doi: 10.15389/agrobiology.2023.1.75rus
УДК 633.12:631.671:581.113:58.1/.2
Исследование выполнено за счет гранта Российского научного фонда № 22-26-00041, https://rscf.ru/pro-ject/22-26-00041/ в рамках тематического плана ЦКП Орловского ГАУ «Генетические ресурсы растений и их использование» по совместной программе с селекционерами ФГБНУ ФНЦ зернобобовых и крупяных культур (ФГБНУ ФНЦ ЗБК).
ЭФФЕКТИВНОСТЬ ИСПОЛЬЗОВАНИЯ ВОДЫ ЛИСТЬЯМИ Fagopyrum esculentum Moench ПОД ВЛИЯНИЕМ ЭКЗОГЕННЫХ И ЭНДОГЕННЫХ ФАКТОРОВ
А.В. АМЕЛИН1 ✉, А.Н. ФЕСЕНКО2, В.В. ЗАИКИН1, Е.И. ЧЕКАЛИН1 , Р.А. ИКУСОВ1
На устойчивости развития современного сельского хозяйства негативно сказывается усиливающаяся аридизация климата на планете. Поэтому важное значение имеет эффективность использования воды растениями, на что влияют различные эндогенные и экзогенные факторы. В настоящей работе мы впервые установили этот показатель для листьев растений гречихи в зависимости от фазы роста, времени суток и погодных условий вегетации, а также выявили характер его связи с интенсивностью фотосинтеза, транспирации и семенной продуктивностью. Цель исследования — обнаружить видовые особенности потребления воды при осуществлении фотосинтеза и продукционного процесса у растений гречихи обыкновенной (Fagopyrum esculentum Moench). Объектами изучения были 22 сортообразца гречихи обыкновенной: К-406 и К-1709 (местные популяции); Калининская, Богатырь, Шатиловская 5 (старые сорта); Дикуль, Дождик, Деметра, Девятка, Дизайн (современные районированные сорта); Башкирская красностебельная, Батыр, Уша, Чатыр-Тау, Инзерская, Дизайн 2, Р 66, Р 69, Р 70, Р 84, Р 85, СПР 52 (сортообразцы, перспективные для разных условий). Сортообразцы выращивали в селекционном севообороте ФНЦ зернобобовых и крупяных культур (Орловская обл., Орловский р-н) в 2010-2015 годах на делянках площадью 10 м2 в 4-кратной повторности рандомизированным методом. Интенсивность фотосинтеза (ИФ), интенсивность транспирации (ИТ) и устьичную проводимость оценивали по оригинальной методике фирмы «Heinz Walz GmbH» (Германия) с помощью переносного газоанализатора GFS-3000 FL. Учет осуществляли на 5-7 типичных для генотипа растениях, произрастающих в середине делянки, у которых листья не имели повреждений вредителями и поражений болезнями. Замеры проводили в режиме реального времени в основные фазы роста (ветвление, цветение + 10 сут, цветение + 20 сут, цветение + 30 сут) на 3-м листе сверху главного стебля с 700 до 1900 с периодичностью 3 ч. В измерительной камере прибора поддерживали интенсивность света 1000 мкмоль фотонов·м-2·с-1, температура воздуха составляла 25 °С. Эффективность использования воды (ЭИВ) определяли, вычисляя отношение текущих значений интенсивности фотосинтеза к значениям интенсивности транспирации. Урожайность зерна учитывали с каждой делянки сорта прямым взвешиванием и по результатам структурного анализа растений. В результате проведенных исследований установлено, что у культуры гречихи эффективность использования воды растениями находилась в существенной зависимости как от условий произрастания, так и от наследственных особенностей. Под влиянием погодных условий вегетации ЭИВ варьировала от 1,03 до 2,08 мкмоль CO2/ммоль Н2O. Наиболее высокое ее значение (2,08 мкмоль CO2/ммоль Н2O) отмечали в 2012 году, когда погода была относительно благоприятной для роста и развития растений. В онтогенезе максимальную эффективность использования воды в осуществлении фотосинтеза фиксировали в фазу ветвления (в среднем 2,43 мкмоль CO2/ммоль Н2O) и массового налива семян (1,78 мкмоль CO2/ммоль Н2O), а самую низкую — во время бутонизации и цветения (в среднем 1,17 мкмоль CO2/ммоль Н2O). Больше всего молекул СО2 на единицу количества воды, испаренной единицей листовой поверхности, ассимилировалось с 900 до 1100, когда отмечалась наиболее высокая интенсивность фотосинтеза в листьях и умеренная активность транспирации. Коэффициент корреляции между ЭИВ и интенсивностью фотосинтеза в листьях был положительным (r = 0,69, р < 0,05), между ЭИВ и интенсивностью транспирации — отрицательным (r = -0,89, р < 0,05), что соответствующим образом повлияло на урожайность. Сортообразцы гречихи существенно различались по ЭВИ. В результате селекции значение ЭВИ выросло (P = 0,95) в среднем на 20,5 %, что было обусловлено увеличением ИФ на 29,0 %, ИТ — на 7,9 %, в том числе благодаря усилению устьичной проводимости листьев в среднем на 18,1 %.
Ключевые слова: Fagopyrum esculentum, гречиха, интенсивность фотосинтеза, интенсивность транспирации, эффективность использования воды, урожайность.
Для современного сельского хозяйства характерна низкая устойчивость развития, в том числе из-за усиливающейся аридизации климата (1, 2). В связи с этим важное значение имеют процессы, связанные с испарением воды растениями (3-5), которые зависят как от экзогенных, так и эндогенных факторов роста и развития (6-8). Из экзогенных факторов определяющую роль играют режим увлажнения и температура воздуха (9), из эндогенных — фотосинтез и транспирация листьев в силу их значимой физиологической роли в продукционном процессе и защитной системе растения (10, 11). На важность изучения эффективности использования воды листьями растений указывают многие исследователи (12, 13).
Эффективность использования воды (ЭИВ) обычно оценивается по трем показателям. Это ассимиляционно-транспирационное отношение (АТО) — отношение текущих значений интенсивности фотосинтеза к транспирации; продуктивность транспирации (ПТ) — сухая биомасса, сформированная на единицу транспирируемой растением воды; изотопная дискриминация углерода (D13С) — характеризует АТО в отдельном листе (10). Несмотря на определенные различия, в основе всех этих показателей находится сопряженность процессов газо- и водообмена на разных уровнях организации растения.
