doi: 10.15389/agrobiology.2023.1.158rus
УДК 633.111.1:632.4:632.937.15:581.1
Работа выполнена в рамках государственного задания в соответствии с тематическим планом ВИР по проекту № 0481-2022-0001 «Структурирование и раскрытие потенциала наследственной изменчивости мировой коллекции зерновых и крупяных культур ВИР для развития оптимизированного генбанка и рационального использования в селекции и растениеводстве».
УСТОЙЧИВОСТЬ К БОЛЕЗНЯМ, ПРОДУКТИВНОСТЬ И СОДЕРЖАНИЕ ФОТОСИНТЕТИЧЕСКИХ ПИГМЕНТОВ В ЛИСТЬЯХ ЯРОВОЙ МЯГКОЙ ПШЕНИЦЫ (Triticum aestivum L.) ПОД ВЛИЯНИЕМ ПОЛИФУНКЦИОНАЛЬНЫХ БИОПРЕПАРАТОВ И КОМПЛЕКСОВ НА ОСНОВЕ МИКРООРГАНИЗМОВ И ХИТОЗАНА
И.И. НОВИКОВА1 ✉, Э.В. ПОПОВА1, Л.Е. КОЛЕСНИКОВ2, Ю.Р. КОЛЕСНИКОВА3, С.С. ЧЕКУРОВА2
Использование полезных микроорганизмов и биологически активных молекул составляет основу современной концепции фитосанитарной оптимизации агроэкосистем. Усиление защитных свойств препаративных форм, в состав которых входят антагонисты фитопатогенов и хитозан, обусловлено способностью полисахарида хитозана индуцировать системную болезнеустойчивость растений. Кроме того, полифункциональные композиции с множественным механизмом действия, эффективные против широкого спектра фитопатогенов, могут положительно влиять на функциональное состояние растений, в том числе на их фотосинтетическую активность, количественные и качественные изменения всей пигментной системы, которые часто отражают характер адаптивных реакций при стрессе. Однако исследования изменений фотосинтетического аппарата во взаимосвязи с болезнеустойчивостью и продуктивностью растений под влиянием таких композиций крайне немногочисленны. В настоящей работе впервые показано, что полифункциональные комплексы Витаплан, КЖ + Хитозан II и Витаплан, КС + Хитозан II существенно повышают продуктивность и болезнеустойчивость пшеницы, при этом содержание хлорофиллов a и b в листовых пластинках также оказалось наибольшим. Соотношение содержания хлорофиллов a+b и каротиноидов, служащее одним из показателей стрессоустойчивости растений, было максимальным при применении комплекса Витаплан, КЖ + Хитозан II. Цель нашего исследования — оценить потенциальную продуктивность пшеницы по морфометрическим показателям развития растений, их поражаемость наиболее вредоносными болезнями (корневая гниль, бурая и желтая ржавчины, мучнистая роса, септориоз) и содержание хлорофиллов a, b, каротиноидов в листьях при применении полифункциональных биопрепаратов и комплексов, объединяющих полезные свойства микроорганизмов — антагонистов фитопатогенов и активатора болезнеустойчивости растений хитозана. Семена мягкой яровой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 (к-64900, коллекция ВИР) перед посевом обрабатывали биопрепаратами на основе штаммов Bacillus subtilis ВКМ В-2604D и B. subtilis ВКМ В-2605D Витаплан, СП, Витаплан, КЖ и комплексами Витаплан, КЖ + Хитозан II, Витаплан, КС + Хитозан II. В полевых условиях этими же препаратами опрыскивали вегетирующие растения в сравнении с контролем без обработки. В целом использованные комплексы оказались эффективнее биопрепаратов на 16,2 %. Применение полифункциональных композиций существенно (при р < 0,05) снижало пораженность растений пшеницы комплексом болезней на 17,9 %. Наибольшие значения потенциальной продуктивности (0,94±0,02 г/растение) и содержания в листьях хлорофиллов a (1,32±0,02 мг/г) и b (2,15±0,04 мг/г) установлены в варианте опыта с применением полифункционального комплекса Витаплан, КЖ + Хитозан II, что на 57,1 %; 16,7 % и 4,3 % больше, чем в контроле (в сравнении с остальными вариантами — больше на 19,7 %; 23,7 % и 11,0 %). Выявлены различия в содержании фотосинтетических пигментов: хлорофилла a и хлорофилла b во флаговых листьях пшеницы при применении полифункционального комплекса Витаплан, КЖ + Хитозан II по сравнению c биопрепаратами на 16,8 %; 3,7 % и 2,0 %, с вариантом Витаплан, КС + Хитозан II — соответсвенно на 1,1 %, 17,7 % и 27,0 %. Наиболее сильная корреляция выявлена между содержанием хлорофилла b во флаговых листьях и продуктивностью пшеницы (r = 0,69, р = 0,03), содержанием хлорофилла b во флаговых листьях и числом зерен в колосе (r = 0,79, р = 0,006), массой зерен с колоса и массой колоса (r = 0,69, р = 0,03; r = 0,72, р = 0,02). Выявлены корреляционные связи между снижением развития желтой ржавчины и увеличением содержания хлорофиллов a и b в листьях (r = -0,66, р = 0,04; r = -0,87, р = 0,005). Наибольшие значения соотношения содержания хлорофиллов a и b в листьях по сравнению с контролем отмечены при применении комплексов Витаплан, КЖ + Хитозан II и Витаплан, КС + Хитозан II. Соотношение содержания хлорофиллов a и b и содержания пигментов группы каротиноидов как одного из показателей устойчивости растений к внешним неблагоприятным факторам достигало максимальных значений также в варианте опыта Витаплан, КЖ + Хитозан II. По сумме показателей наиболее перспективен при возделывании пшеницы полифункциональный комплекс Витаплан, КЖ + Хитозан II, обладающий выраженным ростостимулирующим и защитным действием при профилактическом применении. Работ по созданию композиций, сочетающих биопрепараты и индукторы болезнеустойчивости, ранее не проводилось. Эффективность таких композиций может зависеть от устойчивости сортов культурных растений, однако, как показывает наш опыт, сочетание подавляющего воздействия на фитопатогенные микроорганизмы и стимулирующего — на растительный организм обеспечивает более надежный защитный эффект.
Ключевые слова: Triticum aestivum, мягкая пшеница, полифункциональные биопрепараты, хлорофилл a, хлорофилл b, каротиноиды, продуктивность, бурая ржавчина, желтая ржавчина, септориоз, мучнистая роса, корневая гниль.
Современная технология возделывания зерновых культур предусматривает проведение комплекса мероприятий по защите от болезней, в частности химическое протравливание семян и опрыскивание посевов фунгицидами, что экологически небезопасно и приводит к формированию устойчивых популяций фитопатогенов. Поэтому необходимы альтернативные экологически малоопасные средства (1).
Штаммы Bacillus subtilis благодаря разнообразию и высокой изменчивости их биохимических свойств и синтезу спектра биоактивных метаболитов — циклических липопептидов, полипептидов, белков и непептидных соединений (2-4) широко используются в борьбе с возбудителями заболеваний сельскохозяйственных культур. Известно, что штаммы B. subtilis способны продуцировать три рибосомных антибиотика (TasA, субтилозин и сублацин), четыре нерибосомных антибиотика (бацитрацин, бацилизин, плипастатин и сурфактин), новый фосфолипидный антибиотик бацилизоцин и аминосахарный антибиотик неотрегалосадиамин (NTD) (2). Нерибосомально синтезированные циклические олигопептиды, такие как сурфактин, соединения итуриновой группы и фенгицин, содержащие цепь жирных кислот, проявляют высокую антифугальную и антибактериальную активность (5). Штаммы B. subtilis продукируют различные гидролитические ферменты, разрушающие клеточную стенку фитопатогенных грибов (6). Ряд активных соединений, образуемых этими микроорганизмами, обладают элиситорной активностью и запускают механизмы индуцированной устойчивости (7, 8).
Также опубликованы многочисленные данные относительно способности полезных микроорганизмов ризо- и филлосферы синтезировать метаболиты, которые влияют на устойчивость и рост растений и обладают сигнальными и гормональными функциями. Ауксины, гиббереллины, цитокинины, абсцизовая (АБК), салициловая, жасмоновая кислоты — природные регуляторы роста (9-11). Многие штаммы бактерий, относящихся к родам Bacillus, Azospirillium, Pseudomonas, могут синтезировать ауксины, что стимулирует развитие корневой системы растения, как следствие, активируется поглощение воды и питательных веществ. Указанные процессы в совокупности повышают устойчивость к болезням и позволяют быстрее проходить те стадии развития, когда растения наиболее восприимчивы к патогенам (12-14).
Наиболее перспективными для защиты сельскохозяйственных культур и повышения их урожайности преставляются композиции, объединяющие полезные свойства микроорганизмов — антагонистов возбудителей болезней и такх активаторов болезнеустойчивости растений, как хитозан и его производных (15). Хитозан и препараты на его основе нашли практическое применение в качестве индукторов, повышающих устойчивость к грибным, бактериальным и вирусным болезням (16-19).
Биологическая активность хитозана как индуктора устойчивости определяется его способностью активировать защитные реакции и индуци-ровать неспецифический клеточный иммунитет — одну из линий защиты врожденного иммунитета растений (19, 20). Хитозан и хитин, присутствующие в клеточных стенках многих паразитарных микроорганизмов и грибов, — молекулярные паттерны фитопатогенных грибов (pathogen-associated molecular patterns, PAMPs), которые распознаются белковыми рецепторами растений (pattern-recognition receptors, PRR) (21) и служат сигналом для активации ответных защитных реакций (pattern-triggered immunity, РTI), предотвращающих развитие инфекции (22, 23). Защитные реакции, вызываемые хитозаном, включают повышение концентрации Ca2+ в цитозоле, окислительный взрыв (образование активных форм кислорода — АФК) (24-26), гиперчувствительный ответ (ГО), синтез белков, связанных с патогенезом (PR, pathogenesis related proteins), с антимикробной и литической активностью, индукцию гормонов защиты (абсцизовая кислота, жасмонаты, салициловая кислота), образование фитоалексинов (26-31). Кроме того, для реакций защиты, индуцируемых хитозаном в растениях, характерно усиление синтеза лигнина (32) и каллозы (30), что приводит к укреплению клеточных стенок и созданию физических барьеров на пути проникновения и передвижения патогенов в растениях (18). Эти реакции, направленные на подавление роста патогена, приводят к формированию у растений системной устойчивости и защищают их от последующей атаки широкого спектра патогенов, а также повышают устойчивость растений к неблагоприятным абиотическим факторам.
Механизмы защитного действия хитозана и его высокая эффективность при защите различных видов сельскохозяйственных растений от действия широкого спектра фитопатогенов подробно рассмотрены в многочисленных обзорных статьях (17, 30). Помимо защиты от фитопатогенов, хитозан ускоряет рост растений, повышает устойчивость к стрессам (заморозки, засуха, излишняя влага) и продуктивность зерновых и овощных культур (33-36). Обработка полисахаридом повышает скорость фотосинтеза, увеличивает число побегов, размеры листьев, высоту растений у пшеницы, кукурузы, фасоли, томата, риса, что обеспечивает повышение урожайности (37-42).
Среди факторов, определяющих высокую зерновую продуктивность пшеницы, выделяют хлорофилльный фотосинтетический потенциал посева, который характеризует суммарное количество хлорофилла в листьях или целых растениях на единице площади посева за вегетацию или ее определенный период. Отмечается тесная положительную связь между хлорофилльным фотосинтетическим потенциалом в течение репродуктивного периода и урожаем пшеницы (43). Установлено (44), что увеличение урожайности современного сорта озимой пшеницы Фаворитка по сравнению с сортом селекции 1960-х годов Мироновская 808 связано с повышением содержания и валового количества хлорофилла, а также удлинением периода функционирования фотосинтетического аппарата посева в репродуктивный период. Современный сорт характеризуется более эффективным использованием поглощенной световой энергии, что приводит к росту фотосинтетической продуктивности. Авторы делают вывод, что повышение эффективности фотосинтеза — перспективная стратегия увеличения продуктивности зерновых культур (44). Оптимизация работы фотосинтетического аппарата на разных уровнях его организации способна повысить зерновую продуктивность на 10-60 % (45, 46).
