








УДК 591.05:577.161.3
ОБ УЧАСТИИ α-ТОКОФЕРИЛХИНОНА В ПРОЦЕССАХ СБРАЖИВАНИЯ ГЛЮКОЗЫ ДРОЖЖАМИ Saccharomyces cerevisiae
В.И. ДУДИН
Изучали влияние α-токоферилхинона на процессы брожения у сухих свежеприго-товленных и подвергшихся продолжительному хранению пекарских дрожжей Saccharomyces cerevisiae в неполной среде с D-глюкозой или пируватом натрия. Установили, что α-токоферилхинон активно воздействует на процессы брожения на второй стадии гликолиза. Утилизация экзогенного пирувата уменьшалась в опытах с сухими свежими и увеличивалась в опытах с подвергшимися хранению пекарскими дрожжами.
Ключевые слова: сухие пекарские дрожжи Saccharomyces cerevisiae, глюкоза, уксусный альдегид, α-токоферилхинон, 3-дезокси-α-токоферилхинон, пировиноградная кислота, брожение.
α-Токоферилхинон — продукт свободнорадикального окисления витамина Е в организме. Для него не найден специфический переносчик, но обнаружен белок с молекулярной массой 40 кД, с которым он связывается в печени (1). При этом исходное для биосинтеза хинона вещество — α-токоферол связан с другим белком. α-Токоферилхинон часто рассматривается как активный метаболит витамина Е (2). Показано, при внутримышечном или оральном введении экзогенного α-токоферилхинона происходит торможение роста животных. Механизм такого торможения может заключаться в шунтировании α-токоферилхиноном дыхательной цепи между НАДН-дегидрогеназой и молекулярным кислородом (3).
Поскольку дрожжи Saccharomyces cerevisiae с хорошо изученной генетической системой обладают чертами, характерными для эукариотических клеток, они могут служить идеальной моделью для изучения регуляторных систем у животных и человека в отличие от классического прокариотического объекта клеточной биологии — Escherichiacoli (4, 5).
Целью нашей работы было изучение влияния α-токоферилхинона на процессы брожения у пекарских дрожжей Saccharomycescerevisiae.
Методика. В каждом опыте использовали коммерческие пекарские дрожжи Saccharomycescerevisiae(штамм S11M, XTP) производства «Lesaffre» (Франция) (сухие свежеприготовленные и хранившиеся в течение года в холодильнике при +4 °С во вскрытых заводских упаковках). Опыты проводили в 3 повторностях. D,L-α-токоферилхинон получали из D,L-α-токоферола окислением азотной кислотой (6) с последующей очисткой на колонке из силикагеля под контролем высокоэффективной жидкостной хроматографии (ВЭЖХ) (7).
В опыте 1 4 г D-глюкозы растворяли в 200 мл воды, разливали по 100 мл в две колбы объемом 300 мл, в одну вводили 10 мг a-токоферил-хинона, солюбилизированного в 10 мл воды с 150 мг Твин-80, в другую (контроль) — такое же количество воды и Твин-80. В каждую колбу помещали по 0,75 г дрожжей и активно перемешивали на водяной бане с шейкером (модель 357, «Olpon», Польша) при 30 °С в течение 48 ч, отбирая пробы по предварительно отработанной схеме (первый образец — через 12 мин после добавления дрожжей, следующие — через 3,5; 7; 14; 24; 36 и 48 ч).
В опыте 2 1 г пирувата натрия растворяли в 200 мл воды, разливали по 100 мл в две колбы объемом 300 мл, в одну добавляли то же количество α-токоферилхинона (контроль — без a-токоферилхинона), затем в обе — по 1 г дрожжей. Образцы инкубировали на водяной бане с шейкером. Спустя 1 сут в колбы вносили по 250 мг пирувата натрия, отбирали пробы по 5 мл (через 12 мин; 3,5; 7; 14; 24; 36 и 48 ч), к пробам в качестве осадителя белка добавляли 1,25 мл 10 % раствора HPO3 и центрифугировали в рефрижераторной центрифуге К 24 («Janetzki», Германия) с угловым ротором (12x11 мл, 28°) при 10 000 g и +4 °С.
Количество пировиноградной кислоты оценивали по ее способности образовывать 2,4-динитрофенилгидразоны, которые очищали обратной экстракцией из толуольного раствора в водный раствор соды (Na2CO3) и переводили в аци-форму добавлением раствора едкого натра (8). Количество α-токоферилхинона определяли с помощью прямофазной ВЭЖХ на хроматографе Милихром (Россия) со стальной колонкой 120x2 мм, заполненной силасорбом-600 («Lachema», Чешская Республика); диаметр частиц — 6 мкм, элюент — смесь гексан:серный эфир:метанол (89:10:1) (7).