При изучении взаимосвязей между параметрами, определяющими ЭИВ и засухоустойчивость, на 120 гибридах пшеницы F2 были установлены QTL (quantitative trait loci), связанные с эффективностью использования воды, и показано, что накопление биомассы положительно коррелирует с АТО, удельной поверхностной плотностью листьев (УПП) и ИТ, но отрицательно — с D13С (9).
Исследование этих вопросов актуально и для такой востребованной на мировом продовольственном рынке культуры, как гречиха, расширение посевных площадей которой во многом сдерживается низкими адаптивными способностями растений (14). Известно, что гречиха расходует в 2-3 раза больше воды, чем просо и другие культуры (15-17). При этом в процессе селекции устойчивость растений к ограничению влагообеспеченности не только не повышается, а имеет выраженную тенденцию к снижению (18). У современных сортов гречихи в сухую и жаркую погоду резко уменьшается урожайность зерна (19).
В настоящей работе мы впервые получили данные об эффективности использования воды листьями растений гречихи в зависимости от фазы роста, времени суток и погодных условий вегетации, а также описали характер ее связи с интенсивностью фотосинтеза, транспирации и семенной продуктивностью.
Цель исследования — выявить видовые особенности потребления воды при осуществлении фотосинтеза и продукционного процесса у растений гречихи обыкновенной (Fagopyrum esculentum Moench).
Методика. Объектами изучения были 22 сортообразца гречихи обыкновенной. Из них 10 созданы в период селекции культуры от местных популяций до лучших современных районированных сортов: К-406 и К-1709 (местные популяции); Калининская, Богатырь, Шатиловская 5 (старые сорта); Дикуль, Дождик, Деметра, Девятка, Дизайн (современные районированные сорта). Другие 12 сортообразцов изучали как перспективные для разных условий: Башкирская красностебельная, Батыр, Уша, Чатыр-Тау, Инзерская, Дизайн 2, Р 66, Р 69, Р 70, Р 84, Р 85, СПР 52.
Изучаемые сортообразцы выращивали в селекционном севообороте ФНЦ зернобобовых и крупяных культур (Орловской обл., Орловский р-н) в 2010-2015 годах на делянках площадью 10 м2 в 4-кратной повторности рандомизированным методом. Уход за посевами и уборку выполняли согласно методическим рекомендациям для региона.
Дополнительно для пяти сортообразцов гречихи (К-406, К-1709, Богатырь, Дикуль, Дождик) были проведены две серии модельных вегетационных опытов по изучению дневной динамики интенсивности фотосинтеза, интенсивности транспирации и эффективности использования воды. Для этого растения выращивали в специальных сосудах объемом 9 л (по 5 типичных для сорта в каждом сосуде, повторность 4-кратная) при влажности почвы 30 и 70 % от полной влагоемкости.
В вегетационном опыте анализируемые показатели определяли для каждого сортообразца. В мелкоделяночных опытах учет осуществляли на 5-7 типичных для генотипа растениях, у которых листья не имели повреждений вредителями и поражений болезнями. Замеры проводили в режиме реального времени в основные фазы роста (ветвление, цветение + 10 сут, цветение + 20 сут, цветение + 30 сут) на 3-м сверху (физиологически зрелом) листе главного стебля.
Интенсивность фотосинтеза (ИФ), интенсивность транспирации (ИТ) и устьичную проводимость (УП) оценивали по оригинальной методике фирмы «Heinz Walz GmbH» (Германия) с помощью переносного газоанализатора GFS-3000 FL. В вегетационном опыте измерения проводили с 700 до 1900 с периодичностью 3 ч, в мелкоделяночном — с 800 до 1100. В измерительной камере прибора поддерживали интенсивность света 1000 мкмоль фотонов·м-2·с-1 и температуру воздуха 25 °С.
ЭИВ определяли, вычисляя отношение текущих значений интенсивности фотосинтеза к значениям интенсивности транспирации (10, 20).
Урожайность зерна учитывали с каждой делянки сорта прямым взвешиванием на весах ВК-600 (ЗАО «Масса-К», Россия) и по результатам структурного анализа 15 растений в 4-кратной повторности.
Статистическую обработку полученных результатов (дисперсионный и корреляционный анализ) проводили с помощью программных пакетов Microsoft Excel 2013 и Statistica v. 10.0 («StatSoft, Inc.», США). Достоверность различий определяли по t-критерию Стьюдента при P = 0,95. Рассчитывали средние значения (М) и стандартные отклонения (±SD).
Результаты. Годы исследований характеризовались разными погодными условиями. Наиболее экстремальным был 2010 год: за полный период вегетации растений выпало на 54,9 % меньше осадков, а среднемесячная температура воздуха была на 5,5 °С выше среднемноголетних значений. Наиболее комфортными для растений гречихи оказались условия 2012 года: количество выпавших осадков и среднемесячная температура за период вегетации были близки среднемноголетней норме. Метеоусловия вегетации в 2011 и 2013-2015 годах в отдельные фазы роста были жесткими, но не экстремальными.
Установлено, что у растений гречихи, как и у других сельскохозяйственных культур (11, 21, 22) эффективность использования воды тесно связана с погодными условиями. Ее величина в годы исследований варьировала от 1,03 до 2,08 мкмоль CO2/ммоль Н2O. Наиболее высокая эффективность использования воды на фотосинтез отмечалась в 2012 году, наименьшая — в 2013-2015 годы. В 2012 году значение ЭИВ было выше (при P = 0,95) в среднем на 14,9 % по сравнению с 2010, на 27,4 % — с 2011, на 50,5 % — с 2013, на 44,7 % — с 2014 году, на 35,6 % — с 2015 годом. Влияние погодных условий вегетации на эффективность использования воды было опосредовано через воздействие на процессы транспирации и фотосинтеза (рис. 1).
Коэффициент корреляции между ЭИВ и интенсивностью фотосинтеза листьев был положительным (r = +0,69, р < 0,05), между ЭИВ и интенсивностью транспирации — отрицательным (r = -0,89, р < 0,05). Поэтому закономерно, что в 2012 году высокая эффективность использования воды была обусловлена повышенной интенсивностью фотосинтеза и умеренной активностью транспирации. В этот год сложились благоприятные погодные условия для вегетации растений гречихи, поэтому не было особой надобности тратить большое количество энергии на испарение воды, в основном происходило образование органического вещества и урожайность оказалась наиболее высокой — 3,5 т/га (18, 19).