У растений хлорофиллы встречаются только в пигмент-белковых комплексах (ПБК), поскольку в свободном виде, будучи сильнейшими фотосенсибилизаторами, они могут вызвать разрушение мембран тилакоидов и стромы хлоропласта из-за фотодинамического эффекта. ПБК позволяют оптимизировать работу фотосинтетического аппарата. Выделяют четыре главных типа ПБК: два из них — это реакционные центры фотоситемы (ФС) I и ФС II, два другие – ССК (светособирающие комплексы) ФС I и ФС II. В хлоропластах антенный комплекс содержит большое число (200-400) молекул хлорофилла и относительно небольшое количество каротиноидов, нековалентно связанных с белком.
Хлорофилл b служит вспомогательным светособирающим пигментом, улавливая и передавая световую энергию в реакционные центры фотосистем. На его долю приходится примерно 15-25 % от общего содержания хлорофиллов. В отличие от хлорофилла а, который входит в состав коровых комплексов фотосистем, хлорофиллb находится только в светособирающих комплексах фотосистем (ССК I и ССК II) и в так называемой малой антенне ФС II. В ССК I хлорофилл b составляет около 22 % от суммарного количества хлорофиллов, в ССК II — около 43 %, в пигмент-белковом комплексе малой антенны — 31-46 % (47).
Каротиноиды — вспомогательные фотосинтетические пигменты и обязательный компонент пигментных систем всех фотосинтезирующих организмов. В хлоропластах каротиноиды находятся в составе ПБК и отчасти в липофильной фазе тилакоидных мембран. Реакционные центры, которые представляют собой комплекс белков, пигментов и других кофакторов и обеспечивают реакцию превращения энергии света в химическую при фотосинтезе, включают только хлорофилл аи β-каротин, а светособирающие антенны — хлорофиллы а и b, каротины и ксантофиллы. Каротиноиды, которые входят в состав светособирающей антенны, расширяют спектральный диапазон фотосинтетически активной радиции (ФАР). Кроме участия в поглощении энергии солнечного излучения и ее миграции от дополнительных пигментов к основным, каротиноиды также выполняют защитную функцию (гашение триплетного хлорофилла и синглетного кислорода), предохраняя фотосинтетический аппарат от фотодеструкции.
Ранее биосинтез фотосинтетических пигментов, в частности хлорофиллов, не считался фактором, активирующим сигнальные пути, которые приводят к инициации или задержке фаз онтогенеза. Однако в недавних работах показана важная роль вспомогательного фотосинтетического пигмента хлорофилла b в регуляции онтогенеза растений (48). Кроме того, опубликованы данные, которые свидетельствуют, что отсутствие хлорофилл b негативно влияет на смену периодов онтогенеза у ячменя. Мутанты chlorina обоих видов отличались от растений родительских линий более поздним наступлением флоральной трансформации. Кроме того, у 30-40 % мутантов ячменя останавливался рост и дифференцировка структурных элементов колоса (49). Фотосинтетические структуры могут участвовать в адаптивном ответе растения на стрессовые воздействия (50). При этом вероятно изменение содержания пигментов (хлорофилла а, хлорофилла b, суммы хлорофиллов а + b и каротиноидов). Абиотические стрессы снижают эффективность фотосинтеза прежде всего из-за негативного влияния на биосинтез хлорофилла, работу фотосистем, механизмы транспорта электронов, параметры газообмена (51, 52).
Следует отметить, что некоторые исследователи либо не выявили прямой связя между эффективностью фотосинтеза и урожаем, либо установили отрицательную корреляцию между этими показателями у многих растений, в том числе у зерновых культур (53).
Тем не менее фотосинтез — основа первичной биопродуктивности как в природных экосистемах, так и при формировании урожая сельскохо-зяйственных культур, следовательно, в изменяющихся условиях среды требуется сохранять и увеличивать фотосинтетическую продуктивность растений (54-56).
Косвенно устойчивость к абиотическим стрессам можно оценить по количественным изменениям пигментного комплекса (57-60). В ряде работ изучено влияния поражения растений возбудителями болезней на состав пигментного комплекса. В частности, при поражении древесных растений паразитическими грибами происходит снижение содержания хлорофиллов и увеличение содержания каротиноидов (61). В результате происходят структурно-функциональные и физиолого-биохимические перестройки, что в итоге приводит к снижению фотосинтетической активности (61). Однако подобные исследования, связывающие продуктивность и устойчивость растений к биотическому стрессу (поражени фитопатогенами) с состоянием фотосинтетического комплекса крайне немногочисленны.
В представленной работе мы впервые показали, что полифункциональные комплексы Витаплан, КЖ + Хитозан II и Витаплан, КС + Хитозан II существенно повышают продуктивность и болезнеустойчивость пшеницы, при этом содержание хлорофиллов a и b в листовых пластинках также оказалось наибольшим. Соотношение содержания хлорофиллов a + b и каротиноидов, служащее одним из показателей стрессоустойчивости растений, было максимальным при применении комплекса Витаплан, КЖ + Хитозан II.
Цель нашего исследования — оценить потенциальную продуктивность пшеницы по морфометрическим показателям развития растений, их поражаемость наиболее вредоносными болезнями (корневая гниль, бурая и желтая ржавчины, мучнистая роса, септориоз) и содержание хлорофиллов a, b, каротиноидов в листьях при применении полифункциональных биопрепаратов и комплексов, объединяющих полезные свойства микроорганизмов-антагонистов фитопатогенов и активатора болезнеустойчивости растений хитозана.
Методика. Экспериментальные исследования осуществляли в условиях опытного поля (ФГБНУ ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова — ВИР, г. Санкт-Петербург—Пушкин, 2016-2018 годы) на яровой мягкой пшенице (Triticum aestivum L.) сорта Ленинградская 6 (к-64900, предоставлен отделом генетических ресурсов пшениц ВИР). В полевом опыте пшеницу высевали на делянках площадью 1,0 м2 рядовым способом с междурядьями 15 см и расстоянием в ряду 2 см. Для каждого образца учетная делянка состояла из 6 рядков, в каждый рядок при посеве помещали 50 зерен. Норма высева составила 300 семян на 1 м2. Глубина заделки семян 5-6 см. Полевой опыт был заложен в 3-кратной повторности. Все мероприятия проводили согласно общепринятым рекомендациям и методикам ВИР.
Схема опыта предусматривала следующие варианты: без обработки (контроль); биопрепарат Витаплан, СП (смачивающийся порошок, стандарт), содержащий клетки штаммов Bacillus subtilis ВКМ В-2604D и B. subtilis ВКМ В-2605D (титр жизнеспособных клеток 1011 КОЕ/г); Витаплан, КЖ — культуральная жидкость штаммов B. subtilis ВКМ В-2604D и B. subtilis ВКМ В-2605D при соотношении 1:1 с титром жизнеспособных клеток B. subtilis 1010 КОЕ/мл; полифункциональный комплекс Витаплан, КЖ + Хитозан II (состав культуральной жидкости в полифункциональном комплексе Витаплан, КЖ + Хитозан II и в препарате Витаплан, КЖ одинаков); полифункциональный комплекс Витаплан, КС (концентрат суспензии, титр жизнеспособных клеток 5×1010 КОЕ/мл) + Хитозан II. Хитозан II с молекулярной массой 50 КДа был получен методом окислительной деструкции (62) из хитозана с молекулярной массой 300 КДа и степенью деацетилирования 85 % (ООО «Биопрогресс», Россия). Хитозан растворяли в янтарной кислоте и при постоянном перемешивании вводили в культуральную жидкость до конечной концентрации 0,1 % (63).
Схема полевого опыта включала обработку семян пшеницы сорта Ленинградка 6 перед посевом и 4-кратное опрыскивание теми же препаратами вегетирующих растений в фазы кущения, выхода в трубку, колошения и цветения.
При анализе элементов продуктивности пшеницы использовали 19 показателей, которые характеризуют морфологические признаки растений и структуру урожая в фазы колошения—цветения и созревания. В фазы колошения—цветения учитывали следующий комплекс показателей: продуктивную и общую кустистость (шт.), площадь флагового и предфлагового листа (см2), высоту растений (см), длину колоса (см), число колосков в колосе (шт.), массу колоса (г.). Кроме того, определяли число и длину корней (главного зародышевого корня, зародышевых и колеоптильных корней), отходящих от эпикотиля, учитывали число и длину узловых корней. Определяли массы корней и вегетативной части растений пшеницы (массу вегетативной части рассчитывали по суммарной массе стебля и листьев без учета массы корней и колоса).
Дополнительно определяли полевую всхожесть образцов (64) и фазу онтогенеза растений по общепринятой шкале J.C. Zadoks (65). В фазу созревания (стадия полной спелости) изучали структуру урожая пшеницы по показателям число колосков в колосе, (шт.), длина колоса (см), масса колоса с зерном (г), число зерен в колосе (шт.), масса зерен с колоса (г), масса 1000 зерен (г).

Интенсивность поражения пшеницы возбудителями болезней листьев определяли с использованием общепринятых шкал учета, а также по дополнительным показателям патогенеза (68). Интенсивность или степень развития ржавчины определяют в процентах по классическим шкалам (63): бурой ржавчины (Puccinia recondita Rob. ex Desm f. sp. tritici – Русакова или Петерсона, желтой ржавчины (Puccinia striiformes Westend) — Маннерса, септориоза (Septoria tritici Roberge in Desmaz.) — Джеймса, мучнистой росы (Blumeria graminis (DC.) Speerf. sp. tritici March) — Петерсона. Кроме того, определяли показатели, дополнительно характеризующие патогенеза: число пустул (суммарное на лист), число полос с пустулами, длину полос с пустулами, площадь пустулы и их число в полосе. Показатели патогенеза учитывали в лабораторных условиях с использованием стереоскопического микроскопа МБС-9 («ООО ПТП «АСМА-Прибор», Россия) и тринокулярного микроскопа Микромед 1 (вар. 3 LED)» (OOO «Наблюдательные приборы», Россия).
Размер инфекционных структур возбудителей болезней, формируемых на листьях при патогенезе (пятна, пустулы и т.п.) определяли с помощью объект- и окулярного микрометров. Значения площади пустулы рассчитывали в предположении об их эллиптической форме с использованием выражения: S = m·pab, где а и b — длина полуосей эллипса (в линиях окулярного микрометра), m — масштабный коэффициент микроскопа.
Содержание хлорофиллов a и b во флаговых листьях пшеницы определяли спектрофотометрически согласно описанию (69) с использованием спектрофотометра SPEKOL-11 («Carl Zeiss AG», Германия). Для определения содержания пигментов фотосинтеза использовали навески из средней части листовой пластинки, из которых готовили спиртовые вытяжки (69).

Статистический анализ результатов проводили в программах IBM SPSS Statistics 21, Statistica 6.0 («StatSoft, Inc.», США), Microsoft Excel 2016. При расчетах использовали методы дисперсионного анализа и критерий множественных сравнений Шеффе, методы параметрической статистики (расчет средних М и их стандартных ошибок ±SEM, 95 % доверительных интервалов и t-критерия Стьюдента), непараметрический метод ранговых корреляций Спирмена, линейный и нелинейный регрессионный анализ, основанный на алгоритме метода наименьших квадратов.
Результаты. В схеме полевого опыта вариант с Хитозаном II с молекулярной массой 50 КДа мы отдельно не использовали, так как проведенные нами ранее полевые опыты показал, что его применени вызывало достоверный рост только массы вегетативной части растений (63). Комплекс Витаплан, КЖ + Хитозан II вызывал достоверное повыешени наибольшего числа показателей продуктивности пшеницы по сравнению с контролем (63). В этом же варианте опыта был зарегистрирован максимальный достоверный прирост урожайности (82,6 %) и максимальная эффективность против гельминтоспориозной корневой гнили (снижение развития корневой гнили на 80-100 % по сравнению с контролем) (63).
