








УДК 636.594:575.113:577.175.322:57.086.864
БИОЛОГИЧЕСКИЕ И ПРОДУКТИВНЫЕ КАЧЕСТВА ПЕРЕПЕЛОВ, ТРАНСГЕННЫХ ПО ГЕНУ БЫЧЬЕГО СОМАТОТРОПИНА
Л.Г. КОРШУНОВА
Методом микроинъекции экзогенной ДНК — генной конструкции pMTbGH (2хatt) в зародышевые диски яйцеклеток получали перепелов, трансгенных по гену бычьего соматотропина, и оценивали их продуктивные качества в ряду поколений (18-33-е). Живая масса, число и средняя масса яйца у трансгенной птицы превышали аналогичные показатели у интактных перепелов. Жирнокислотный состав липидов яиц у трансгенных перепелов на зависел от этой генетической модификации.
Ключевые слова: перепел, яйцо, трансген, ген бычьего соматотропина, жирные кислоты.
Современная селекция в птицеводстве базируется на отборе лучшего поголовья из высокопродуктивных семей и семейств и требует длительного времени. Одним из нетрадиционных подходов к генетическому улучшению птицы может стать трансгенез. Несмотря на методические трудности, работы по созданию трансгенной птицы активно ведутся во многих странах, поскольку перспективны с точки зрения экономической выгоды (1-3).
Интеграция чужеродных генов в геном животного или растительного организма часто носит вероятностный характер. На сегодняшний день не всегда возможно интегрировать генную конструкцию в определенную точку генома, прогнозировать функционирование генов, расположенных рядом с ней, и, соответственно, предвидеть реакцию генома в целом на введение чужеродного материала. Трансгенез во многом напоминает мутацию, в значительной степени направленную, но сохраняющую множество неопределенностей. С одной стороны, это расширяет возможности получения животных с измененными признаками, многие из которых могут оказаться интересными для селекционера, с другой — усложняет работу, особенно когда в геном интегрируются генные конструкции, связанные с экспрессией биологически активных веществ регуляторного характера и оказывающие сильное воздействие на многие процессы обмена веществ.
Целью настоящей работы было изучение влияния интегрированной генной конструкции на хозяйственно ценные признаки у потомков генетически модифицированных перепелов.
Методика. Исследования проводили в 2001-2010 годах на перепелах эстонской породы в условиях вивария ГУП «Загорское ЭПХ ВНИТИП РАСХН» (Московская обл.) (4). Птицу содержали в индивидуальных клетках при свободном доступе к воде и корму.
Трансгенную птицу получали методом микроинъекции экзогенной ДНК в зародышевые диски яйцеклеток (5, 6). Инъецированная генная конструкция pMTbGH (2 x att) была предоставлена Институтом молекулярной биологии РАН (г. Москва) (7).
По данным оценки в предыдущих поколениях (8, 9) отобрали 20 пар из семей с высокими показателями яичной продуктивности, потомков которых использовали в настоящей работе. Опытную группу составляли потомки (18-33-е поколения) трансгенных особей, контрольную — эстонские перепела 15 генераций, не подвергшиеся генетической модификации. Отвод потомства в опытной и контрольной группах проводили естественным спариванием определенных особей. Каждая особь из опытной группы имела шифр в компьютерной базе данных, позволяющий проследить ее родословную, начиная от первичных трансгенных особей. Родословную для перепелов контрольной группы не отслеживали.
Для получения очередного поколения птицы яйца инкубировали в лабораторных условиях. Выведенных перепелов из опытной и контрольной групп взвешивали индивидуально в 1-суточном, 1-, 7- и 20-недельном возрасте (с разделением по полу). В течение 10 мес продуктивного периода проводили индивидуальный учет яйценоскости. Также в опытной и контрольной группах в течение 10 мес продуктивного периода оценивали массу каждого снесенного яйца (всего было взвешено по 135-150 тыс. яиц от каждой группы). Птицу и яйца взвешивали с точностью до 0,01 г. Первичные данные заносили в журналы учета и регистрации, а также в компьютерные базы для последующей сортировки и обработки.