В то же время в более жестких погодных условиях вегетации 2013-2015 годов растения испытывали потребность в защите от перегрева, что обеспечивалось повышенной транспирацией в ущерб фотосинтезу: эффективность использования воды на поддержание фотосинтеза в эти годы была снижена по сравнению с более благоприятными условиями в среднем на 34,6 %, а семенная продуктивность — на 25,4 % (P = 0,95).
Однако в стрессовых ситуациях направленность ответных физиологических реакций растений была несколько иной. В 2010 году, в условиях ярко выраженной почвенной и воздушной засухи (гидротермический коэффициент — 0,36), чтобы избежать обезвоживания клеток и сохранить их жизнеспособность, растения были вынуждены весьма экономно расходовать воду и энергию. Это достигалось за счет низкой скорости транспирации и фотосинтеза, что в результате приводило к снижению эффективности продукционного процесса в целом. В модельных вегетационных опытах при уменьшении влажности почвы с 70 до 30 % от полной влагоемкости у растений гречихи интенсивность фотосинтеза снижалась в 4,4 раза, интенсивность транспирации — более чем на 35 %, продуктивность — на 43,6 % (P = 0,95).
Как следствие, в 2010 году эффективность использования воды у растений всех сортообразцов была выше средней, а общая и семенная продуктивность — меньше в среднем на 66,6 % (P = 0,95) по сравнению с менее экстремальными условиями вегетации (табл. 1).
Резкое снижение интенсивности фотосинтеза во время засухи и, как следствие, продуктивности может быть обусловлено как недостаточным обеспечением водой и элементами питания листьев из-за низкой транспирационной активности, так и нарушением работы реакционных центров фотосистем (23).
Эффективность использования воды растениями гречихи варьировала в онтогенезе от 1,17 до 2,43 мкмоль CO2/ммоль Н2O. Самое высокое значение отмечали в фазу ветвления и в период массового налива семян: ЭИВ в период вегетативного роста была в среднем на 52 и 34 % больше по сравнению с фазами цветение + 10 сут и цветение + 30 сут. Изменчивость показателя и в этом случае имела тесную связь с характером проявления фотосинтетической и транспирационной активности листьев.
Повышение интенсивности фотосинтеза наблюдалось в основном до фазы цветение + 20 сут, а затем резко снижалось, тогда как скорость транспирации была наибольшей в фазу цветение + 10 сут, а в период вегетативного роста и массового налива семян (фаза цветение + 20 сут) — минимальной (4,31 ммоль Н2O·м-2·с-1) или умеренной (5,92 ммоль Н2O·м-2·с-1), что соответствующим образом отразилось на эффективности использования воды растениями (рис. 2).
Проявление определенной несопряженности между интенсивностью фотосинтеза и транспирации объясняется тем, что молекулы CO2 и Н2О используют один путь попадания в лист — через устьица. Однако диффузный градиент, управляющий транспирацией, примерно в 50 раз больше, чем при поглощении углекислоты, что находит свое отражение в эффективности использования воды растениями в онтогенезе (10, 24). У сельскохозяйственных культур ее величина обычно низкая, поскольку растения теряют воды почти в 100 раз больше, чем ассимилируют эквивалентные единицы углерода в результате фотосинтеза. Для увеличения производства продукции растениеводства важно уделять внимание повышению ЭИВ у культурных видов растений (11). Анализ эффективности использования воды растениями сои показал, что увеличение ЭИВ в листовых пластинах на 1 % в масштабах поля приводит к повышению ЭИВ примерно на 10 % (25). Предлагается использовать этот показатель в качестве вторичного критерия для отбора генотипов культуры по урожайности семян (26). Установлено, что по абсолютной величине наиболее близким к показателю ЭИВ в полевых условиях является так называемая кратковременно (instantaneous, i) измеренная ЭИВ (ЭИВi) листа, рассчитанная по величине дыхания (27).
В течение светового времени суток наиболее высокая скорость ассимиляции CO2 в расчете на единицу количества воды, испаренной единицей листовой поверхности, у растений гречихи наблюдалась в предобеденное время (с 900 до 1100), при повышении солнечной радиации, когда температура воздуха еще невысокая (20-25 °С). В этот период отмечался пик фотосинтетической активности и умеренная транспирация. После 1100 значения ЭИВ заметно снижались, прежде всего вследствие выраженного падения активности фотосинтеза и усиления транспирации из-за высокой температуры воздуха и инсоляции (рис. 3). Очевидно, что в послеобеденное время транспирационный процесс в листьях направлен в большей степени на недопущение перегрева надземных органов, что уменьшает возможности фотосинтеза. Снижение ЭИВ в полуденное время наблюдали и у растений такой теплолюбивой культуры, как соя, что также объясняется повышенными величинами интенсивности транспирации и ростом ФАР (фотосинтетически активная радиация) (27).
Важно отметить, что в результате селекции эффективность использования воды растениями гречихи существенно возросла, в первую очередь из-за увеличения интенсивности фотосинтеза во время формирования и массового налива семян. За период селекции культуры гречихи от местных популяций до лучших современных районированных сортов значения ЭИВ растений стали выше (P = 0,95) в среднем на 20,5 %, ИФ — на 29,0 %, ИТ — на 7,9 %, чему в значительной степени способствовало усиление устьичной проводимости листьев (табл. 2). Аналогичные изменения наблюдаются и при селекции твердой и мягкой пшеницы, при переходе от старых сортов к современным (28, 29).
Это в известной степени соответствует эволюционным изменениям растений, сопровождающимся повышением (30, 31) или оптимизацией водоиспользования (31, 32). Стратегия заключается в открытии устьиц на столько, чтобы обеспечить поглощение необходимое количество молекул CO2 при фотосинтезе и при этом избежать обезвоживания тканей в процессе транспирации. В этом случае отношение ИФ к УП остается достаточно постоянным в широком диапазоне действия факторов (32).
Во многом схожие тенденции по характеру проявления выявлены нами и у других культурных видов растений (22, 33), хотя имеются и определенные различия. Например, у яровой и озимой пшеницы в одинаковых погодных и агротехнических условиях произрастания эффективность использования воды растениями была в среднем на 23,4 и 35,1 % выше, чем у гречихи (рис. 4).