Полученные в нстоящей работе результаты показали существенное положительное влияние изученных полифункциональных композиций на фитосанитарное состояние агроценоза яровой мягкой пшеницы и показатели ее продуктивности. В 2018 году потенциальная урожайность пшеницы в варианте Витаплан, КЖ + Хитозан II превышала контроль на 57,1 % (табл. 1), а в среднем за 2016-2018 годы — на 64,1 % (рис. 1, А).
Мы проанализировали изменения некоторых морфометрических показателей, связаннных с зерновой продуктивностью мягкой пшеницы, при применении биопрепаратов и полифункциональных комплексов по сравнению с контролем (табл. 2, табл. 3, табл. 4).
В варианте опыта Витаплан, КЖ + Хитозан II отмечалось сокращение периода созревания пшеницы по фазам онтогенеза (на 10,2 %), достоверное (р ≤ 0,05) увеличение высоты растений (на 24,3 %), длины корней (на 11,0 %), массы корней (на 50,6 %), площади флагового листа (на 30,9%), числа колосков в колосе (на 6,1 %).
Между потенциальной продуктивностью и продуктивной кустистостью была выявлена сильная корреляционная связь (коэффициент непараметрической корреляции Спирмена r = 0,76; р = 0,029E-9). В нашем опыте наибольшую продуктивную кустистость отмечали при использовании полифункционального комплекса Витаплан, КЖ + Хитозан II (см. рис. 1, Б).
В среднем за период наблюдений (2016-2018 годы) этот показатель вырос на 80,1 % по сравнению с контролем.
В 2018 году отмечали достоверное (р ≤ 0,05) увеличение полевой всхожести пшеницы при применении полифункциональных комплексов Витаплан, КЖ + Хитозан II и Витаплан, КС + Хитозан II, а также биопрепарата Витаплан, КЖ (рис. 2). Максимальное увеличение полевой всхожести зарегистрировали при применении полифункционального комплекса: Витаплан, КЖ + Хитозан II (в 2018 году на 20,8 %, за период 2016-2018 годы — на 19,6 % по сравнению с контролем, р ≤ 0,05). Возможно, это связано с наиболее выраженным по сравнению с другими вариантами опыта снижением развития гельминтоспориозной гнили на пшенице (в 2018 году — на 13,7 %, за 2016-2018 годы — на 24,8 % по сравнению с контролем; р ≤ 0,05).
Потенциальная продуктивность пшеницы в средней степени положительно коррелировала с массой корней (r = 0,46; р = 0,005E-8 (рис. 3, А) и площадью флагового листа (r = 0,40; р = 0,006E-11 (см. рис. 3, Б). Максимальные значения массы корней зарегистрировали при применении полифункционального комплекса Витаплан, КЖ + Хитозан II.
В 2018 году в варианте с комплексом Витаплан, КЖ + Хитозан II отмечали максимальное содержание хлорофилла a во флаговых листьях в фазу цветения пшеницы — 1,32±0,02 мг/г (на 16,7 % больше, чем в контроле, р ≤ 0,05) (табл. 5). Содержание хлорофилла b при этом изменилось незначительно, но на 4,3 % выше по сравнению с контролем (см. табл. 5). Наибольшее изменение соотношения между содержание хлорофиллов a, b и каротиноидов (на 55,2 %) по отношению к контролюмы регистрировали в варианте опыта Витаплан, КЖ + Хитозан II (табл. 6). Некоторые авторы полагают, что соотношение содержания хлорофиллов и каротиноидов может быть одним из показателей устойчивости к внешним неблагоприятным факторам и отражать экологическую пластичность растений (70).
Методом непараметрического корреляционного анализа было установлено, что с увеличением содержания хлорофилла b (Chl b) во флаговых листьях пшеницы отмечалось увеличение числа зерен в колосе (коэффициент корреляции Спирмена r = 0,79; р = 0,006), массы зерен с одного колоса (r = 0,69; р = 0,03); массы колоса с зернами (r = 0,72; р = 0,02) и в целом потенциальной урожайности (r = 0,69; р = 0,03).Зависимость изменения массы колоса с зернами от содержания во флаговых листьях пшеницы хлорофилла b можно описать уравнением регрессии: Mк = 0,26Chl b + 0,81; R2 = 0,45 (рис. 4).
Основными болезнями за период фитосанитарного мониторинга посевов пшеницы (2016-2018 годы) были гельминтоспориозная корневая гниль, вызываемая грибом Bipolaris sorokiniana (Sacc.) Shoemaker, бурая ржавчина (возбудитель Puccinia triticina Eriks.), желтая ржавчина (P. striiformes Westend); мучнистая роса (Blumeria graminis (DC.) Speer.), септориоз, вызываемый Stagonospora nodorum (Berk.) Castellani & E.G. Germano и Zymoseptoria tritici (Desm.) Quaedvlieg & Crous.
Полифункциональные комплексы существенно повлияли на развитие болезней пшеницы. Согласно полученным данным (табл. 7, табл. 8), за период наблюдений комплекс Витаплан, КЖ и Хитозан II достоверно (р ≤ 0,05) снизил развитие корневой гнили (на 24,8 %) и септориоза на предфлаговых листьях (на 22,8 %) по сравнению с контролем. Полифункциональный комплекс Витаплан, КС + Хитозан II достоверно (р ≤ 0,05) уменьшил пораженность пшеницы корневой гнилью (на 11,3 %). Развитие бурой ржавчины снизилось на 8,5 %, число уредопустул — на 81,5 %, площадь уредопустулы — на 43,9 % (р ≤ 0,05). При поражении мучнистой росой число пятен на флаговых листьях уменьшилось на 86,9 %. Симптомов развития мучнистой росы на предфлаговых листьях мы не выявили.
В 2018 году в вариантах с применением полифункциональных комплексов Витаплан, КЖ + Хитозан II и Витаплан, КС + Хитозан II мы отмечали, что интенсивность развития желтой ржавчины на пшенице (R), число пустул (N) и площадь пустул (S) были минимальными (соответственно R = 2,8±1,7 %, N = 106,6±28,5, S = 0,013±0,001 мм2 и R = 2,3±1,3 %, N = 83,0±38,5, S = 0,010±0,0009 мм2) относительно контроля (R = 9,5±3,3 %, N = 221,7±97,8, S = 0,023±0,004 мм2) (табл. 9). Наиболее выраженным оказалось влияние на площадь пустулы, которая уменьшалась по сравнению с контролем.
Растения подвержены негативному воздействию стресс-факторов различной природы в течение всего вегетационного периода, что приводит к снижению продуктивности вследствие ингибирования роста и фотосинтеза. Многие исследователи отмечают существенное снижение фотосинтетической активности растений при поражении фитопатогенными грибами, что связано с уменьшением ассимиляционной поверхности из-за отмирания тканей листа или разрастания мицелия, с разрушением хлоропластов, снижением содержания хлорофилла, нарушением оттока продуктов фотосинтеза из-за повреждения флоэмы (71, 72).
Как показали результаты наших исследований, обработка растений полифункциональными комплексами существенно снизила пораженность пшеницы болезнями, что положительно сказалось на содержании хлорофилла в листьях. Содержание хлорофиллов a и b в листьях пшеницы увеличивалось при снижении степени развития желтой ржавчины в процентах по шкале Маннерса (для хлорофилла а r = -0,66; р = 0,04; для хлорофилла b r = -0,87; р = 0,005), уменьшении числа полос (соответсвтенно r = -0,79; р = 0,02 и r = -0,63; р = 0,04) и числа пустул желтой ржавчины (r = -0,73; р = 0,04 и r = -0,97; р = 0,00007). Зависимость содержания хлорофиллов a и b во флаговых листьях от интенсивности развития желтой ржавчины может быть описана уравнениями: для хлорофилла а — R = -37,03 + 52,72Сhl a, R2 = 0,60, для хлорофилла b — R = 144,52 – 108,23Сhl b + 20,53Сhl b2, R2 = 0,81.
Таким образом, в результате проведенных исследований выявлено статистически достоверное повышение содержания фотосинтетических пигментов (хлорофилла a во флаговых листьях пшеницы) и незначительное увеличение — хлорофилла b при применении биопрепаратов. При использован полифункционального комплекса Витаплан, КЖ + Хитозан II отмечали наибольшую потенциальную продуктивность и самое высокое содержание хлорофиллов a и b в листьях. Выявлена корреляция между увеличением содержания хлорофиллов a и b в листьях и снижением интенсивности развития желтой ржавчины. На основании критерия Спирмена показаны наиболее сильные корреляционные взаимосвязи содержания хлорофилла b с массой колоса, массой зерен с колоса и числом зерен в колосе.
Наблюдаемый положительный эффект биопрепаратов и полифункциональных комплексов может быть связан со способнотью полезные микроорганизмов синтезировать сложные по составу биоактивные комплексы, включающие антибиотики различных химических классов, ферменты, метаболиты с сигнальными и гормональными функциями, фитогормоны, которые могут оказывать значительное влияние на фотосинтетическую функцию, рост и продуктивность растений (9-11). Так, установлено стимулирующее действие синтетического ауксина (индолил-3-масляной кислоты, ИМК) и цитокинина (6-бензиламинопурина, БАП) на накопление биомассы растений, чистую продуктивность фотосинтеза, функционирование фотосинтетического аппарата кукурузы (73). Показано, что гиббереллин усиливает процессы фотосинтетического фосфорилирования, при этом содержание хлорофилла снижается. Таким образом, интенсивность использования хлорофилла в расчете на его количество повышается под воздействием гиббереллина, кроме того, возрастает ассимиляционное число (9). Многие бактерии родов Bacillus, Azospirillium, Pseudomonas, как уже отмечалось, синтезируют ауксины, стимулирующие развитие корневой системы. В совокупности эти процессы повышают устойчивость растений к болезням (74). Многие штаммы бактерий рода Bacillus могут синтезировать гиббереллин (75). Бактерии родов Bacillus, Rhizobium, Arthtrobacter, Azotobacter, Azospirillium, Pseudomonas способны продуцировать цитокинины. При инокуляции цитокинин-продуцирующими штаммами B. subtilis в растениях повышалось содержание хлорофилла и цитокининов, что впоследствии вызывало увеличение биомассы корневой системы и вегетативной части растения. У штаммов Bacillus, Brevibacterium, Azospirillum, Pseudomonas, Lysinibacillus выявлена способность синтезировать абсцизовую кислоту и влиять на ее содержание в растениях, что вызывало оптимизацию эндогенного гормонального баланса (76-79).
Штаммы B. subtilis, составляющие основу Витаплана, продуцируют разнообразные антимикробные метаболиты — липопептиды и полипептиды (80), что в значительной степени определяет фунгицидный эффект биопрепарата в отношении особо опасных фитопатогенных грибов. Биологическая активность хитозана определяется его способностью индуцировать биохимические (сигнальные) пути, приводящие к активации реакций защиты и повышению устойчивости растений к болезням (33-36). Сообщалось, что хитозан стимулирует рост и развитие, повышает урожайность многих сельскохозяйственных растений — кукуруза (81-83), бобовых (84), пшеницы (85), риса (86). Убедительно показано, что обработка хитозаном увеличивает скорость фотосинтеза и содержание хлорофилла в растениях риса и сои (87, 88), кукурузы (38), вигны (89), фасоли (39), томата (40) и пшеницы (37). Возможно, хитозан может быть источником углерода для образования антиоксидантов (90). Хотя точные механизмы влияния хитозана на фотосинтез не установлены, согласно некоторым исследованиям, у кукурузы хитозан и его производные улучшали фотосинтез и флуоресценцию хлорофилла, повышая активность устьиц, скорость транспирации и активность ФС II (38). Есть сообщения о том, что хитозан увеличивает эндогенный уровень цитокининов, стимулирующих синтез хлорофилла (83). Некоторые исследователи связывают увеличение урожайности при применении хитозана с эффектом стимуляции физиологических процессов и последующим активным перемещением фотоассимилятов в потребляющие их ткани. Этот эффект отмечен у кукурузы (38), фасоли (39), сои (87), вигны (89), томата (40) риса (41), огурца (91).
Таким образом, все механизмы действия полезных микроорганизмов в сочетании с биологической активностью хитозана, описанные в приведенных научных публикациях, способны оптимизировать физиологическое состояние растений посредством увеличения скорости фотосинтеза, стимулирования роста и развития, что ведет к повышению стрессо- и болезнеустойчивости и, как следствие, урожайности сельскохозяйственных культур.
