doi: 10.15389/agrobiology.2022.1.141rus
УДК 636.086.1:636.085.19
Исследования поддержаны Российским научным фондом (проект РНФ № 19-76-10031), за исключением экспериментов по выбору наиболее эффективного штамма актинобактерий, проведенных ранее в рамках проекта РНФ № 14-16-00150.
ПОВЫШЕНИЕ ЭФФЕКТИВНОСТИ ДЕКОНТАМИНАЦИИ ЗЕРНА ПШЕНИЦЫ, ЗАГРЯЗНЕННОГО АФЛАТОКСИНОМ В1, ПОСРЕДСТВОМ CОВМЕСТНОГО ИСПОЛЬЗОВАНИЯ АГЕНТОВ БИОДЕГРАДАЦИИ ТОКСИНА И ИНГИБИТОРА ЕГО БИОСИНТЕЗА
Т.М. ВОИНОВА, Л.А. ЩЕРБАКОВА✉, С.Б. ПОПЛЕТАЕВА, В.Г. ДЖАВАХИЯ
Деконтаминация загрязненного микотоксинами фуражного зерна — одна из актуальных задач обеспечения пищевой безопасности сельскохозяйственных кормов. В последние годы на территории Российской Федерации все чаще фиксируются случаи значительного загрязнения фуражного зерна и других кормов афлатоксином В1 (АФВ1). Вероятность афлатоксинового загрязнения травостоя и фуражного зерна злаков, культивируемых в центральных и северных регионах страны, может в дальнейшем возрастать из-за вызванного потеплением климата расширения ареала Aspergillus flavus — основного продуцента АФВ1. К одним из перспективных методов деконтаминации зерна, загрязненного токсином, относится его биодеградация микроорганизмами-деструкторами, которые синтезируют ферменты, детоксицирующие АФВ1. Другим методом может быть обработка инфицированного аспергиллами зерна ингибиторами афлатоксигенеза. В настоящей работе впервые было показано, что обработка супернатантом культуральной жидкости Rhodococcus erythropolus АС-884 вслед за обработкой компактином практически полностью предотвращает накопление микотоксина в зараженном зерне. Цель работы — оценка эффективности снижения содержания микотоксина в зерне пшеницы, искусственно контаминированном афлатоксином В1, после обработки актинобактериями рода Rhodococcus или ингибитором афлатоксигенеза (компактином), а также при сочетании этих методов. Способность к деструкции АФВ1 была протестирована у четырех штаммов:Rhodococcus sp. AC-1260, R. erythropolus AC-1269 и АС-884, R. ruber AC-1801. С помощью количественного анализа АФВ1 методом высокоэффективной жидкостной хроматографии обнаружено, что наиболее активная деградация микотоксина происходила в освобожденном от бактериальных клеток супернатанте культуральной жидкости (КЖ) штамма АС-884. После инкубации АФВ1 (0,2 мкг/мл) в КЖ АС-884 при нейтральном рН в течение 48 ч при 30 °С были обнаружены лишь следовые количества микотоксина, в то время как в супернатантах КЖ других штаммов при тех же условиях сохранялось от 15 до 50 % внесенного АФВ1. Обработка КЖ АС-884 зерна пшеницы (Triticum aestivum L.) сорта Дарья, искусственно загрязненного препаратом АФВ1 (1,0-5,0 мкг/г зерна), через 3 сут приводила к удалению 60 % добавленного токсина, а после использования супернатантов КЖ или клеточных суспензий штаммов AC-1260, AC-1801 или AC-1269 количество АФВ1 оставалось таким же, как в контроле. Обработка зерна супернатантом КЖ штамма АС-884 давала лучший результат, чем обработка аналогичным супернатантом другого биодеструктора — Phoma glomerata PG-41. Через 7 сут после использования компактина (0,05 мг/г) количество токсина в зараженном Aspergillus flavus А11 зерне пшеницы уменьшалось вдвое по сравнению с контролем. Через 1 сут после обработки КЖ АС-884 (0,25 мл/г) образцов зерна, на котором гриб развивался в течение 6 сут, содержание АФВ1 сокращалось почти в 3 раза. В то же время в результате комбинирования обеих обработок (компактин, 0,05 мг/г + КЖ АС-884, 0,25 мл/г) концентрация АФВ1 в экстрактах зараженного зерна снижалась более чем в 200 раз, и при этом достигалась практически полная очистка его от микотоксина. Таким образом, подход, основанный на сочетании биодеградации с ингибированием афлатоксигенеза, может обеспечивать эффективную деконтаминацию зерна, загрязненного продуцентами АФВ1, в тех случаях, когда использование ингибиторов не приводит к полному подавлению биосинтеза микотоксина.
