d o t t i r E., D a n e l l B., A d a l s t e i n s s o n S. Genetic diversity and population structure of 20 North European cattle breeds. Heredity, 2000, 91: 446-457.
24. С т о л п о в с к и й Ю.А. Ключевой вопрос в сохранении культурного биоразнообразия животных — сохранение породного многообразия. Изв. ТСХА, 2007, 5: 125-134.
25. Б р о д е л ь Ф. Средиземное море и средиземноморский мир в эпоху Филиппа II. Ч. 1. Роль среды. М., 2002: 208-210.
26. А г р б а Б.С., Х о т к о С.Х. «Островная» цивилизация Черкесии. Майкоп, 2004.
27. М а р з а н о в Н.С. Использование полиморфизма биологических маркеров для оценки генетического разнообразия и хозяйственно-полезных признаков у крупного рогатого скота и коз (отчет по теме). Дубровицы, 2010.
28. М а р з а н о в Н.С., С а м о р у к о в Ю.В., Е с к и н Г.В., Н а с и б о в Н.Г., М а р-
з а н о в а Л.К., К а н а т б а е в С.Г., Б у к а р о в Н.Г. Сохранение биоразнообразия. Генетические маркеры и селекция животных. С.-х. биол., 2006, 4: 3-19.
GENETIC LABELING, CONSERVATION OF BIODIVERSITY AND THE PROBLEMS IN ANIMALS BREEDING
N.S. Marzanov1, D.A. Devrishov2, S.N. Marzanova2, E.A. Komkova1, M.Yu. Ozerov3, Yu. Kantanen3
The problems in conservation of gene pool of agricultural animals in Russia and abroad are discussed. The authors showed the importance of development of new labeling systems for estimation of animals’ biodiversity and revelation of inherited molecular-genetic diseases. The influence of unidirectionality of breeding process and total merit index (TMI) on the viability of animals and the quality of obtained production was established.
Keywords: evolution, breed, gene pool, conservation of biodiversity, microsatellite, allele, genotype, mutation, civilization islands.
1ГНУ Всероссийский НИИ животноводства |
Поступила в редакцию |