По годам исследований ЭИВ изменялась у растений яровой пшеницы от 1,63 до 2,73, у озимой — от 1,88 до 4,19 мкмоль CO2/ммоль Н2О, что оказывало значимое влияние на формирование урожая, главным образом в жестких погодных условиях вегетации растений. В засушливом 2018 году по этому показателю высокоурожайные сорта яровой пшеницы превосходили средне- и низкоурожайные в среднем на 70 % (P = 0,95) (22). При этом у озимой пшеницы выявлена тесная связь ЭИВ с содержанием в зерне белка и клейковины (33).
У растений Fagopyrum esculentum высокая эффективность использования растениями воды и урожайность семян отмечались в основном в благоприятных погодных условиях вегетации. Это указывает на их более низкие адаптивные возможности, особенно в условиях засухи, очевидно, по причине формирования большей (на 40,8 %) испаряющей поверхности листьев, повышенной (на 21,2 %) интенсивности транспирации и пониженной (на 13,9 %) активности фотосинтеза по сравнению с яровой и озимой пшеницей. Выявленные видовые особенности культурных растений, по-видимому, обусловлены различными условиями их эволюции. Гречиха, как известно, субтропическая культура, развитие которой проходило в Южном Китае, в условиях повышенного увлажнения и высоких температур воздуха (34), тогда как исторические корни происхождения пшеницы, согласно Н.И. Вавилову (35), берут свое начало из Юго-Западной Азии — из мест с засушливым климатом.
О правильности высказанного предположения также свидетельствуют результаты исследований Д.А. Ронжиной с соавт. (36). Изучая структурно-функциональные параметры листьев у 21 вида растений, произраставших в различных условиях водообеспечения, они пришли к заключению, что при уменьшении степени гидрофильности в ряду гелофиты—гигрофиты происходит структурная перестройка листа в сторону увеличения его плотности и содержания сухого вещества, что сопровождается уменьшением скорости транспирации и закономерным возрастанием эффективности использования воды (36).
Повысить эффективность использования воды растениями можно как агротехническими, так и селекционными способами. По мнению Е.И. Кошкина (9), наиболее простой путь для зерновых культур С3 — это совмещение периода накопления максимальной биомассы с наиболее прохладными погодными условиями, для чего необходимо изменить сроки посева. При этом селекцию следует направить на активизацию процессов начального роста, что приведет к увеличению индекса листовой поверхности, поглощения ФАР и, как следствие, возрастанию ЭИВ (9).
Для достижения этой цели у гречихи, на наш взгляд, важно добиться улучшения светопропускной способности агроценоза посредством регулирования нормы высева и оптимизации архитектоники растений. Целесообразно отбирать формы с меньшей листовой поверхностью, но плотными листовыми пластинками, то есть вести селекцию на ксероморфность. Новые сорта должны также обладать повышенной активностью фотосинтеза при умеренной транспирации. Работа в этом направлении вполне может быть успешной, поскольку генофонд гречихи характеризуется широким полиморфизмом показателей фотосинтетической и транспирационной активности (37, 38).
Суммируя, отметим, что, в отличие от яровой и озимой пшеницы, у гречихи Fagopyrum esculentum Moench высокая эффективность использования воды и урожайность семян отмечались в основном при благоприятных погодных условиях вегетации. Это указывает на более низкие адаптивные возможности (особенно при воздействии засухи) из-за формирования большей (на 40,8 %) испаряющей поверхности листьев, повышенной (на 21,2 %) интенсивности транспирации и пониженной (на 13,9 %) активности фотосинтеза. Выявленные различия между культурными видами растений во многом обусловлены неодинаковыми условиями их эволюционного формирования. Гречиха, как известно, субтропическая культура, развитие которой происходило в Южном Китае, при повышенном увлажнении и высоких температурах.
Таким образом, на эффективность использования воды (ЭИВ) растениями гречихи обыкновенной существенное влияние оказывают наследственные особенности и погодные условия произрастания. При повышенной температуре воздуха и ограниченном количестве осадков ЭИВ снижается, а в благоприятных погодных условиях повышается. Из эндогенных факторов положительное влияние на ЭИВ растений оказывает интенсивность фотосинтеза листьев (r = +0,69, р < 0,05), тогда как интенсивность транспирации — отрицательное (r = -0,89, р < 0,05). В результате селекции значение ЭВИ выросло (P = 0,95) в среднем на 20,5 %, что было обусловлено увеличением интенсивности фотосинтеза на 29,0 %, интенсивности транспирации — на 7,9 %, в том числе благодаря усилению устьичной проводимости листьев в среднем на 18,1 %. Это оказывало положительное влияние на рост урожайности культуры лишь в оптимальных условиях вегетации, потому что устойчивость растений к воздушной и почвенной засухе уменьшилась. Отбор форм с повышенной активностью фотосинтеза и умеренной транспирациейможет рассматриваться как одно из приоритетных направлений селекции культуры.