Яйца для морфологического и биохимического анализа отбирали в течение 5 смежных суток в начале продуктивного периода, а также у птицы в возрасте 3, 6 и 10 мес. Проанализировали по 80 яиц из опытной и контрольной групп перепелов 30-го поколения. Для учета массы яйцо, скорлупу и желток взвешивали с точностью до 20 мг; количество белка вычисляли по разности между массой яйца и скорлупы с желтком. Содержание жирных кислот в липидах желтка определяли методом газожидкостной хроматографии (Fractovap-2450, «Carbo Erba», Италия). Использовали производные жирных кислот — метиловые эфиры, что обеспечило высокую эффективность разделения при более низких температурах, а также меньшее время анализа (10).
Статистическую обработку данных проводили в программе Microsoft Excel с определением t-критерия Стьюдента.
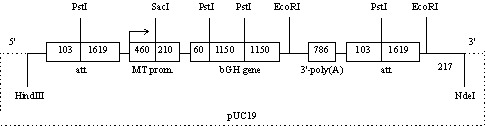
Результаты. Основной элемент конструкции pMTbGH (2 x att) (рис. 1) — содержащий интроны природный ген бычьего соматотропина (bGH gene), находящийся под контролем металлотионинового промотора мыши (MT prom.). Последний активируется ионами цинка и других тяжелых металлов, содержащихся в организме птицы. Активированный промотор заставляет экспрессироваться ген бычьего гормона роста. У трансгенных по этой конструкции перепелов происходит синтез указанного гормона, что продемонстрировано нами ранее (5, 11).
|
Рис. 1. Физическая карта плазмиды pMTbGH (2 x att) (указаны сайты рестрикции эндонуклеазами), использованной в качестве генной конструкции при получении генетически модифицированных перепелов (описание см. в разделе «Результаты»). |
У трансгенных особей в 15 исследованных поколениях сохранялось превосходство над контрольными по живой массе (рис. 2), причем у самок различия оказались более существенными. В 1-суточном и 1-недельном возрасте птица в опыте была крупнее контрольной соответственно на 21 и 13 %. Средняя масса 1-суточных перепелят в опытной группе составляла 9,1, в контрольной — 7,4 г. В 7- и 20-недельном возрасте достоверные различия наблюдались только у самок — соответственно 8 и 6 % (p < 0,001).
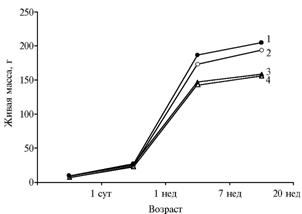 |
Рис. 2. Живая масса интактных перепелов эстонской породы (2, 4) и трансгенных по гену бычьего соматотропина (1, 3): 1, 2 — самки; 3, 4 — самцы (в среднем по 15 поколениям) (ГУП «Загорское ЭПХ ВНИТИП РАСХН», Московская обл., 2001-2010 годы). |
Масса 1-суточного молодняка птицы в значительной степени обусловлена массой инкубационных яиц. В наших опытах с увеличением массы яиц пропорционально увеличивалась масса трансгенных 1-суточных перепелят (r = +0,810). При этом все перепелята проявляли под-вижность, активно реагировали на звук, имели мягкий подобранный живот, чистую клоаку. Пух был мягким, ровным, блестящим, правильно пигментированным (коричневого цвета с двумя светлыми полосками на спинке), ноги и клюв — крепкими, глаза блестящими. Взрослые особи внешне не отличались от контрольных.
Живая масса трансгенных перепелов варьировала от поколения к поколению, однако линия тренда для этого показателя в 20-недельном возрасте свидетельствовала об отсутствии тенденции к снижению (наклон незначительный, R2 = 0,019) (рис. 3). Изменение средней живой массы у некоторых выводков, скорее всего, было связано с условиями содержания птицы, а не с особенностями их генотипа.
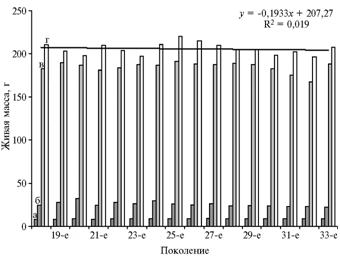 |
Рис. 3. Живая масса самок перепелов, трансгенных по гену бычьего соматотропина, в разных поколениях в зависимости от возраста: а — 1 сут, б — 1 нед, в — 7 нед, г — 20 нед (ГУП «Загорское ЭПХ ВНИТИП РАСХН», Московская обл., 2001-2010 годы). График соответствует линии тренда для живой массы в возрасте 20 нед. |
У трансгенных перепелов масса снесенных яиц оказалась больше, чем в контроле (рис. 4). Эта тенденция сохранялась во всех исследованных поколениях. Кроме того, в опытной группе число яиц, снесенных самками, было больше, чем в контроле. Масса яйца — один из основных показателей яичной продуктивности. В первую очередь он определяется видовой принадлежностью птицы и зависит от размеров са-мок. Обычная масса перепелиного яйца — 10-12 г. Внутри вида она различается по породам, но даже в пределах одной породы колебания достигают 20 % и более. Подобная вариабельность признака дает большие возможности для селекции. В наших опытах на пике продуктивности средняя масса яйца у потомков трансгенных перепелов по 30-му поколению составляла 13,0 г. Разброс показателя по отдельным особям был от 11,3 до 15,0 г (32,7 %).