Результаты проведенных исследований позволяют предположить, что эффект полифункциональных комплексах Витаплан, КЖ + Хитозан II и Витаплан, КС + Хитозан II обусловлен рассмотренными выше механизмами, что выразилось в высоком защитном действии от грибной инфекции, усилении ростовых процессов, стимуляции репродуктивных свойств растений, увеличении содержания пигментов фотосинтеза и в итоге в увеличении потенциальной продуктивности пшеницы. В разработанных нами композициях сочетание подавляющего воздействия на фитопатогенные микроорганизмы и стимулирующего — на растения обеспечивает более высокую эффективность и надежную защиту по сравнению с биопрепаратами. По нашему мнению, сложные механизмы повышения болезнеустойчивости и обеспечения стабильной продуктивности растений включают адаптивные реакции, в которых задействован фотосинтетический аппарат и вся система фотосинтетических пигментов. Можно полагать, что этот механизм универсален, однако эффективность таких композиций может зависеть от биологических особенностей штаммов микроорганизмов и свойств индукторов, а также от устойчивости сортов культурных растений.
Итак, показано, что полифункциональные комплексы Витаплан, КЖ + Хитозан II и Витаплан, КС + Хитозан II оптимизируют физиологическое состояние растений пшеницы, существенно повышают ее продуктивность и болезнеустойчивость (пораженность растений комплексом болезней снижалась на 17,9 %, р < 0,05). При обработке растений комплексом Витаплан, КЖ + Хитозан II обнаружена самая высокая потенциальная продуктивность пшеницы (0,94±0,02 г/растение), комплексом Витаплан, КС + Хитозан II — наименьшая пораженность растений комплексом болезней. Кроме того, в этих вариантах отмечено наибольшее содержание хлорофиллов a и b в листьях. В варианте Витаплан, КЖ + Хитозан II оно составило для хлорофилла a 1,32±0,02 мг/г, для хлорофилла b — 2,15±0,04 мг/г. При применении комплекса Витаплан, КЖ + Хитозан II также было максимальным соотношение содержания хлорофиллов a и b и пигментов группы каротиноидов, служащее одним из показателей стрессоустойчивости растений. Наиболее сильная корреляция выявлена между содержанием хлорофилла b во флаговых листьях и продуктивностью пшеницы (r = 0,69, р = 0,03), содержанием хлорофилла b во флаговых листьях и числом зерен в колосе (r = 0,79, р = 0,006), массой зерен с колоса и массой колоса (r = 0,69, р = 0,03; r = 0,72, р = 0,02). Обнаружены корреляционные связи между снижением развития желтой ржавчины и увеличением содержания хлорофиллов a и b в листьях (r = -0,66, р= 0,04; r = -0,87, р = 0,005). Проведенные исследования свидетельствуют, что применение полифункциональных композиций на основе отселектированных штаммов бактерий — антагонистов фитопатогенов и индукторов болезнеустойчивости существенно снижает пораженность растений пшеницы комплексом болезней, положительно влияет на содержание фотосинтетических пигментов (хлорофиллы а и b, а также каротиноиды) и показатели продуктивности.
1. Зерновая продуктивность яровой мягкой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 при применении биопрепаратов и полифункциональных комплексов на основе штаммов Bacillus subtilis и хитозана (n = 17, N = 3, M±SEM; опытное поле ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург—Пушкин, 2018 год) |
||||
Вариант опыта |
Продуктивность, г/растение |
t-критерий Стьюдента |
Доверительный интервал |
К контролю, % |
Контроль (вода) |
0,60±0,02 |
|
0,56–0,64 |
|
Витаплан, КЖ |
0,81±0,03 |
1,17 |
0,75–0,87 |
35,3 |
Витаплан, КЖ + Хитозан II |
0,94±0,02 |
2,40 |
0,90–0,98 |
57,1 |
Витаплан, СП |
0,80±0,05 |
0,90 |
0,70–0,90 |
33,7 |
Витаплан, КС + Хитозан II |
0,93±0,03 |
1,97 |
0,88–0,99 |
55,7 |
Примечание. Описание препаратов см. в разделе «Методика». |
||||
2. Морфометрические показатели продуктивности (корневая система) у яровой мягкой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 при применении биопрепаратов и полифункциональных комплексов на основе штаммов Bacillus subtilis и хитозана (n = 51, N = 3, M±SEM; опытное поле ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург—Пушкин, 2016-2018 годы) |
||||||||||||||||
Вариант |
Параметр |
Прохождение |
Высота растения |
Число корней |
Длина корней |
Число узловых корней |
Длина узловых корней |
Масса корней |
||||||||
балл |
к контролю, % |
см |
к контролю, % |
шт. |
к контролю, % |
мм |
к контролю, % |
шт. |
к контролю, % |
мм |
к контролю, % |
г |
к контролю, % |
|||
Контроль (вода) |
M |
51,0 |
|
48,4 |
|
4,9 |
|
73,2 |
|
9,6 |
|
58,0 |
|
0,5 |
|
|
±SEM |
1,5 |
|
2,7 |
|
0,2 |
|
2,6 |
|
0,6 |
|
2,2 |
|
0,0 |
|
||
Витаплан, КЖ |
M |
52,4 |
2,7 |
51,4 |
6,1 |
5,5 |
14,1 |
85,6 |
17,0 |
9,4 |
-2,0 |
59,3 |
2,3 |
0,6 |
11,5 |
|
±SEM |
1,9 |
|
3,2 |
|
0,3 |
|
2,9 |
|
0,6 |
|
2,0 |
|
0,1 |
|
||
Витаплан, КЖ + Хитозан II |
M |
56,2 |
10,2* |
60,2 |
24,3* |
5,3 |
9,5 |
81,2 |
11,0* |
10,1 |
5,9 |
58,6 |
1,1 |
0,7 |
50,6* |
|
±SEM |
1,3 |
|
3,2 |
|
0,3 |
|
2,3 |
|
0,7 |
|
2,4 |
|
0,1 |
|
||
Витаплан, СП |
M |
52,4 |
2,8 |
53,8 |
11,0 |
4,9 |
1,1 |
74,2 |
1,3 |
10,2 |
6,6 |
63,7 |
9,9 |
0,7 |
35,6 |
|
±SEM |
1,9 |
|
3,7 |
|
0,3 |
|
3,2 |
|
0,6 |
|
2,7 |
|
0,2 |
|
||
Витаплан, КС + Хитозан II |
M |
48,9 |
-4,1 |
37,6 |
-22,3 |
4,8 |
-0,8 |
67,3 |
-8,1 |
8,9 |
-7,3 |
48,0 |
-17,2* |
0,3 |
-32,1* |
|
±SEM |
2,0 |
|
3,2 |
|
0,2 |
|
3,5 |
|
0,4 |
|
1,9 |
|
0,0 |
|
||
Примечание. Описание препаратов см. в разделе «Методика». |
||||||||||||||||
3. Морфометрические показатели продуктивности (надземная биомасса) у яровой мягкой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 при применении биопрепаратов и полифункциональных комплексов на основе штаммов Bacillus subtilis и хитозана (n = 51, N = 3, M±SEM; опытное поле ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург—Пушкин, 2016-2018 годы) |
|||||||||||
Вариант опыта |
Параметр |
Площадь флаг-листа |
Площадь предфлаг-листа |
Масса вегетативной части |
Длина колоса |
Число колосков в колосе |
|||||
см2 |
к контролю, % |
см2 |
к контролю, % |
г |
к контролю, % |
мм |
к контролю, % |
шт. |
к контролю, % |
||
Контроль (вода) |
M |
3,8 |
|
4,3 |
|
2,2 |
|
63,2 |
|
13,1 |
|
±SEM |
0,3 |
|
0,3 |
|
0,2 |
|
1,6 |
|
0,2 |
|
|
Витаплан, КЖ |
M |
4,8 |
27,1 |
4,7 |
9,1 |
2,4 |
8,1 |
57,7 |
-8,7 |
13,2 |
1,2 |
±SEM |
0,5 |
|
0,4 |
|
0,2 |
|
1,7 |
|
0,3 |
|
|
Витаплан, КЖ + Хитозан II |
M |
5,0 |
30,9* |
4,5 |
5,1 |
2,5 |
14,1 |
64,6 |
2,1 |
13,8* |
6,1* |
±SEM |
0,3 |
|
0,3 |
|
0,2 |
|
2,4 |
|
0,2 |
|
|
Витаплан, СП |
M |
5,4 |
42,6* |
4,7 |
9,8 |
3,0 |
33,6* |
60,2 |
-4,9 |
13,8 |
6,0 |
±SEM |
0,6 |
|
0,5 |
|
0,3 |
|
2,3 |
|
0,3 |
|
|
Витаплан, КС + Хитозан II |
M |
2,3 |
-40,6* |
2,5 |
-40,8* |
2,0 |
-8,9 |
62,3 |
-1,4 |
11,5 |
-11,6 |
±SEM |
0,1 |
|
0,2 |
|
0,2 |
|
2,9 |
|
0,2 |
|
|
Примечание. Описание препаратов см. в разделе «Методика». |
|||||||||||
4. Морфометрические показатели продуктивности (структура колоса) у яровой мягкой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 при применении биопрепаратов и полифункциональных комплексов на основе штаммов Bacillus subtilis и хитозана (n = 60, N = 3, M±SEM; опытное поле ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург—Пушкин, 2016-2018 годы) |
|||||||||||
Вариант опыта |
Параметр |
Масса колоса |
Число зерен в колосе |
Масса зерен c колоса |
Масса 1000 зерен |
Масса колоса с зернами |
|||||
г |
к контролю, % |
шт. |
к контролю, % |
г |
к контролю, % |
г |
к контролю, % |
г |
к контролю, % |
||
Контроль (вода) |
M |
0,6 |
|
28,9 |
|
0,9 |
|
30,7 |
|
1,1 |
|
±SEM |
0,0 |
|
0,7 |
|
0,0 |
|
0,9 |
|
0,0 |
|
|
Витаплан, КЖ |
M |
0,6 |
-2,6 |
30,1 |
4,3 |
0,9 |
5,8 |
31,1 |
1,3 |
1,3 |
12,5 |
±SEM |
0,0 |
|
1,0 |
|
0,0 |
|
1,1 |
|
0,1 |
|
|
Витаплан, КЖ + Хитозан II |
M |
0,6 |
-5,1 |
28,3 |
-1,8 |
0,8 |
-6,1 |
28,6 |
-6,8 |
1,1 |
-3,1 |
±SEM |
0,0 |
|
0,8 |
|
0,0 |
|
1,0 |
|
0,0 |
|
|
Витаплан, СП |
M |
0,8 |
27,3 |
31,3 |
8,6 |
1,0 |
13,7* |
31,0 |