1. Эффективность использования воды (ЭИВ, мкмоль CO2/ммоль Н2O), масса семян и сухая масса надземных органов у 11 соортообразцов гречихи обыкновенной (Fagopyrum esculentum Moench) в разные годы исследования (М±SD; ФГБНУ ФНЦ зернобобовых и крупяных культур, Орловская обл., Орловский р-н) |
||||||
Показатель |
Год |
|||||
2010 |
2011 |
2012 |
2013 |
2014 |
2015 |
|
ЭИВ |
1,77±0,24 |
1,51±0,12 |
2,08±0,26 |
1,03±0,08 |
1,15±0,10 |
1,34±0,14 |
Сухая масса надземных органов, г/растение |
3,20±0,40 |
5,10±0,65 |
6,70±0,13 |
6,04±0,55 |
5,09±0,46 |
5,38±0,49 |
Масса семян, г/растение |
0,52±0,17 |
1,41±0,45 |
1,77±0,29 |
1,60±0,48 |
1,54±0,37 |
1,51±0,35 |
Примечание. Значения ЭИВ приведены для периода массового формирования плодов (фаза цветение + 10 сут), для их расчета использовали данные, полученные с 800 до 1100 (по каждому сорту n = 5). Сухую массу надземных органов определяли в фазу уборочной спелости (у каждого сорта анализировали по 15 растений в 4-кратной повторности). Описании сортообразцов см. в разделе «Методика». |
||||||
2. Интенсивность фотосинтеза, интенсивность транспирации, устьичная проводимость и эффективность использования воды у 10 местных популяций и сортообразцов гречихи обыкновенной (Fagopyrum esculentum Moench) разных периодов селекции в фазу цветение + 20 сут (по каждому образцу n= 5, М±SD; ФГБНУ ФНЦ зернобобовых и крупяных культур, Орловская обл., Орловский р-н, 2013-2015 годы) |
||||
Сортообразец |
Интенсивность фотосинтеза, мкмоль CO2·м-2·с-1 |
Интенсивность транспирации, ммоль Н2О·м-2·с-1 |
Устьичная проводимость, моль Н2О·м-2·с-1 |
Эффективность использования воды, мкмоль CO2/ммоль Н2О |
Местные популяции (Орловская обл.) |
||||
К-406 |
9,55±0,30 |
6,20±0,21 |
0,452±0,025 |
1,54±0,28 |
К-1709 |
11,53±0,29 |
7,28±0,20 |
0,441±0,023 |
1,58±0,30 |
Сорта селекции 1930-1960-х годов |
||||
Калининская |
11,76±0,31 |
7,61±0,22 |
0,447±0,024 |
1,55±0,24 |
Богатырь |
12,35±0,27 |
7,37±0,19 |
0,465±0,025 |
1,68±0,10 |
Шатиловская 5 |
9,76±0,25 |
6,47±0,17 |
0,472±0,027 |
1,51±0,31 |
Сорта селекции 1990-2010-х годов |
||||
Деметра |
13,97±0,29 |
8,07±0,17 |
0,551±0,027 |
1,73±0,27 |
Дождик |
13,28±0,27 |
7,36±0,19 |
0,564±0,026 |
1,80±0,28 |
Дикуль |
15,58±0,28 |
7,84±0,21 |
0,553±0,025 |
1,99±0,44 |
Девятка |
13,22±0,30 |
6,49±0,20 |
0,480±0,024 |
2,04±0,51 |
Дизайн |
11,96±0,31 |
6,58±0,22 |
0,492±0,026 |
1,82±0,36 |
НСР05 |
0,31 |
0,22 |
0,027 |
0,52 |
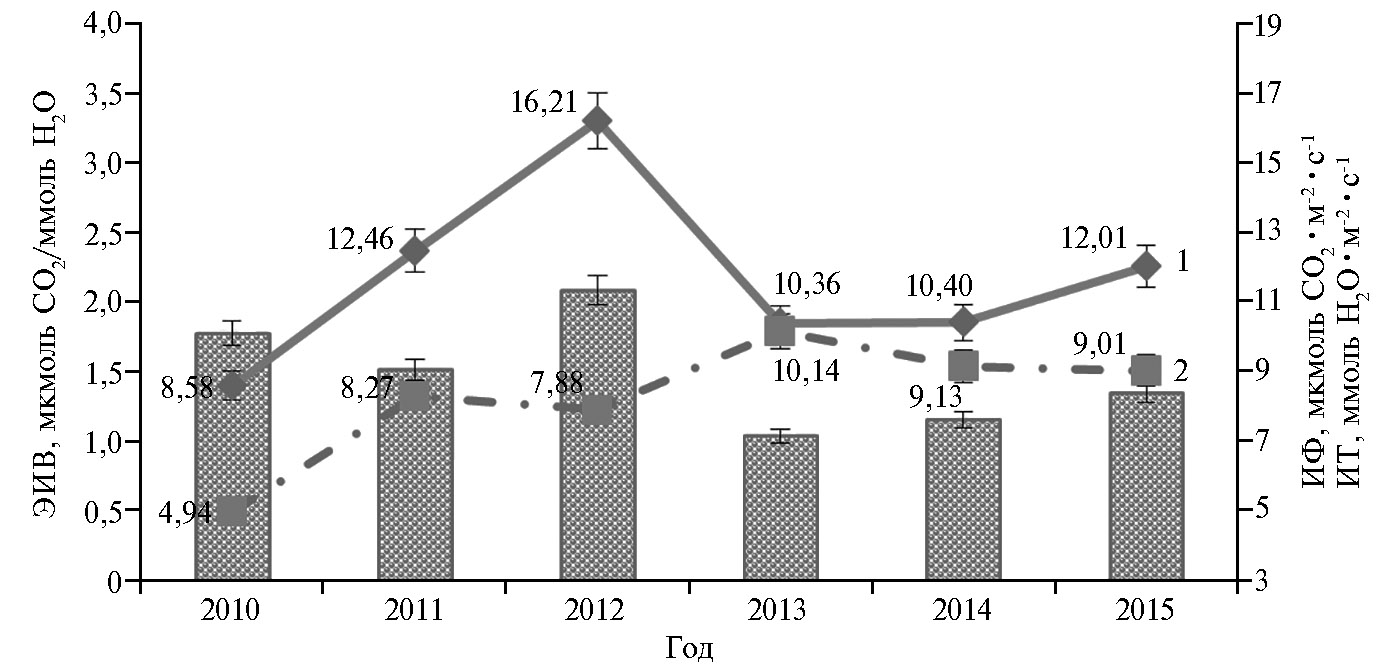
Рис. 1. Эффективность использования воды (ЭИВ) (диаграмма), интенсивность фотосинтеза (ИФ) (1) и транспирации (ИТ) (2) в листьях соортообразцов гречихи обыкновенной(Fagopyrum esculentum Moench) в разные годы в фазу цветение + 10 сут (среднее по 22 соортообразцам, для каждого сортообразца n = 5, М±SD; ФГБНУ ФНЦ зернобобовых и крупяных культур, Орловская обл., Орловский р-н). Описании сортообразцов см. в разделе «Методика».