Ключевые слова: афлатоксин В1, корма, зерно, деконтаминация, Aspergillus flavus, компактин, Rhodococcus erythropolus.
В последние годы в собранных на территории Российской Федерации образцах фуражного зерна, силоса и других кормов, загрязненных микотоксинами (1-3), все чаще обнаруживают афлатоксин В1 (АФВ1) (4, 5), в том числе в количествах, превышающих его предельно допустимые концентрации (2, 6, 7). Поскольку глобальное потепление способствует миграции теплолюбивых микромицетов в зоны с умеренным и холодным климатом, следует ожидать расширения ареала Aspergillus flavus и других АФВ1-образующих видов аспергиллов в этих регионах и, соответственно, повышения вероятности афлатоксинового загрязнения травостоя и фуражного зерна культивируемых там злаков.
Как и другие микотоксины, АФВ1 включается в пищевые цепи и может попадать в продукты питания, создавая серьезную угрозу здоровью людей из-за высокой гепатотоксичности и гепатокарциногенной активности (8). Поступая с кормом в организм животных, АФВ1 метаболизируется с образованием повреждающего ДНК экзо-8,9-эпоксида и афлатоксина М1 (9, 10), который накапливается в яйцах домашней птицы, молоке коров и обладает, как и сам АФВ1, канцерогенным, тератогенным, мутагенным и иммуносупрессивным действием (11).
В связи с высокой опасностью АФВ1 разработка эффективных методов снижения загрязненности им фуражного зерна, используемого в различных отраслях животноводства, представляет собой одну из наиболее важных задач обеспечения пищевой безопасности сельскохозяйственных кормов. Для ее решения исследователи привлекают как физические и химические, так и биологические методы деконтаминации (12, 13). Среди последних особое место занимает биодеградация — подход, основанный на разрушении или детоксифицирующей трансформации АФВ1 микроорганизмами-деструкторами (14, 15), которые секретируют метаболиты, обладающие соответствующей ферментативной активностью (16-18).
Способность к биодеградации афлатоксинов обнаружена у многих микроорганизмов, в том числе у актинобактерий рода Rhodococcus(19-22). Тем не менее изучение деструкционного потенциала этой таксономической группы в отношении AФB1 с целью выявления наиболее эффективных штаммов по-прежнему остается актуальным, поскольку свойство продуцировать афлатоксин-деградирующие метаболиты не относится к видоспецифичным. Как виды Rhodococcus, так и штаммы в пределах одного вида значительно различаются по продуктивности (20, 22). Кроме того, практически во всех проведенных исследованиях сообщалось только о способности этих актинобактерий деградировать АФВ1 при добавлении его в питательную среду и лишь в очень редких случаях — о возможности использовать бактерии для деградации AФB1, извлеченного из контаминированного зерна или отходов его переработки (23).
Содержание AФB1 в зерне, загрязненном его продуцентами, может быть снижено и с помощью синтетических или биологических ингибиторов афлатоксигенеза (12), обработка которыми будет препятствовать накоплению токсина в инфицированном субстрате. Перспективность такого подхода была продемонстрирована нами в предыдущих исследованиях, показавших, что компактин — один из вторичных метаболитов Penicillium citrinum способен подавлять образование AФB1 у Aspergillus flavus (24). Однако при повышении температуры и влажности в зернохранилищах нередко создаются условия, благоприятные для быстрого и интенсивного развития аспергиллов, что может существенно снизить результативность использования ингибиторов из-за недостаточно полного подавления токсигенеза. Чтобы обеспечить необходимую деконтаминацию, остатки токсина в обработанном ингибиторами зерне могут быть устранены посредством биодеградации.