1,0 |
1,3 |
19,5* |
±SEM |
0,1 |
|
1,0 |
|
0,0 |
|
1,2 |
|
0,0 |
|
|
Витаплан, КС + Хитозан II |
M |
0,5 |
-22,7* |
28,4 |
-1,6 |
0,9 |
4,6 |
33,4 |
8,8* |
1,2 |
7,9 |
±SEM |
0,0 |
|
0,7 |
|
0,0 |
|
0,4 |
|
0,0 |
|
|
Примечание. Описание препаратов см. в разделе «Методика». |
|||||||||||
5. Содержание хлорофиллов a и b во флаговых листьях яровой мягкой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 при применении биопрепаратов и полифункциональных комплексов на основе штаммов Bacillus subtilis и хитозана (n = 17, N = 3, M±SEM; опытное поле ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург—Пушкин, 2018 год) |
|||||||||||
Вариант опыта |
1 |
2 |
3 |
4 |
5 |
2 |
3 |
6 |
7 |
8 |
|
Контроль (вода) |
1,13±0,07 |
|
0,99 |
1,26 |
|
2,06±0,20 |
|
1,67 |
2,45 |
|
0,55 |
Витаплан, КЖ |
1,13±0,10 |
0,06 |
0,94 |
1,33 |
0,69 |
2,07±0,12 |
0,05 |
1,84 |
2,30 |
0,58 |
0,55 |
Витаплан, КЖ + Хитозан II |
1,32±0,02 |
2,55* |
1,27 |
1,36 |
16,72 |
2,15±0,04 |
0,43 |
2,07 |
2,23 |
4,30 |
0,61 |
Витаплан, СП |
1,05±0,20 |
-0,36 |
0,67 |
1,44 |
-6,62 |
1,99±0,40 |
-0,15 |
1,20 |
2,78 |
-3,24 |
0,53 |
Витаплан, КС + Хитозан II |
0,96±0,17 |
-0,90 |
0,64 |
1,29 |
-14,38 |
1,63±0,27 |
-1,26 |
1,10 |
2,16 |
-20,70 |
0,59 |
Примечание. 1 — содержание хлорофилла a, мг/г; 2 — критерий Стьюдента; 3 — доверительные интервалы для 5 % уровня значимости; 4 — изменениt содержания хлорофилла a относительно контроля, %; 5 — содержание хлорофилла b, мг/г; 6 — изменениt содержания хлорофилла b относительно контроля, %; 7 — соотношение хлорофиллов a и b; 8 — изменение соотношения хлорофиллов a и b относительно контроля, %. Описание препаратов см. в разделе «Методика». |
|||||||||||
6. Соотношение содержания хлорофиллов a + b и каротиноидов во флаговых листьях яровой мягкой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 при применении биопрепаратов и полифункциональных комплексов на основе штаммов Bacillus subtilis и хитозана (n = 17, N = 3, M±SEM; опытное поле ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург—Пушкин, 2018 год) |
||||
Вариант опыта |
Соотношение |
Критерий Стьюдента |
Доверительный |
Изменение соотношения хлорофиллов a + b и каротиноидов к контролю, % |
Контроль (вода) |
7,60±2,30 |
|
3,09–12,11 |
|
Витаплан, КЖ |
10,80±4,20 |
0,67 |
2,57–19,03 |
42,09 |
Витаплан, КЖ + Хитозан II |
11,80±2,23 |
1,31 |
7,42–16,17 |
55,22 |
Витаплан, СП |
6,92±1,62 |
-0,24 |
3,74–10,10 |
-8,92 |
Витаплан, КС + Хитозан II |
8,26±0,87 |
0,27 |
6,56–9,97 |
8,73 |
Примечание. Описание препаратов см. в разделе «Методика». |
||||
7. Развитие бурой ржавчины, мучнистой росы и септориоза на флаговых листьях у яровой мягкой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 при применении биопрепаратов и полифункциональных комплексов на основе штаммов Bacillus subtilis и хитозана (n = 60, N = 3, M±SEM; опытное поле ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург—Пушкин, 2016-2018 годы) |
|||||||||||||||
Вариант опыта |
Параметр |
Бурая ржавчина |
Мучнистая роса |
Септориоза |
|||||||||||
развитие |
число пустул |
площадь пустулы |
развитие |
число пятен |
площадь пятен |
||||||||||
% |
к контролю, % |
шт. |
к контролю, % |
мм2 |
к контролю, % |
% |
к контролю, % |
шт. |
к контролю, % |
мм2 |
к контролю, % |
% |
к контролю, % |
||
Контроль (вода) |
M |
12,5 |
|
81,6 |
|
0,08934 |
|
5,8 |
|
7,6 |
|
3,6 |
|
17,5 |
|
±SEM |
3,8 |
|
24,6 |
|
0,00880 |
|
1,9 |
|
2,3 |
|
1,0 |
|
2,5 |
|
|
Витаплан, КЖ |
M |
7,6 |
-4,9 |
133,2 |
63,2 |
0,04766 |
-46,7* |
1,8 |
-4,0 |
2,8 |
-63,3 |
1,3 |
-63,0 |
7,5 |
-10,0 |
±SEM |
1,8 |
|
49,0 |
|
0,00524 |
|
0,8 |
|
1,4 |
|
0,4 |
|
2,5 |
|
|
Витаплан, КЖ + Хитозан II |
M |
4,8 |
-7,7 |
42,2 |
-48,3 |
0,07286 |
-18,5 |
3,8 |
-2,0 |
5,5 |
-27,9 |
2,5 |
-29,2 |
2,0 |
-15,5 |
±SEM |
1,3 |
|
22,6 |
|
0,01149 |
|
1,4 |
|
1,7 |
|
0,4 |
|
1,0 |
|
|
Витаплан, СП |
M |
10,2 |
-2,4 |
52,2 |
-36,0 |
0,10333 |
15,7 |
2,9 |
-2,9 |
3,1 |
-59,0 |
4,3 |
21,7 |
0,0 |
-17,5* |
±SEM |
5,0 |
|
33,2 |
|
0,02187 |
|
1,1 |
|
1,0 |
|
1,3 |
|
0,0 |
|
|
Витаплан, КС + |
M |
4,1 |
-8,5* |
15,1 |
-81,5* |
0,05016 |
-43,9* |
1,0 |
-4,8* |
1,0 |
-86,9* |
5,5 |
53,9 |
0,0 |
-17,5* |
±SEM |
1,2 |
|
5,0 |
|
0,00688 |
|
0,2 |
|
0,1 |
|
|
|
0,0 |
|
|
Примечание. Описание препаратов см. в разделе «Методика». |
|||||||||||||||
8. Развитие мучнистой росы и септориоза на предфлаговых листьях и корневой гнили у яровой мягкой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 при применении биопрепаратов и полифункциональных комплексов на основе штаммов Bacillus subtilis и хитозана (n = 60, N = 3, M±SEM; опытное поле ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург—Пушкин, 2016-2018 годы) |
|||||||||||
Вариант опыта |
Параметр |
Мучнистая роса |
Септориоз |
Корневая гниль |
|||||||
развитие |
число пятен |
площадь пятен |
|||||||||
% |
к контролю, % |
шт. |
к контролю, % |
мм2 |
к контролю, % |
% |
к контролю, % |
% |
к контролю, % |
||
Контроль (вода) |
M |
22,0 |
|
23,1 |
|
3,3 |
|
31,5 |
|
39,7 |
|
±SEM |
5,0 |
|
4,7 |
|
0,5 |
|
8,4 |
|
2,9 |
|
|
Витаплан, КЖ |
M |
24,2 |
2,2 |
22,3 |
-3,3 |
2,9 |
-11,8 |
21,9 |
-9,6 |
19,7 |
-20,0* |
±SEM |
5,8 |
|
3,8 |
|
0,4 |
|
7,8 |
|
5,5 |
|
|
Витаплан, КЖ + Хитозан II |
M |
18,6 |
-3,4 |
20,6 |
-10,7 |
2,8 |
-14,4 |
8,8 |
-22,8* |
14,9 |
-24,8* |
±SEM |
4,3 |
|
3,9 |
|
0,3 |
|
2,7 |
|
6,5 |
|
|
Витаплан, СП |
M |
30,5 |
8,5 |
43,7 |
88,9 |
4,8 |
45,4* |
6,3 |
-25,2* |
33,8 |
-5,9 |
±SEM |
5,9 |
|
13,4 |
|
0,5 |
|
3,1 |
|
6,2 |
|
|
Витаплан, КС + Хитозан II |
M |
0,0 |
-22,0* |
0,0 |
-100,0* |
0,0 |
-100,0* |
15,9 |
-15,6 |
28,3 |
-11,3* |
±SEM |
0,0 |
|
0,0 |
|
0,0 |
|
8,0 |
|
4,7 |
|
|
Примечание. Описание препаратов см. в разделе «Методика». |
|||||||||||
9. Развитие желтой ржавчины на флаговых листьях у яровой мягкой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 при применении биопрепаратов и полифункциональных комплексов на основе штаммов Bacillus subtilis и хитозана (n = 60, N = 3, M±SEM; опытное поле ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург—Пушкин, 2018 год) |
|||||||||||||
Вариант опыта |
Параметр |
Развитие |
Число полос |
Длина полосы |
Пустулы |
Площадь пустулы |
|||||||
число в полосе |
суммарное число |
||||||||||||
% |
к контролю, % |
шт. |
к контролю, % |
мм |
к контролю, % |
шт. |
к контролю, % |
шт. |
к контролю, % |
мм2 |
к контролю, % |
||
Контроль (вода) |
M |
9,5 |
|
2,2 |
|
40,9 |
|
102,7 |
|
221,7 |
|
0,02312 |
|
±SEM |
3,3 |
|
0,5 |
|
7,6 |
|
26,4 |
|
97,8 |
|
0,00391 |
|
|
Витаплан, КЖ |
M |
5,7 |
-3,8 |
2,6 |
18,2 |
31,9 |
-22,0 |
66,0 |
-35,7 |
168,8 |
-23,9 |
0,02329 |
0,7 |
±SEM |
1,4 |
|
0,5 |
|
2,6 |
|
6,0 |
|
45,4 |
|
0,00183 |
|
|
Витаплан, КЖ + Хитозан II |
M |
2,8 |
-6,7 |
1,4 |
-36,4 |
28,5 |
-30,3 |
58,1 |
-43,4 |
106,6 |
-51,9 |
0,01327 |
-42,6* |
±SEM |
1,8 |
|
0,3 |
|
3,4 |
|
6,8 |
|
28,5 |
|
0,00143 |
|
|
Витаплан, СП |
M |
7,7 |
-1,8 |
3,3 |
50,0 |
18,2 |
-55,5* |
41,9 |
-59,2* |
148,3 |
-33,1 |
0,01653 |
-28,5 |
±SEM |
4,1 |
|
1,1 |
|
1,7 |
|
6,4 |
|
77,6 |
|
0,00341 |
|
|
Витаплан, КС + Хитозан II |
M |
2,3 |
-7,2 |
1,3 |
-40,9 |
26,3 |
-35,7 |
62,3 |
-39,3 |
83,0 |
-62,6 |
0,01022 |
-55,8* |
±SEM |
1,3 |
|
0,3 |
|
8,0 |
|
32,8 |
|
38,6 |
|
0,00086 |
|
|
Примечание. Описание препаратов см. в разделе «Методика». |
|||||||||||||
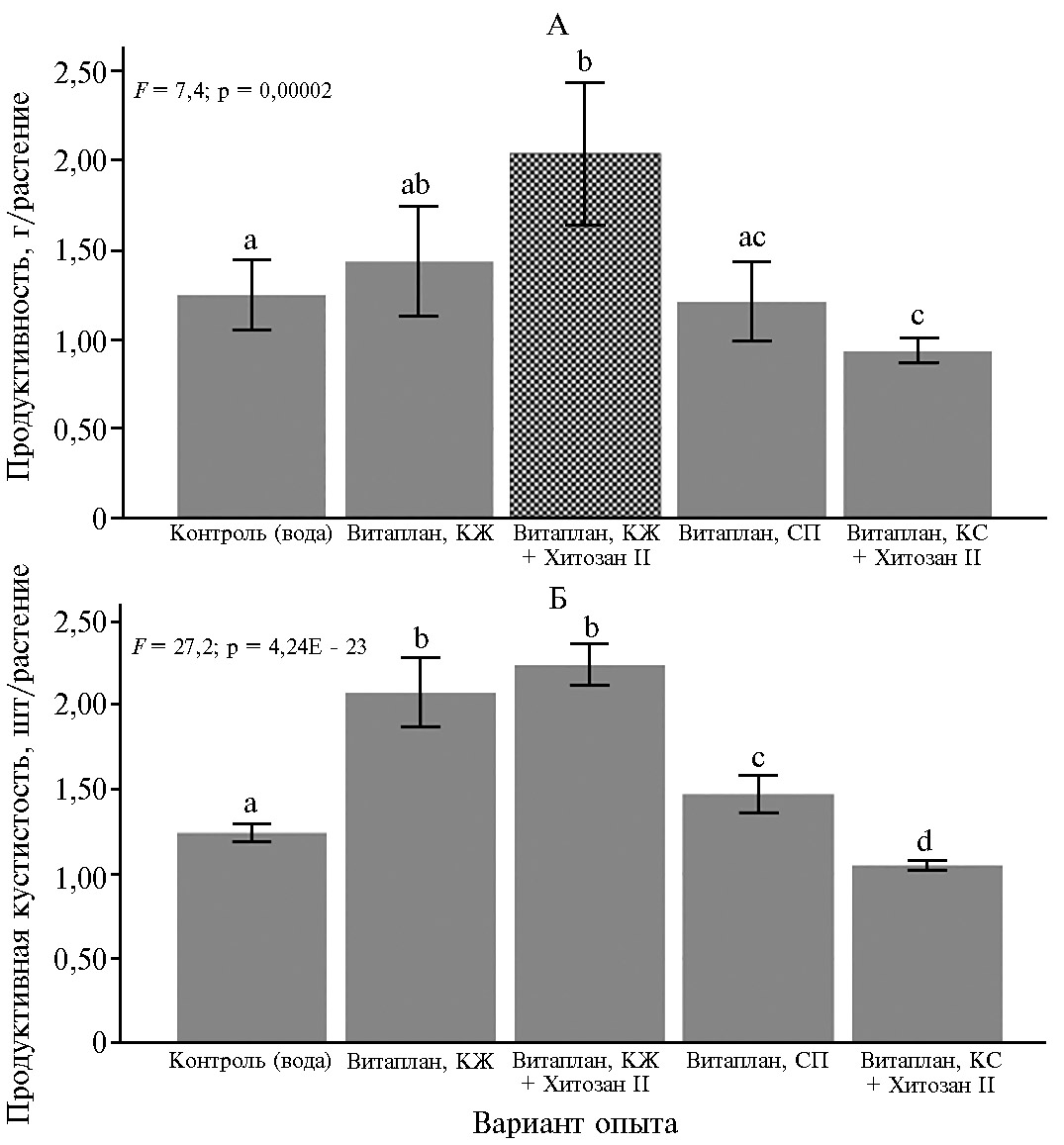
Рис. 1. Потенциальная зерновая продуктивность (А) и продуктивная кустистость (Б) у яровой мягкой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 при применении биопрепаратов и полифункциональных комплексов на основе штаммов Bacillus subtilis и хитозана (n = 51, N = 3, M±SEM; опытное поле ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург—Пушкин, 2016-2018 годы). Описание препаратов см. в разделе «Методика». Одинаковыми буквами отмечены значения, не различающиеся с контролем и между собой по критерию Шеффе при р < 0,05; F — критерий Фишера.