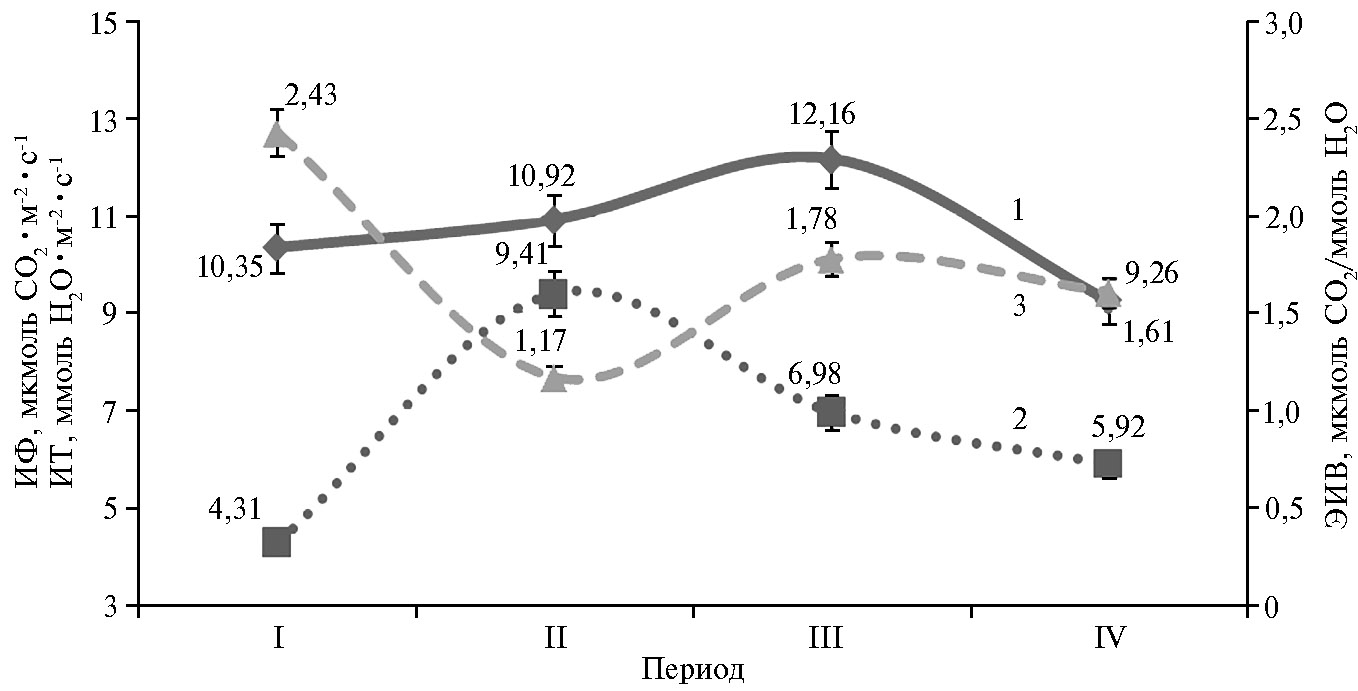
Рис. 2. Интенсивность фотосинтеза (ИФ) (1), интенсивность транспирации (ИТ) (2) и эффективность использования воды (ЭИВ) (3) листьями растений гречихи обыкновенной (Fagopyrum esculentum Moench) сортообразцов К-406, К-1709, Калининская, Богатырь, Шатиловская 5, Деметра, Дождик, Дикуль, Инзерская, Девятка, Дизайн в разные фазы роста и периоды развития: I — вегетативный рост, II — цветение + 10 сут, III — цветение + 20 сут, IV — цветение + 30 сут (среднее по 11 соортообразцам, для каждого сортообразца n = 5, М±SD; ФГБНУ ФНЦ зернобобовых и крупяных культур, Орловская обл., Орловский р-н, 2013-2015 годы).
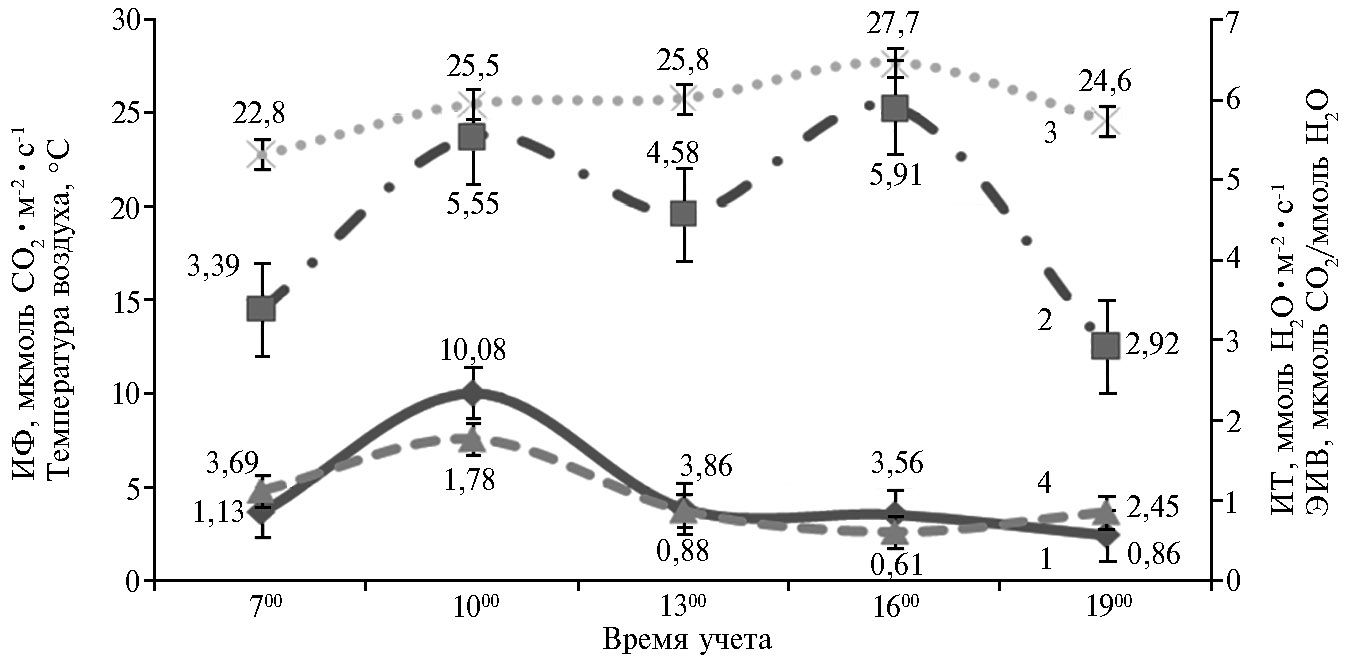
Рис. 3. Дневная динамика интенсивности фотосинтеза (ИФ) (1), интенсивности транспирации (ИТ) (2), температуры воздуха (3) и эффективности использования воды (ЭИВ) (4) листьями растений гречихи обыкновенной (Fagopyrum esculentum Moench) сортообразцов К-406, К-1709, Богатырь, Дикуль, Дождик в фазу цветение + 20 сут по данным двух серий вегетационных опытов (n = 5, М±SD; ФГБНУ ФНЦ зернобобовых и крупяных культур, Орловская обл., Орловский р-н, 2014-2015 годы).