Концентрацию гексоз измеряли методом, основанным на их дегидратации с образованием формальдегида, количество которого учитывали по реакции с хромотроповой кислотой (9). Уксусный альдегид выделяли в виде 2,4-динитрофенилгидразона из толуольного раствора, оставшегося после экстракции 2,4-динитрофенилгидразона пировиноградной кислоты. Раствор упаривали и подвергали двойной очистке хроматографией со свидетелем в тонком закрепленном слое силикагеля L («Lachema», Чешская Республика), используя бензол в качестве носителя. Гидразон экстрагировали метанолом. Количество альдегида определяли с помощью спектрального колориметра Specol-11 («Carl Zeiss», Германия) при 350 нм.
В работе использовали реагенты отечественного производства с квалификациями «хч» или «чда».
Статистическую обработку проводили на основе общепринятых принципов (10) с использованием программного обеспечения Microsoft (Microsoft Exсel).
Результаты. В варианте со свежеприготовленными дрожжами в опыте 1 добавление α-токоферилхинона приводило к заметному снижению концентрации пировиноградной кислоты в инкубационной среде (рис. 1, А, а), в опыте 2 — к резкому уменьшению скорости утилизации исходного вещества по сравнению с контролем (см. рис. 1, А, б). В опыте 1 разница между усредненными показателями составляла +20,2±0,27 % (Р = 0,014), в опыте 2 — +34,6±8,80 % (Р = 0,054).
Хранение дрожжей заметно повлияло на их способность сбраживать глюкозу в условиях активного перемешивания инкубационной среды. При добавлении α-токоферилхинона (см. рис. 1, Б, а) после 14 ч инкубации наблюдали уменьшение содержания пирувата (+17,2±0,5 %; Р = 0,007). Концентрация вещества в среде между 36-м и 48-м ч инкубации была минимальной и не менялась. Эти данные свидетельствуют об ускорении брожения под влиянием α-токоферилхинона. В опыте с пируватом также отмечали усиление метаболизации (см. рис. 1, Б, б) (+45,2±5,4 %, Р = 0,025).
В опыте 1 в варианте со свежими дрожжами не наблюдали заметных отличий от контроля по динамике содержания гексоз, количество которых резко падало к 3,5 ч без существенных дальнейших изменений (рис. 2, А). При использовании долго хранившихся дрожжей концентрация уменьшалась постепенно, что свидетельствует о потере активности в процессе хранения (см. рис. 2, Б).
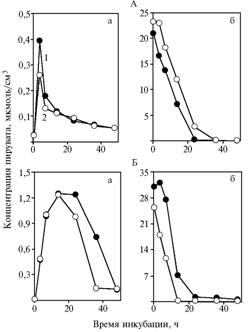 |
Рис. 1. Концентрация пирувата в инкубационной среде с глюкозой (а) и пируватом натрия (б) для сухих свежих (А) и подвергшихся длительному хранению (Б) дрожжей Saccharomycescerevisiae: 1 — без добавления в среду α-токоферилхинона (контроль), 2 — с добавлением в среду α-токо-ферилхинона. |
Отсутствие заметной разницы между контролем и опытом по метаболизации глюкозы демонстрирует, что ускорение брожения под влиянием α-токоферилхинона происходило на второй стадии этого процесса.