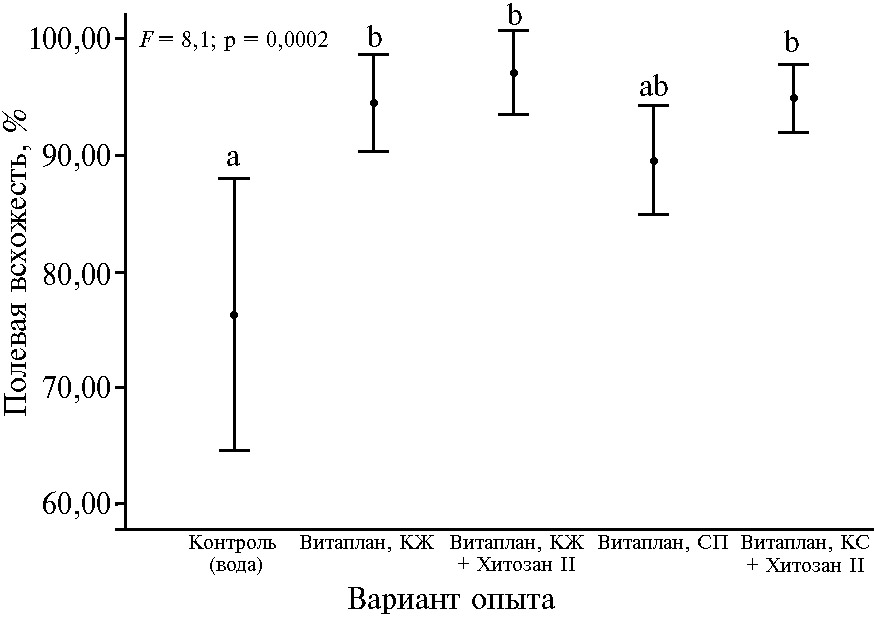
Рис. 2. Полевая всхожесть у яровой мягкой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 при применении биопрепаратов и полифункциональных комплексов на основе штаммов Bacillus subtilis и хитозана (n = 1050, N = 3, M±t05SEM; опытное поле ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург—Пушкин, 2016-2018 годы). Описание препаратов см. в разделе «Методика». Одинаковыми буквами отмечены значения, не различающиеся с контролем и между собой по критерию Шеффе при р < 0,05; F — критерий Фишера.
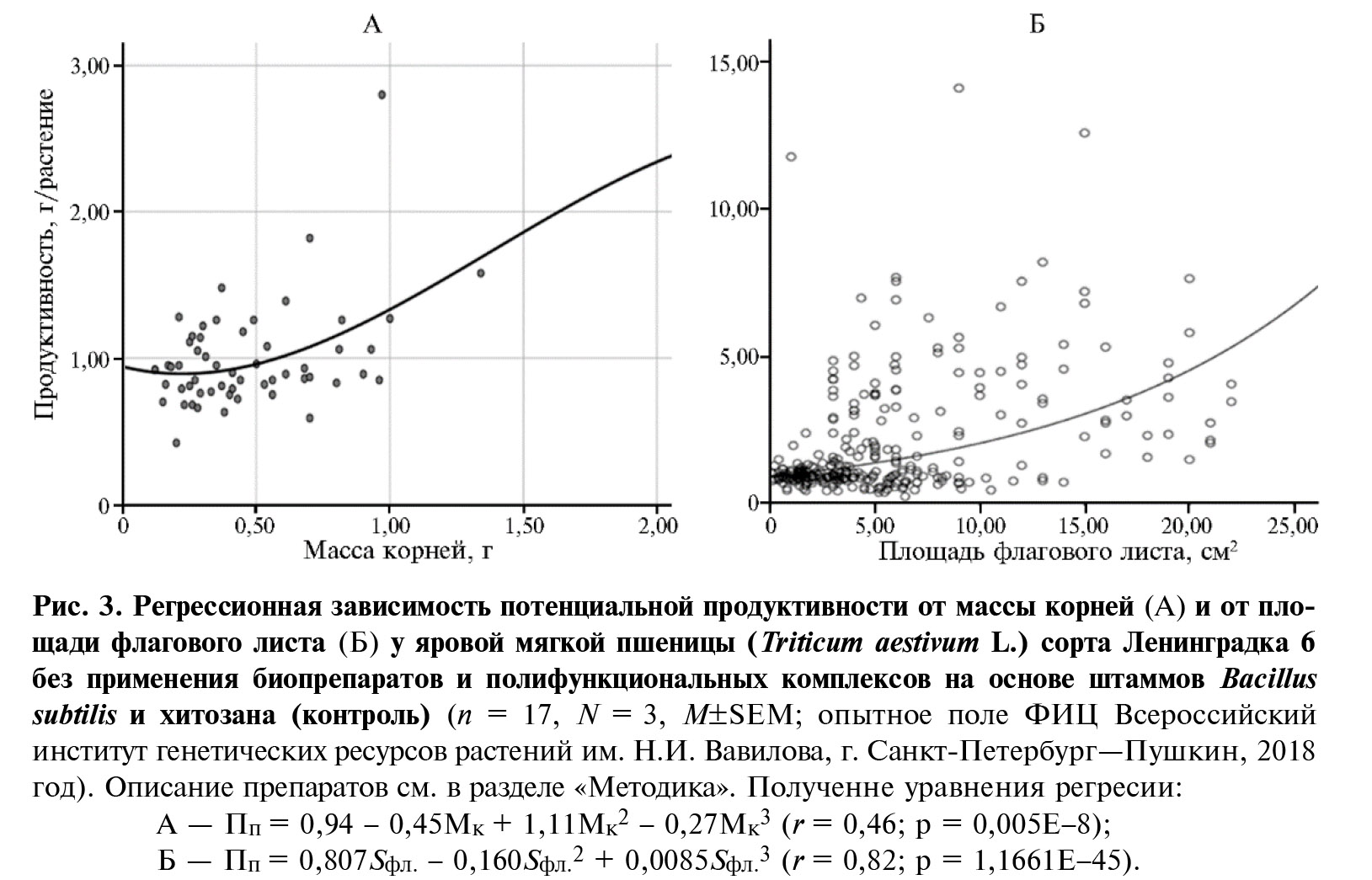
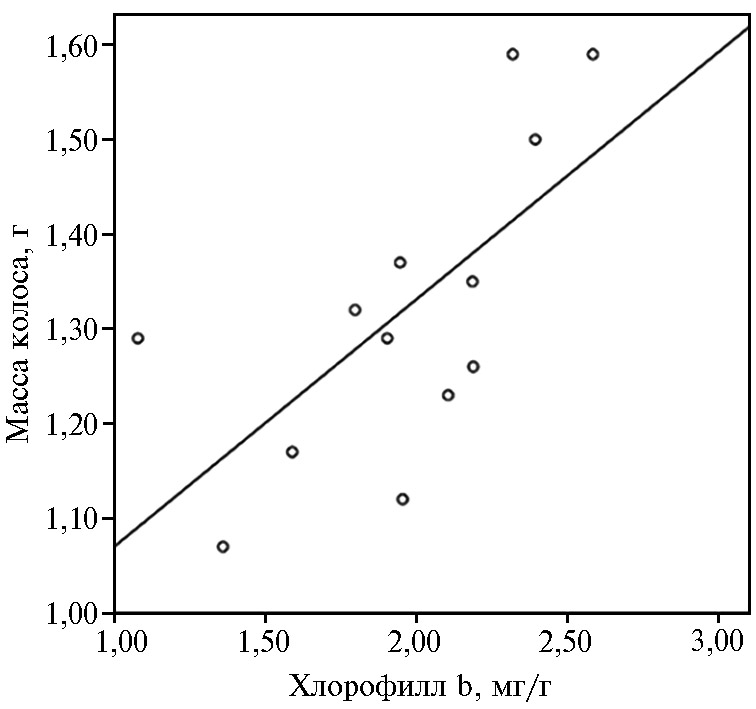
Рис. 4. Зависимость изменения массы колоса с зерном от содержания хлорофилла b во флаговых листьях у яровой мягкой пшеницы (Triticum aestivum L.) сорта Ленинградка 6 без применения биопрепаратов и полифункциональных комплексов на основе штаммов Bacillus subtilis и хитозана (контроль) (n = 13, N = 3; опытное поле ФИЦ Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова, г. Санкт-Петербург—Пушкин, 2018 год).
ЛИТЕРАТУРА
- Новикова И.И., Титова Ю.А., Бойкова И.В., Зейрук В.Н., Краснобаева И.Л., Серова Т.А. Биологическое обоснование оптимизации препаративных форм биопрепаратов на основе микробов-антагонистов для контроля популяций фитопатогенных грибов и бактерий — возбудителей болезней растений. Вестник защиты растений, 2017, 3(93): 16-23.
- Tamehiro N., Okamoto-Hosoya Y., Okamoto S., Ubukata M., Hamada M., Naganawa H., Ochi K. Bacilysocin, a novel phospholipid antibiotic produced by Bacillus subtilis 168. Antimicrobial Agents and Chemotherapy Journal, 2002, 46(2): 315-320 (doi: 10.1128/aac.46.2.315-320.2002).
- Tojo S., Tanaka Y., Ochi K. Activation of antibiotic production in Bacillus spp. by cumulative drug resistance mutations. Antimicrobial Agents and Chemotherapy Journal, 2015, 59(12): 7799-7804 (doi: 10.1128/AAC.01932-15).
- Hamdache A., Lamarti A., Aleu J., Collado I.G. Non-peptide metabolites from the genus Bacillus. Journal of Natural Products, 2011, 74(4): 893-899 (doi: 10.1021/np100853e).
- Quardros C.P., Teixeira Duarte M.C., Pastore G.M. Biological activities of a mixture of biosurfactant from Bacillus subtilis and alkaline lipase from Fusarium oxysporum. Brazilian Journal of Microbiology, 2011, 42: 354-361 (doi: 10.1590/s1517-83822011000100045).