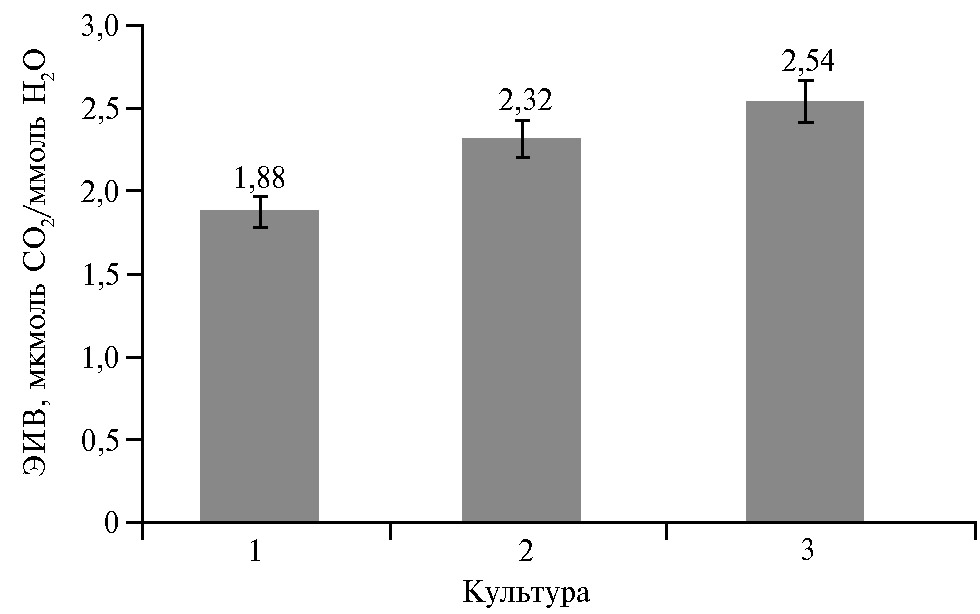
Рис. 4. Эффективность использования воды в период формирования зерновок у разных сельскохозяйственных культур: 1 — гречиха Fagopyrum esculentum Moench современных районированных сортов Деметра, Дождик, Дикуль, Девятка, Дизайн (по каждому сорту n = 5, М±SD, ФГБНУ ФНЦ зернобобовых и крупяных культур, Орловский р-н, Орловская обл., 2013-2015 годы); 2 — яровая пшеница Triticum aestivum L. сортов Бурлак, Воронежская 13, Воронежская 18, Рима, Ульяновская 105, Черноземноуральская 2, Воронежская 20, Лиза, Тулайковская Надежда, Хуторянка, Юбилейная 80, Арсея, Аль Варис, Злата, Радмира, Triticum durum Desf. сортов Донэла М, Мелодия Дона, Безенчукская Нива, Безенчукская 210, Донская Элегия); 3 — озимая пшеница Triticum aestivum L. сортов Московская 40, Московская 39, Немчиновская 17, Аскет, Поволжская Нива, Ангелина, Морозко, Московская 56, Бирюза, Октава 15, Черноземка 130, Немчиновская 57, Юка, Ариадна, Донэра (n = 5, М±SD) (33) (ФГБНУ ФНЦ зернобобовых и крупяных культур, Орловская обл., Орловский р-н, 2017-2019 годы).
ЛИТЕРАТУРА
- Куликов А.И., Убугунов Л.Л., Мангатаев А.Ц. О глобальном изменении климата и его экосистемных следствиях. Аридные экосистемы, 2014, 3(60): 5-13.
- Stuart D., Schewe R.L. Constrained choice and climate change mitigation in US agriculture: structural barriers to a climate change ethic. Journal of Agricultural and Environmental Ethics, 2016, 29: 369-385 (doi: 10.1007/s10806-016-9605-z).
- Zhang Y., Yu X., Chen L., Jia G. Whole-plant instantaneous and short-term water-use efficiency in response to soil water content and CO2 concentration. Plant Soil, 2019, 444: 281-298 (doi: 10.1007/s11104-019-04277-6).
- Tränkner M., Jákli B., Tavakol E., Geilfus C.-M., Cakmak I., Dittert K., Senbayram M. Magnesium deficiency decreases biomass water-use efficiency and increases leaf water-use efficiency and oxidative stress in barley plants. Plant Soil, 2016, 406: 409-423 (doi: 10.1007/s11104-016-2886-1).
- Karthika G., Kholova J., Alimagham S., Ganesan M., Chadalavada K., Kumari R., Vadez V. Measurement of transpiration restriction under high vapor pressure deficit for sorghum mapping population parents. Plant Physiology Reports, 2019, 24: 74-85 (doi: 10.1007/s40502-019-0432-x).
- Li Y., Li H., Li Y., Zhang S. Improving water-use efficiency by decreasing stomatal conductance and transpiration rate to maintain higher ear photosynthetic rate in drought resistant wheat. The Crop Journal, 2017, 5(3): 231-239 (doi: 10.1016/j.cj.2017.01.001).
- Zhu X., Yu G., Wang Q., Hu Z., Han S., Yan J., Wang Y., Zhao L. Seasonal dynamics of water use efficiency of typical forest and grassland ecosystems in China. Journal of Forest Research, 2014, 19(1): 70-76 (doi: 10.1007/s10310-013-0390-5).
- Damour G., Simonneau T., Cochard H., Urban L. An overview of models of stomatal conductance at leaf level. Plant, Cell & Environment, 2010, 33(9): 1419-1438 (doi: 10.1111/j.1365-3040.2010.02181.x).
- Кошкин Е.И. Физиологические основы селекции растений. М., 2014.
- Кошкин Е.И. Физиология устойчивости сельскохозяйственных культур. М., 2010.
- Bramley H., Turner N.C., Siddique K.H.M. Water use efficiency. In: Genomics and breeding for climate-resilient crops /C. Kole (ed.). Springer, Berlin, Heidelberg, 2013: 225-268 (doi: 10.1007/978-3-642-37048-9_6).