Наиболее вероятным объектом атак окислителей служат сульфгидрильные группы, в частности cys-149 в активном центре фосфорилирующей глицер-альдегид-3-фосфатдегидрогеназы (ГАФД, КФ 1.2.1.12). Обработка ГАФД перекисью водорода или старение фермента ведут к образованию неактивного «окисленного» фермента, не обладающего поглощением при 360 нм. При восстановлении, например в присутствии глутатиона (11), возвращается исходная активность и спектр поглощения. Мягкое окисление SH-группы cys-149 пероксидом водорода в концентрации 100 мкМ инициирует, в частности, образование сульфеновой кислоты (12) и, как следствие, снижение дегидрогеназной и появление ацилфосфатазной активностей, что в итоге ускоряет гликолиз с уменьшением выхода АТФ. Окисление сульфгид-
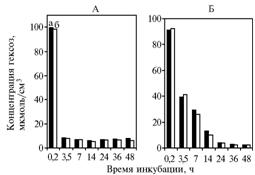 |
Рис. 2. Концентрация гексоз в инкубационной среде с глюкозой для сухих свежих (А) и подвергшихся длительному хранению (Б) дрожжей Saccharomycescerevisiae: а — без добавления в среду α-токоферилхинона (контроль), б — с добавлением в среду α-токоферилхинона. |
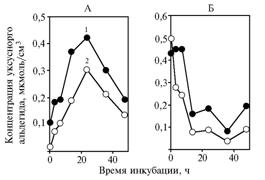 |
Рис. 3. Концентрация уксусного альдегида в инкубационной среде с глюкозой (А) и пируватом натрия (Б) для подвергшихся длительному хранению дрожжей Saccharomycescerevisiae: 1 — без добавления в среду α-токоферилхинона (контроль), 2 — с добавлением в среду α-токоферилхинона. |
Как в опыте 1, так и в опыте 2 в варианте с продолжительно хранившимися дрожжами при добавлении α-токоферилхинона к инкубационной среде концентрация уксусного альдегида заметно уменьшалась (рис. 3) (соответственно +42,0±1,7 % при Р = 0,0003 и +31,5±0,35 % при Р = 0,002). Количество уксусного альдегида в среде инкубации в этих условиях составляло около 1 % от неиспользованного пирувата.
Переход добавленного в среду a-токоферилхинона в клетки дрожжей, выявляемый после центрифугирования образцов, инкубируемых с глюкозой в качестве субстрата (рис. 4, А), с течением времени несколько усиливался при снижении концентрации вещества в супернатанте.
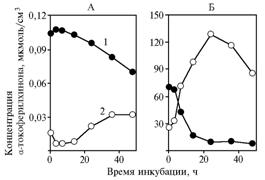 |
Рис. 4. Концентрация a-токоферилхинона в инкубационной среде (1) и в осадках (2) при использовании в качестве субстрата для Saccharomycescerevisiae глюкозы (А) и пирувата натрия (Б). |
При использовании пирувата в качестве субстрата различия были существенно более контрастными (см. рис. 4, Б), но характер наблюдаемых изменений сохранялся. То же отмечали в варианте с дрожжами после долгого хранения (данные не приведены).
Участие α-токоферилхинона в изучаемых процессах, возможно, состоит в его превращении в вещество с меньшей хроматографической подвижностью (объем элюента, выводящего пик вещества в кювету прибора, — 398 мкл против 900 мкл). Изменение концентрации этого производного α-токоферилхинона в зависимости от времени инкубации (рис. 5) повторяет динамику концентрации исходного соединения в дрожжевых центрифугатах (для пирувата — r = 0,97, P < 0,05; для глюкозы — r = 0,95, P < 0,05). Синтез предполагаемого активного агента происходит с участием структур клеток S. cerevisiae. При этом средняя концентрация a-токоферилхинона в дрожжевых осадках в значительной степени зависит от субстрата (для пирувата — 0,132 мкмоль/см3; для глюкозы — 0,017 мкмоль/см3).
Причиной «утечки» α-токоферилхинона в опыте 1 могло быть либо разрушение в процессе брожения, либо превращение в неустановленное вещество. По УФ-спектру поглощения, близкому к характерному для a-токоферилхинона (рис. 6), это вещество, вероятнее всего, представляет собой неполярный α-токоферилхинон — 3-дезокси-α-токоферил-п-хинон.
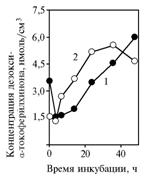 |
Рис. 5. Концентрация неполярного a-токоферилхинона в клетках дрожжей Saccharomycescerevisiae при использовании в качестве инкубационной среды глюкозы (1) и пирувата натрия (2). |
Например, показано, что 3-дезокси-a-токоферил-п-хинон содержится в экстракте Buty-rivibriofibrisolvens как альтернатива аутентичному a-токоферилхинону и синтезируется из него, а частично очищенный препарат фермента, выделенного из бактерии, катализирует восстановление a-токоферилхинона в соответствующий хинол в присутствии НАДН со стехиометрией 1:1 (15).
В интактных дрожжевых клетках мы не обнаружили ни аутентичного, ни неполярного α-токоферилхинона. Тем не менее, сам факт биосинтеза неполярного хинона из добавленного указывает на то, что дрожжи сохраняют способность метаболизировать α-токоферилхинон, который может поступать из натуральных субстратов, в частности на начальных этапах аэробного разложения силосов, которые протекают за счет дрожжей (16).