- Актуганов Г.Э., Галимзянова Н.Ф. Мелентьев А.И., Кузьмина Л.Ю. Внеклеточные гидролазы штамма Basillus sp. 739 и их участие в лизисе клеточных стенок микромицетов. Микробиология, 2007, 76: 471-479.
- Kloepper J.W., Gutierrez-Estrada A., McInroy J.A.Photoperiod regulates elicitation of growth promotion but not induced resistance by plant growth-promoting rhizobacteria. Canadian Journal of Microbiology, 2009, 53(2): 159-167 (doi: 10.1139/w06-114).
- Ohkama-Ohtsu N., Wasaki J. Recent progress in plant nutrition research: cross-talk between nutrients, plant physiology and soil microorganisms. Plant and Cell Physiology, 2010, 51(8): 1255-1264 (doi: 10.1093/pcp/pcq095).
- Dodd I.C., Zinovkina N.Y., Safronova V.I., Belimov A.A. Rhizobacterial mediation of plant hormone status. Annals of Applied Biology, 2010, 157: 361-379 (doi: 10.1111/j1744-7348.2010.00439.x).
- Forchetti G., Masciarelli O., Alemano S., Alvarez D., Abdala G. Endophytic bacteria in sunflower (Helianthus annuus L.): isolation, characterization, and production of jasmonates and abscisic acid in culture medium. Applied Microbiology and Biotechnology, 2007, 76(5): 1145-1152 (doi: 10.1007/s00253-007-1077-7).
- Kudoyarova G.R., Melentiev A.I., Martynenko E.V., Timergalina L.N., Arkhipova T.N., Shendel G.V., Kuz’mina L.Y., Dodd I.C., Veselov S.Y. Cytokinin producing bacteria stimulate amino acid deposition by wheat roots. Plant Physiology and Biochemistry, 2014, 83: 285-291 (doi: 10.1016/j.plaphy.2014.08.015).
- Sivasakthi S., Kanchana D., Usharani G., Saranraj P. Production of plant growth promoting substance by Pseudomonas fluorescens and Bacillus subtilis isolates from paddy rhizosphere soil of Cuddalore district, Tamil Nadu, India. International Journal of Microbiological Research, 2013, 4(3): 227-233 (doi: 10.5829/idosi.ijmr.2013.4.3.75171).
- Kumar P., Dubey R.C., Maheshwari D.K. Bacillus strains isolated from rhizosphere showed plant growth promoting and antagonistic activity against phytopathogens. Microbiological Research, 2012, 167: 493-499 (doi: 10.1016/j.micres.2012.05.002).
- Максимов И.В., Веселова С.В., Нужная Т.В., Сарварова Е.Р., Хайруллин Р.М. Стимулирующие рост растений бактерии в регуляции устойчивости растений к стрессовым факторам. Физиология растений, 2015, 62(6): 763-775 (doi: 10.7868/S0015330315060111).
- Kolesnikov L.E., Novikova I.I., Surin V.G., Popova E.V., Priyatkin N.S., Kolesnikova Yu.R. Evaluation of the effectiveness of the combined use of chitosan and antagonist microbes in protection of spring soft wheat from diseases using spectrometric analysis. Applied Biochemistry and Microbiology, 2018, 54(5): 546-552 (doi: 10.1134/S0003683818050083).
- El Amerany F., Meddich A., Wahbi S., Porzel A, Taourirte M., Rhazi M., Hause B. Foliar application of chitosan increases tomato growth and influences mycorrhization and expression of endochitinase-encoding genes. International Journal of Molecular Sciences, 2020, 21(2): 535 (doi: 10.3390/ijms21020535).
- Yang H., Zhang Y., Zhou F., Guo J., Tang J., Han Y., Fu C. Preparation, bioactivities and applications in food industry of chitosan-based maillard products: A review. Molecules, 2021, 26(1): 166 (doi: 10.3390/molecules26010166).
- Тютерев С.Л. Природные и синтетические индукторы устойчивости растений к болезням. СПб, 2014.
- Badawy M.E.I., Rabea E.I. A biopolymer chitosan and its derivatives as promising antimicrobial agents against plant pathogens and their applications in crop protection. International Journal of Carbohydrate Chemistry, 2011, 2011: Article ID 460381 (doi: 10.1155/2011/460381).
- Abdul Malik N.A., Kumar I.S., Nadarajah K. Elicitor and receptor molecules: orchestrators of plant defense and immunity, review. International Journal of Molecular Sciences, 2020, 21(3): 963 (doi: 10.3390/ijms21030963).
- Lopez-Moya F., Suarez-Fernandez M., Lopez-Llorca L.V. Molecular mechanisms of chitosan interactions with fungi and plants. International Journal of Molecular Sciences, 2019,20(2): 332(doi: 10.3390/ijms20020332).
- Meenakshi T., Baldev S.S., Role of elisitors in inducing resistanse in plant against pathogen infection: a review. ISRN Biochemistry, 2013, 2013: Article ID 762 (doi: 10.1155/2013/762412).
- Iriti M., Faoro F. Chitosan as a MAMP, searching for a PRR. Plant Signaling and Behavior, 2009, 4(1): 66-68 (doi: 10.4161/psb.4.1.7408).
- Deepmala K., Hemantaranjan A., Bharti S., Nishant Bhanu A. A future perspective in crop protection: chitosan and its oligosaccharides. Advances in Plants & Agriculture Research, 2014, 1(1): 23-30 (doi: 10.15406/apar.2014.01.00006).
- Bigeard J., Colcombet J., Hirt H. Signaling mechanisms in pattern-triggered immunity (PTI). Molecular Plant, 2015, 8(4): 521-539 (doi: 10.1016/j.molp.2014.12.022).
- Silva W.B., Silva G.M.C., Santana D.B., Salvador A.R., Medeiros D.B., Belghith I., Misobutsi G.P. Chitosan delays ripening and ROS production in guava (Psidium guajava L.) fruit. Food Chemistry, 2018, 242: 232-238 (doi: 10.1016/j.foodchem.2017.09.052).
- Gai Q.Y., Jiao J., Wang X., Liu J., Wang Z., Fu Y.J. Chitosan promoting formononetin and calycosin accumulation in Astragalus membranaceus hairy root cultures via mitogen-activated protein kinase signaling cascades. Scientific Reports, 2019, 9: 10367 (doi: 10.1038/s41598-019-46820-6).
- Li Y., Zhang Q., Ou L., Ji D., Liu T., Lan R., Li X., Jin L. Response to the cold stress signaling of the tea plant (Camellia sinensis) elicited by chitosan oligosaccharide. Agronomy, 2020, 10: 915 (doi: 10.3390/agronomy10060915).
- Dubin A., Likhanov A., Klyachenko O., Subin A., Kluvadenko A. Effect of chitosan formulations of different biological origin on tobacco (Nicotiana tabacum L.) PR-genes expression. Journal of Microbiology, Biotechnology and Food Sciences, 2021, 9(6): 1141-1144 (doi: 10.15414/jmbfs.2020.9.6.1141-1144).
- Chun S.C., Chandrasekaran M. Chitosan and chitosan nanoparticles induced expression of pathogenesis-related proteins genes enhances biotic stress tolerance in tomato. International Journal of Biological Macromolecules, 2019, 125: 948-954 (doi: 10.1016/j.ijbiomac.2018.12.167).
- Liu J., Zhang X., Kennedy J., Jiang M., Cai Q., Wu X. Chitosan induces resistance to tuber rot in stored potato caused by Alternaria tenuissima. International Journal of Biological Macromolecules, 2019, 140: 851-857 (doi: 10.1016/j.ijbiomac.2019.08.227).
- Jiang X., Lin H., Lin M., Chen Y., Wang H., Lin Y., Lin Y. A novel chitosan formulation treatment induces disease resistance of harvested litchi fruit to Peronophythora litchii in association with ROS metabolism. Food Chemistry, 2018, 266: 299-308 (doi: 10.1016/j.foodchem.2018.06.010).
- Xing K., Zhu X., Peng X., Qin S. Chitosan antimicrobial and eliciting properties for pest control in agriculture: a review. Agronomy for Sustainable Development, 2015, 35(2): 569-588 (doi: 10.1007/s13593-014-0252-3).
- El Hadrami A., Adam L.R., El Hardrami I., Daayf F. Chitosan in plant protection. Marine Drugs, 2010, 8(4): 968-987 (doi: 10.3390/md8040968).
- Yin H., Du Y., Dong Z. Chitin oligosaccharide and chitosan oligosaccharide: two similar but different plant elicitors. Frontiers in Plant Science, 2016, 7: 522 (doi: 10.3389/fpls.2016.00522).
- Torres-Rodriguez J.A., Reyes-Perez J.J., Castellanos T., Angulo C., Quinones-Aguilar E.E., Hernandez-Montiel L.G. A biopolymer with antimicrobial properties and plant resistance inducer against phytopathogens: chitosan. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 2021, 49(1): 12231 (doi: 10.15835/nbha49112231).
- Zhang Z., Li K., Liu S., Xing R., Yu H., Chen X., Li P. Size effects of chitooligomers on the growth and photosynthetic characteristics of wheat seedlings. Carbohydrate Polymers, 2016, 138: 27-33 (doi: 10.1016/j.carbpol.2015.11.050).
- dos Reis C.O., Magalhaes P.C., Avila R.G., Almeida L.G., Rabelo M., Carvalho D.T., Cabral D.F., Karam D., de Sousa T.C. Action of N-succinyl and N,O-dicarboxymethyl chitosan derivatives on chlorophyll photosynthesis and fluorescence in drought-sensitive maize. Journal of Plant Growth Regulation, 2019, 38: 619-630 (doi: 10.1007/s00344-018-9877-9).
- Mondal M.M.A., Malek M.A., Puteh A.B., Ismail M.R. Foliar application of chitosan on growth and yield attributes of mungbean (Vigna radiata (L.) Wilczek). Bangladesh Journal of Botany. 2013, 42(1): 179-183 (doi: 10.3329/bjb.v42i1.15910).
- Khan H., Basit A., Alam M., Ahmad I., Ullah Iz., Alam N., Ullah In., Khalid M.A., Shair M., ul Ain N. Efficacy of chitosan on performance of tomato (Lycopersicon esculentum L.) plant under water stress condition. Pakistan Journal of Agricultural Research, 2020, 33(1): 27-41 (doi: 10.17582/journal.pjar/2020/33.1.27.41).
- Rutairat P., Chonlada D.T. Effect of chitosan on physiology, photosynthesis and biomass of rice (Oryza sativa L.) under elevated ozone. Australian Journal of Crop Science, 2017, 11(5): 624-630 (doi: 10.21475/ajcs.17.11.05.p578).
- Zeng D., Luo X. Physiological effects of chitosan coating on wheat growth and activities of protective enzyme with drought tolerance. Open Journal of Soil Science, 2012, 2: 282-288 (doi: 10.4236/ojss.2012.23034).
- Дуденко Н.В., Андрианова Ю.E., Максютова Н.Н. Формирование хлорофилльного фотосинтетического потенциала пшеницы в сухой и влажный годы. Физиология растений, 2002, 49(5): 684-687.
- Прядкина Г.А. Пигменты, эффективность фотосинтеза и продуктивность пшеницы. Сортоизучение и охрана прав на сорта растений, 2018, 14(1): 97-108 (doi: 10.21498/2518-1017.14.1.2018.126524).
- Furbank R.T., Quick W.P., Sirault X.R.R. Improving photosynthesis and yield potential in cereal crops by targeted genetic manipulation: prospects, progress and challenges. Field Crops Research, 2015, 182: 19-29 (doi: 10.1016/j.fcr.2015.04.009).
- Gu J., Yin X., Stomph T.J., Struik P.C. Can exploiting natural genetic variation in leaf photosynthesis contribute to increasing rice productivity? A simulation analysis. Plant Cell and Environment, 2014, 37(1): 22-34 (doi: 10.1111/pce.12173).
- Voitsekhovskaja O.V., Tyutereva E.V. Chlorophyll b in angiosperms: functions in photosynthesis, signaling and ontogenetic regulation. Journal of Plant Physiology, 2015, 189: 51-64 (doi: 10.1016/j.jplph.2015.09.013).