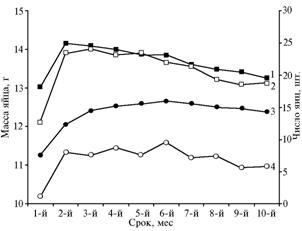 |
Рис. 4. Число (1, 2) и масса (3, 4) яиц, снесенных самками перепелов эстонской породы в период продуктивности: 1, 3 — трансгенная, 2, 4 — интактная птица (в среднем по 15 поколениям) (ГУП «Загорское ЭПХ ВНИТИП РАСХН», Московская обл., 2001-2010 годы). |
В течение опыта мы не всегда имели возможность поддерживать оп-тимальные условия содержания и необходимое качество кормов, что в большей степени влияло на продуктивность лучших не-сушек в опытной группе, чем в контрольной.
В опытной группе увеличение массы яйца до 15,0 г было обусловлено пропорциональным возрастанием массы его составных частей — скорлупы, белка и желтка. При этом доля желтка составляла 29,0-33,0 %, белка — 58-61 %, скорлупы — 9,0-10,0 %. Скорлупа яиц и в опытной, и в контрольной группе, как правило, имела характерную пеструю окраску с коричневыми пятнами.
Известно, что содержание основных компонентов птичьего яйца (протеинов, липидов, углеводов, воды, кальция, фосфора, железа, натрия, калия, хлора, магния, меди, серы) мало зависит от кормления и других внешних факторов. Однако другие биохимические показатели (количество витаминов, фтора, марганца, йода, олеиновой, линолевой, линоленовой кислот) в определенной степени вариабельны.
В яйцах у контрольных перепелов на долю насыщенных жирных кислот приходилось 41,35, мононенасыщенных — 41,08, полиненасыщенных — 17,59 %. Сумма насыщенных и полиненасыщенных жирных кислот в липидах желтка у птицы в опытной группе была соответственно на 1,77 и 10,35 % больше, сумма мононенасыщенных жирных кислот — на 6,26 % меньше, чем в контрольной. Выявленные различия обусловлены в основном увеличением содержания стеариновой (на 12,3 %) и линолевой (на 10,70 %) и уменьшением количества олеиновой (на 6,40 %) кислоты в яйцах у перепелов из опытной группы. При этом массовая доля олеиновой кислоты изменялась от 31,40 до 37,00 %, линолевой — от 19,90 до 17,30 %. Содержание жирных кислот в яйце не зависело от возраста несушек. В опытной группе количество стеариновой кислоты в яйцах у несушек 3- и 6-месячного возраста было соответственно на 13,61 и 20,75 %, линолевой кислоты — на 13,21 и 9,83 % выше контроля. Доля олеиновой кислоты в яйцах у 6-месячных несушек из опытной группы оказалась на 12,14 % ниже, чем в контроле. При изменении массы яйца и в опытных группах, и у контрольных перепелов наблюдались колебания в содержании отдельных жирных кислот в липидах желтка. Главным образом это относилось к жирным кислотам с 18 атомами углерода: стеариновой, олеиновой, линолевой и линоленовой. Выявленные модификации жирнокислотного состава липидов желтка — экспериментальный факт, однако чем они обусловлены и, в частности, связаны ли с трансгенностью, определить затруднительно, поскольку количество этих кислот в птичьем яйце может зависеть от многих факторов — питания, содержания, состояния здоровья и др. Полученные нами ранее результаты биохимического анализа перепелиных яиц по содержанию воды, сухих веществ, фосфора, витаминов А, Е, В, аминокислотному составу протеина (12) показали отсутствие закономерных различий между яйцами от интактных перепелов и потомков трансгенных особей (отмечались только незначительные изменения некоторых показателей).