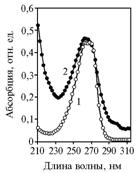 |
Рис. 6. УФ-спектры в остановленном потоке элюента для аутентичного a-токоферилхинона (1) и продукта его превращения дрожжамиSaccharomycescerevisiae (2). |
Таким образом, α-токоферилхинон активно воздействует на процессы брожения на стадиях после образования фосфотриоз, причем характер влияния на метаболизм экзогенного пирувата у сухих свежих и подвергшихся хранению на воздухе при 4 °С пекарских дрожжей Saccharomycescerevisiae различается по скорости метаболизации. Следует отметить, что в других партиях реакция свежих и хранившихся в холодильнике дрожжей на введение D,L-α-токоферилхинона была гораздо менее контрастной (данные не приведены). Следовательно, сила влияния α-токоферилхинона на гликолиз зависит от характера метаболизма в дрожжевой (или животной) клетке, в частности от конкурентных взаимоотношений между НАД и НАДН в дегидрогеназных (изоцитратдегидрогеназная, алкогольдегидрогеназная) реакциях.
Л И Т Е Р А Т У Р А
1. A r i t a M., S a t o Y., A r a i F. e.a. Binding of α-tocopherylquinone, an oxidized form of a-tocopherol to glutathione-S-transferase in the liver cytosol. FEBS Lett., 1998, 436(3): 424-426.
2. Д о н ч е н к о Г.В. Биохимия убихинона. Киев, 1988.
3. Д у д и н В.И, Г р и щ у к С.В., С а к о в ц е в а Т.В. Об участии α-токоферилхи-нона и убихинона в регуляции процессов роста у свиней и птицы. С.-х. биол., 2008, 6: 53-58.
4. Б е р р и Д. Биология дрожжей. М, 1985.
5. З в я г и л ь с к а я Р.А., К о т е л ь н и к о в а А.В. Структура и функциональная активность дрожжевых митохондрий. М., 1991, т. 36.
6. R a o G.H.R., K e i c h T.P., W h i t e J.G. Preparation, separation and characterization of vitamin E quinone. J. Chromat., 1980, 196: 506-511.
7. Д у д и н В.И. Биохимия витамина Е и связанных с ним биологически активных веществ. М., 2004.
8. П е т р у н ь к и н а А.М. Практическая биохимия. Л., 1961: 384-390.
9. К о р е н м а н И.М. Методы определения органических соединений. М., 1970: 65-66.
109. М е р к у р ь е в а Е.К. Биометрия в животноводстве. М., 1964.
11. Р э к е р Э. Биоэнергетические механизмы. М., 1967.
12. Д а н ь ш и н а П.В., Ш м а л ь г а у з е н Е.В., А р у т ю н о в Д.Ю. и др. Ускорение гликолиза нефосфорилирующей и окисленной фосфорилирующей глицеральдегид-3-фосфатдегидрогеназами. Биохимия, 2003, 68(5): 725-733.
13. A s a h i M., F u j i i J., S u z u k i K. e.a. Inactivation of glutathione peroxidase by nitric oxide. Implication for cytotoxicity. J. Biol. Chem., 1995, 270(36): 21035-21039.
14. У а й т А., Х е н д л е р Ф., С м и т Э. и др. Основы биохимии. М., 1981., т. 2.
15. H u g h e s P.E., T o v e S.B. Identification deoxy-α-tocopherolquinol as another endogenous electron donor for biohydrogenation. J. Biol. Chem., 1980, 255(24): 11802-11806.
16. М к Д о н а л ь д П. Биохимия силоса. М., 1985.
ABOUT PARTICIPATION OF α-TOCOPHERILQUINONE IN GLUCOSE FERMENTATION BY Saccharomyces cerevisiae
V.I. Dudin
The author studies the effect of α-tocopherilquinone on fermentation processes in cultures of dry freshly-prepared and long-term keeping baker's yeast Saccharomyces cerevisiae in incomplete medium with D-glucose or sodium pyruvate. It was established, that α-tocopherilqui-none takes an active part in fermentation of the second stage of glycolysis. The utilization of exogenous pyruvate reduces in experiments with dry fresh baker’s yeast, but raises in experiments with yeast after its keeping.
Key words: dried baker`s yeast Saccharomyces cerevisiae, glucose, acetic aldehyde, α-tocopherolquinone, 3-deoxy-α-tocopherolquinone, pyruvic acid, fermentation.
ГНУ Всероссийский НИИ физиологии, |
Поступила в редакцию |
Оформление электронного оттиска
![]()
![]()