- Тютерева Е.В., Дмитриева В.А., Войцеховская О.В. Хлорофилл b как источник сигналов, регулирующих развитие и продуктивность растений (обзор). Сельскохозяйственная биология, 2017, 52(5): 843-855 (doi: 10.15389/agrobiology.2017.5.843rus).
- Тютерева Е.В., Иванова А.Н., Войцеховская О.В. К вопросу о роли хлорофилла b в онтогенетических адаптациях растений. Успехи современной биологии, 2014, 134: 249-256.
- Mamadyusupova M.G., Saboiev I.A., Nigmonov M., Nasyrova F.Yu., Aliev K. The influence of stress factors on the content of photosynthetic pigments and biometric indicators in wheat varieties and their wild relatives. News of the Academy of Sciences of the Republic Tajikistan, 2012, 3(180): 35-42.
- Sharma A., Kumar V., Shahzad B., Ramakrishnan M., Shreeya Bali A., Yadav P., Rehman A., Daman R., Renu P., Gagan Preet B., Sidhu S., Handa N., Khanna K., Kohli S., Yuan H., Zheng Bi., Kapoor D., Bakshi P., Khan E.A., Kumar Thukral A. Photosynthetic response of plants under different abiotic stresses: a review. Journal of Plant Growth Regulation,2020, 39(2): 509-531 (doi: 10.1007/s00344-019-10018-x).
- Muhammad I., Shalmani A., Ali M., Yang Q.-H., Ahmad H., Li F.B. Mechanisms regulating the dynamics of photosynthesis under abiotic stresses. Frontiers in Plant Science, 2021, 11: 615942 (doi: 10.3389/fpls.2020.615942).
- Абдуллаев Х.А., Гиясидинов Б.Б., Солиева Б.А., Миракилов Х.М. Парадокс: фотосинтез хлопчатника за сто лет селекции новых сортов как консервативный признак существенно не изменился. Тезисы Всероссийской научной конференции с международным участием и Школы для молодых ученых «Растения в условиях глобальных и локальных природно-климатических и антропогенных воздействий» (21-26 сентября 2015 г.). Петрозаводск, 2015: 19.
- Андрианова Ю.Е., Тарчевский И.А. Хлорофилл и продуктивность растений. М., 2000.
- Юрин В.М. Физиология растений. Минск, 2010.
- Мокроносов А.Т., Гавриленко В.Ф., Жигалова Т.В. Фотосинтез. Физиолого-экологические и биохимические аспекты. М., 2006.
- Амунова О.С., Лисицын Е.М. Влияние различных условий увлажнения на пигментный комплекс листьев сортов мягкой яровой пшеницы разных групп спелости. Самарский научный вестник, 2019, 8(3): 19-25 (doi: 10.24411/2309-4370-2019-13102).
- Sui N., Li M., Meng Q.-W., Tian J., Zhao S. Photosynthetic characteristics of a super high yield cultivar of winter wheat during late grown period. Agricultural Sciences in China, 2010, 9(3): 346-354 (doi: 10.1016/S1671-2927(09)60103-6).
- Моргун В.В., Прядкина Г.А. Эффективность фотосинтеза и перспективы повышения продуктивности озимой пшеницы. Физиология растений и генетика, 2014, 46(4): 279-301.
- Мартиросян Л.Ю., Кособрюхов А.А., Мартиросян В.В., Мартиросян Ю.Ц. О влиянии различных источников света на фотосинтетические параметры продукционного процесса у Cucumis sativus L. (гибрид Тристан F1) в условиях аэропонного фитотрона. Сельскохозяйственная биология, 2021, 56(5): 934-947 (doi: 10.15389/agrobiology.2021.5.934rus).
- Корнилина В.В. Влияние ложного осинового трутовика (Phellinus tremulae Bond et Boriss) на содержание пигментов в листьях осины в лесах Ульяновской области. Фундаментальные исследования, 2012, 9(3): 568-572.
- Muzzarelli R.A.A. Chitin. Pergamon Press, Oxford, 1977.
- Колесников Л.Е., Попова Э.В., Новикова И.И., Прияткин Н.С., Архипов М.В., Колесникова Ю.Р., Потрахов Н.Н., Van Duijn B., Гусаренко А.С. Совместное использование штаммов микроорганизмов и хитозановых комплексов для повышения урожайности пшеницы (Triticuma estivum L.). Сельскохозяйственная биология, 2019, 54(5): 1024-1040 (doi: 10.15389/agrobiology.2019.5.1024rus).
- Захарова Н.Н., Захаров Н.Г. Посевные качества и полевая всхожесть семян яровой мягкой пшеницы. Вестник Ульяновской государственной сельскохозяйственной академии, 2016, 4(36): 17-23 (doi: 10.18286/1816-4501-2016-4-17-23).
- Zadoks J.C., Chang T.T., Konzak C.F. A decimal code for the growth stages of cereals. Weed Research, 1974, 14: 415-421 (doi: 10.1111/j.1365-3180.1974.tb01084.x)
- Дмитриев Н.Н., Хуснидинов Ш.К. Методика ускоренного определения площади листовой поверхности сельскохозяйственных культур с помощью компьютерной технологии. Вестник КрасГАУ, 2016, 7(118): 88-93.
- Попов Ю.В. Метод оценки развития корневых гнилей зерновых культур. Защита и карантин растений, 2011, 8: 45-47.
- Новикова И.И., Попова Э.В., Коваленко Н.М., Краснобаева И.Л. Факторы повышения индуцирующей активности двух штаммов Bacillussubtilis в защите пшеницы от возбудителей темно-бурой пятнистости (Bipolarissorokiniana) и бурой ржавчины (Pucciniatriticina). Вестник защиты растений, 2022, 105(3): 122-134 (doi: 10.31993/2308-6459-2022-105-3-15320).
- Ермаков А.И., Арасимович В.В., Ярош Н.П. Методы биохимического исследования растений. Л., 1987.
- Калинина А.В., Лящева С.В. Состав и содержание пигментов фотосинтеза в листьях проростков озимой мягкой пшеницы. Известия Самарского научного центра Российской академии наук, 2018, 20(2): 286-290.
- Khalil R.R., Bassiouny F.M., El-Dougdoug K.A., Abo-Elmaty S., Yousef M.S. A dramatic physiological and anatomical changes of tomato plants infecting with tomato yellow leaf curl germinivirus. Journal of Agricultural Technology, 2014, 10(5): 1213-1229.
- Акиньшина Н.Г., Рашидова Д.К., Азизов А.А. Капсулирование семян препаратами хитозана и его производных восстанавливает фотосинтез у растений хлопчатника (Gossypium L., 1753) на фоне вилта. Сельскохозяйственная биология, 2016, 51(5): 696-704 (doi: 10.15389/agrobiology.2016.5.696rus).
- Калинина Е.А. Влияние биологически активных соединений на рост, фотосинтез и продуктивность кукурузы (ZeamaysL.). Известия ТСХ, 2009, 3: 181-186.
- Kilian M., Steiner U., Krebs B., Junge H., Schmeiedeknecht G., Hain R. FZB24® Bacillus subtilis — mode of action of microbial agent enhancing plant vitality. Pflanzenschutz-Nachrichten Bayer, 2000, 1: 72-93.
- Arkhipova T.N., Prinsen E., Veselov S.U., Martynenko E.V., Melentiev A.I., Kudoyarova G.R. Cytokinin producing bacteria enhance plant growth in drying soil. Plant and Soil, 2007, 292: 305-315 (doi: 10.1007/s11104-007-9233-5).
- Cohen A.C., Travaglia C.N., Bottini R., Piccoli P.N. Paticipation of abscisic acid and gibberellins produced by endophytic Azospirillum in the alleviation of drought effects in maize. Botany, 2009, 87: 455-462 (doi: 10.1139/B09-023).
- De Meyer G., Audenaert K., Hufte M. Pseudomonas aeruginosa 7NSK2-induced systemic resistance in tobacco depends on in planta salicylic acid accumulation but is not associated with PR1a expression. European Journal of Plant Pathology, 1999, 105: 513-517 (doi: 10.1023/A:1008741015912).
- Belimov A.A., Dodd I.C., Safronova V.I., Dumova V.A., Shaposhnikov A.I., Ladatko A.G., Davies W.J. Abscisic acid metabolizing rhizobacteria decrease ABA concentrations in planta and alter plant growth. Plant Physiology and Biochemistry, 2014, 74: 84-91 (doi: 10.1016/j.plaphy.2013.10.032).
- Porcel R., Zamarreсo A.M., Garcia-Mina J.M., Aroca R. Involvement of plant endogenous ABA in Bacillus megaterium PGPR activity in tomato plants. BMC Plant Biology, 2014, 14: 36 (doi: 10.1186/1471-2229-14-36).
- Павлюшин В.А., Новикова И.И., Бойкова И.В. Микробиологическая защита растений в технологиях фитосанитарной оптимизации агроэкосистем: теория и практика (обзор). Сельскохозяйственная биология, 2020, 55(3): 421-438 (doi: 10.15389/agrobiology.2020.3.421rus).
- Lizarraga-Pauli E.G., Torres-Pacheco I., Moreno Martinez E., Miranda-Castro S.P. Chitosan application in maize (Zea mays) to сounteract the effects of abiotic stress at seedling level. African Journal of Biotechnology, 2011, 10(34): 6439-6446 (doi: 10.5897/ajb10.1448).
- Mondal M.M.A., Puteh A.B., Dafader N.C., Rafii M.Y., Malek M.A. Foliar application of chitosan improves growth and yield in maize. Journal of Food Agriculture and Environment, 2013, 11: 520-523.
- Martins M., Carvalho M., Carvalho D.T., Barbosa S., Doriguetto A.C., Magalhaes P.C., Ribeiro C. Physicochemical characterization of chitosan and its effects on early growth, cell cycle and root anatomy of transgenic and non-transgenic maize hybrids. Australian Journql of Crop Science, 2018, 12: 56 (doi: 10.21475/ajcs.18.12.01.pne649).
- Ibrahim E.A., Ramadan W.A. Effect of zinc foliar spray alone and combined with humic acid or/and chitosan on growth, nutrient elements content and yield of dry bean (Phaseolus vulgaris L.) plants sown at different dates. Sci. Hort., 2015, 184: 101-105 (doi: 10.1016/j.scien ta.2014.11.010).
- Wang X., Vigjevic M., Liu F., Jacobsen S., Jiang D., Wollenweber B. Drought priming at vegetative growth stages improves tolerance to drought and heat stresses during grain filling in spring wheat (Triticum aestivum L. cv. Vinjett). Plant Growth Regulation, 2015, 75: 677-687 (doi: 10.1007/s1072 5-014-9969-x).
- Pongprayoon W., Roytrakul S., Pichayangkura R., Chadchawan S. The role of hydrogen peroxide in chitosan-induced resistance to osmotic stress in rice (Oryza sativa L.). Plant Growth Regulation, 2013, 70: 159-173 (doi: 10.1007/s10725-013-9789-4).
- Costales M., Falcon R. Combination of application forms of chitosan in the development of biofertilized soybean. Cultivos Tropicales, 2018, 39(3): 71-79.
- Chamnanmanoontham N., Pongprayoon W., Pichayangkura R., Roytrakul S., Chadchawan S. Chitosan enhances rice seedling growth via gene expression network between nucleus and chloroplast. Plant Growth Regulation, 2015, 75: 101-114 (doi: 10.1007/s10725-014-9935-7).
- Farouk S., Amany A.R. Improving growth and yield of cowpea by foliar application of chitosan under water stress. Egyptian Journal of Biology, 2012, 14: 14-26 (doi: 10.4314/ejb.v14i1.2).
- Ашаева О.В., Монин Д.С. Пигментный состав листьев различных сортов яровой пшеницы. Вестник Нижегородской государственной сельскохозяйственной академии, 2012, 1: 111-113.
- Shehata S.A., Fawzy Z.F., El-Ramady H.R. Response of cucumber plants to foliar application of chitosan and yeast under greenhouse conditions. Australian Journal of Basic and Applied Sciences, 2012, 6(4): 63-71.