Таким образом, живая масса, число и средняя масса яиц у перепелов, трансгенных по гену бычьего соматотропина, превышали аналогичные показатели у птицы, которая не была генетически модифицирована. Биохимический и морфологический анализ яиц трансгенных перепелов не выявил закономерных отличий их состава от такового у нетрансгенной птицы.
Л И Т Е Р А Т У Р А
1. K w o n M.S., K o o B.C., C h o i B.R., P a r k Y.Y., L e e Y.M., S u h H.S., P a r k Y.S., L e e H.T., K i m J.H., R o h J.Y., K i m N.H., K i m T. Generation of transgenic chickens that produce bioactive human granulocyte-colony stimulating factor. Mol. Reprod. Dev., 2008, 75(7): 1120-1126.
2. L e e S.H., G u p t a M.K., H a n D.W., H a n S.Y., U h m S.J., K i m T., L e e H.T. Development of transgenic chickens expressing human parathormone under the control of a ubiquitous promoter by using a retrovirus vector system. Poult. Sci., 2007, 86(10): 2221-2227.
3. V o l k o v a N.A., Z i n o v 'e v a N.A., V o l k o v a L.V., E r n s t L.K. Retroviral-mediated gene transfer as an effective tool for the in vitro genetic transformation of chicken embryonic cells and production of transgenic chickens. Genetika, 2006, 42(1): 84-88.
4. К о ч е т о в а З.И., Б е л я к о в а Л.С., Ф и л о н е н к о В.И.,
Ч и н ц о в а А.И. Разведение и содержание перепелов /Под ред. Т.А. Столяра. Сергиев Посад, 2006.
5. К а р а п е т я н Р.В. Фенотипические проявления трансгенности перепелов, развившихся из яйцеклеток, инъецированных чужеродной ДНК. С.-х. биол., 1997, 4: 89-96.
6. Ф и с и н и н В.И., Э р н с т Л.К., К а р а п е т я н Р.В.,
М а т в е е н к о Н.П., З а
з ы к и н а Т.В., З и а д и н о в а О.Ф.,
Ж а д а н о в А.Б. Способ повышения яичной продуктивности птицы. Всероссийский научно-исследовательский и технологический институт птицеводства. C1 6 A 01 K 67/02. RU Патент 2061366. М., 1996.
7. Ж а д а н о в А.Б. Получение векторов для экспрессии чужеродных генов в трансгенных животных. Автореф. канд. дис. М., 1991.
8. К а р а п е т я н Р.В., К о р ш у н о в а Л.Г., К о р ш у н о в К.Р.,
А м д и й Э.М., Э р н с т Л.К. Особенности фенотипа популяции потомков перепелов, трансгенных по гену бычьего соматотропина. Тез. докл. II Межд. науч. конф. «Биотехнология в растениеводстве, животноводстве и ветеринарии». М., 2000: 140-141.
9. К о р ш у н о в а Л.Г., К а р а п е т я н Р.В., К о р ш у н о в К.Р.,
С е р и к о в а В.А., З и а д и н о в а О.Ф. Сравнительная оценка яичной продуктивности потомков трансгенных и серых перепелов эстонской породы. Сб. науч. тр. ВНИТИП (Сергиев Посад), 2001, 76: 78-85.
10. А р х и п о в А.В. Изучение липидов и липидного обмена у птиц с применением тонкослойной и газожидкостной хроматографии: Метод. реком. М., 1973.
11. К о р ш у н о в а Л.Г., К а р а п е т я н Р.В. Использование полимеразной цепной реакции для идентификации интегрированного в геном перепела гена гормона роста быка. Мат. межд. симп. «Молекулярная генетика и биотехнология в оценке и изменении геномов с.-х. животных». СПб—Пушкин, 1994: 18-19.
12. К о р ш у н о в а Л.Г. Качество яиц трансгенных перепелов. Птицеводство, 2009, 4: 35-36.
BIOLOGICAL AND PRODUCTIVE PROPERTIES OF QUAILS TRANSGENIC ON BOVINE SOMATOTROPIN GENE
L.G. Korshunova
The practical useful determinants in quails of the Estonian breed, transgenic on bovine somatotropin gene, were studied. The live weight, number and average mass of eggs in transgenic birds were higher than similar parameters in native quails. The fatty-acid composition of egg lipids in transgenic quails do not depend on such genetic modification.
Keywords: quail, egg, transgen, bovine growth hormone gene, fat acids.
ГНУ Всероссийский научно-исследовательский |
Поступила в редакцию |