- Zhou L., Meng X., Zhang Z., Wu Q. Association analysis of growth characteristics, WUE, and RUE of rice in cold region under different irrigation patterns. Journal of The Institution of Engineers (India): Series A, 2020, 101: 421-431 (doi: 10.1007/s40030-020-00452-6).
- Moradgholi A., Mobasser H., Ganjali H., Fanaie H., Mehraban A. WUE, protein and grain yield of wheat under the interaction of biological and chemical fertilizers and different moisture regimes. Cereal Research Communications, 2022, 50: 147-155 (doi: 10.1007/s42976-021-00145-1).
- Фесенко А.Н., Фесенко И.Н. Результаты селекции, динамика производства и рынок зерна гречихи (анализ многолетних данных). Земледелие, 2017, 3: 24-26.
- Важов В.М. Гречиха на полях Алтая. М., 2013.
- Вавилов П.П., Гриценко В.В., Кузнецов В.С. Растениеводство. М., 1986.
- Лаханов А.П., Коломейченко В.В., Фесенко Н.В., Наполова Г.В., Музалевская Р.С., Савкин В.И., Фесенко А.Н. Морфофизиология и продукционный процесс гречихи. Орел, 2004.
- Амелин А.В., Фесенко А.Н., Чекалин Е.И., Заикин В.В. Адаптивный потенциал фотосинтеза и продукционного процесса у местных форм и сортообразцов гречихи (FagopyrumesculentumMoench) разных периодов селекции. Сельскохозяйственная биология, 2016, 51(1): 79-88 (doi: 10.15389/agrobiology.2016.1.79rus).
- Амелин А.В., Фесенко А.Н., Заикин В.В., Бойко Т.В.Изменчивость элементов структуры урожая у растений гречихи в зависимости от сорта и погодных условий вегетации. Аграрный научный журнал, 2014, 11(23): 3-6.
- Polley W.H. Implications of atmospheric and climate change for crop yield and water use efficiency. Crop Sciense, 2002, 42(1): 131-140 (doi: 10.2135/cropsci2002.1310).
- Ruggiero A., Punzo P., Landi S., Costa A., Van Oosten M.J., Grillo S. Improving plant water use efficiency through molecular genetics. Horticulturae, 2017, 3(2): 31 (doi: 10.3390/horticulturae3020031).
- Амелин А.В., Чекалин Е.И., Заикин В.В., Икусов Р.А., Шишкин А.С. Интенсивность транспирации листьев растений у современных сортов яровой пшеницы. Вестник Курской ГСХА, 2022, 6: 6-12.
- Удовенко Г.В. Устойчивость растений к абиотическим стрессам. В сб.: Физиологические основы селекции растений. СПб, 1995, 2: 293-352.
- Алехина Н.Д, Балнокин Ю.В., Гавриленко В.Ф., Жигалова Т.В. Физиология растений. М., 2007.
- Gorthi A. Quantifying water use efficiency at leaf and field-scales for soybean, miscanthus and switchgrass. Doctoral Thesis. Purdue University, 2017.
- Amoanimaa-Dede H., Su C., Yeboah A., Zhou H., Zheng D., Zhu H. Growth regulators promote soybean productivity: a review. PeerJ, 2022, 10: e12556 (doi: 10.7717/peerj.12556).
- Харчук О.А., Кириллов А.Ф., Будак А.Б. Эффективность использования воды листьями растений сои: традиции и новые критерии. Евразийский Союз Ученых, 2018, 11-1(56): 42-46.
- Li L.H., Chen S.B. Study on root function efficiency of spring wheat under different moisture condition. Sci. Agric. Sin., 2002, 35: 867-871 (in Chinese).
- Fan X.-W., Li F.-M., Xiong Y.-C., An L.-Z., Long R.-.J. The cooperative relation between non-hydraulic root signals and osmotic adjustment under water stress improves grain formation for springwheat varieties. Physiologia Plantarum, 2008, 132(3): 283-292 (doi: 10.1111/j.1399-3054.2007.01007.x).
- Li Y.Y., Zhang S.Q., Shao M.A. Interrelationship between water use efficiency and nitrogen use efficiency of different wheat evolution materials. Chinese Journal of Applied Ecology, 2003, 14(9): 1478-1480 (in Chinese).
- Song L., Li F.M., Fan X.W., Xiong Y.C., Wang W.Q., Wu X.B., Turner N.C. Soil water availability and plant competition affect the yield of spring wheat. European Journal of Agronomy, 2009, 31(1): 51-60 (doi: 10.1016/j.eja.2009.03.003).
- Bacon M. Water use efficiency in plant biology. Armsterdam, 2004.
- Amelin A.V., Chekalin E.I., Zaikin V.V., Mazalov V.I., Ikusov R.A. Biochemical grain quality indicators and photosynthetic rate of leaves in modern varieties of winter wheat. IOP Conference Series: Earth and Environmental Science, 2021, 848: 012096 (doi: 10.1088/1755-1315/848/1/012096).
- Ohnishi O. Distribution and classification of wild buckwheat species. 1. Cymosum group. Fagopyrum, 2010, 27: 1-8.
- Вавилов Н.И. Центры происхождения культурных растений. Труды по прикладной ботанике и селекции,1926, том 16, вып. 2.
- Ронжина Д.А., Рупышев Ю.А., Иванова Л.А., Мигалина С.В., Иванов Л.А. Разнообразие структурно-функциональных параметров фотосинтетического аппарата гидрофильных растений реки Максимиха (Республика Бурятия, Россия). Проблемы ботаники Южной Сибири и Монголии, 2022, 21(2): 171-174 (doi: 10.14258/pbssm.2022077).
- Амелин А.В., Фесенко А.Н., Заикин В.В. Гено- и фенотипические особенности проявления интенсивности фотосинтеза листьев у растений гречихи. Вестник ОрелГАУ, 2015, 6(57): 18-22 (doi: 10.15217/issn1990-3618.2015.6.18).
- Amelin A.V., Fesenko A.N., Zaikin V.V., Chekalin E.I., Ikusov R.A. Transpiration activity of leaves in buckwheat varieties of different breeding periods. BIO Web of Conferences, 2022, 47: 01002 (doi: 10.1051/bioconf/20224701002).