В настоящей работе впервые было показано, что обработка супернатантом культуральной жидкости Rhodococcus erythropolusАС-884 вслед за обработкой компактином практически полностью предотвращает накопле-ние микотоксина в зараженном зерне.
Цель выполненной работы — оценка эффективности снижения содержания микотоксина в зерне пшеницы, искусственно контаминированном афлатоксином В1, после обработки актинобактериями рода Rhodococcus или ингибитором афлатоксигенеза (компактином), а также при сочетании этих методов.
Методика. Штаммы Rhodococcus sp. AC-1260, R. ruber AC1801, R. erythropolus AC-1269 и АС-884 (Всероссийская коллекция промышленных микроорганизмов, ФГУП Государственный НИИ генетики и селекции промышленных микроорганизмов НИЦ Курчатовский институт, г. Москва) культивировали на среде R1 (г/л): пептон — 15,0, дрожжевой экстракт — 6,0, NaCl — 1,0, глюкоза — 1,0, рН 7,0, которую используют при тестировании биологической активности актинобактерий рода Rhodococcus (25). В колбы объемом 50 мл с 10 мл питательной среды вносили равные количества бактериального инокулюма с концентрацией 109 клеток/мл и выращивали 48 ч на орбитальной качалке Excella® E-25R («New Brunswick», США) при температуре 30 °С и 290 об/мин. Одновременно в тех же условиях выдерживали колбы с питательными средами без актинобактерий (контроль). Количество образовавшейся бактериальной биомассы (число клеток/мл) определяли в конце культивирования.
Штамм PG-41 гриба Phoma glomerata — обнаруженного нами ранее биодеструктора AФB1 (26), токсин-деградирующую активность метаболитов которого сравнивали с активностью R. erythropolus АС-884, был получен из Государственной коллекции фитопатогенных микроорганизмов Всероссийского НИИ фитопатологии.
Для оценки способности вышеуказанных коллекционных штаммов актинобактерий продуцировать метаболиты, деградирующие АФВ1, бактериальные культуры центрифугировали 30 мин при 10000 g (5702 R, «Ep-pendorf AG», Германия). Перед добавлением токсина рН полученных супернатантов доводили до 7,0, чтобы предотвратить неэнзиматическое обратимое раскрытие лактонного кольца АФВ1, происходящее в щелочной среде при высоких значениях рН (27), которое может приводить к ложноположительным результатам анализа методом высокоэффективной жидкостной хроматографии (ВЭЖХ). Супернатанты фильтровали через мембраны (0,22 мкм, «Millipore», США). В освобожденные от бактериальных клеток супернатанты, а также в супернатанты контрольных сред добавляли АФВ1 («Sigma-Aldrich», США) до концентрации 0,2 мкг/мл, инкубировали смесь при 30 °С, постоянно перемешивая ее на качалке в течение 24 ч при указанных выше условиях, и определяли в смеси содержание токсина.
В опытах по деградации АФВ1 в искусственно загрязненном зерне семена мягкой яровой пшеницы (Triticum aestivum L.) сорта Дарья, помещенные в качалочные колбы объемом 250 мл (20 г на колбу), замачивали в воде (10 мл на 20 г зерна) и автоклавировали 1 ч при 1 атм. После стерилизации в колбы вносили по 1 мл сток-растворов АФВ1 в 20 % этаноле до конечных концентраций 1,0; 2,5 и 5,0 мкг/г зерна и интенсивно встряхивали в течение 10-15 мин, чтобы равномерно распределить токсин. В контрольные колбы добавляли по 1 мл 20 % этанола. Культуры Rhodococcus выращивали и центрифугировали, как указано выше, супернатанты отделяли, осадки суспендировали в 20 мл стерильной воды. Затем в колбы c зерном вносили выравненные по концентрации суспензии клеток тестируемых бактериальных штаммов из расчета 0,05 мл суспензии на 1 г зерна или 0,25 мл супернатантов. В контроль добавляли те же объемы стерильной воды. При сравнении токсин-деградирующей активности суспензии клеток и супернатанта КЖ R. erythropolus АС-884 супернатантом обрабатывали зерно, загрязненное максимальной из использованных концентраций АФВ1 (5,0 мкг/г).
В экспериментах с деконтаминацией зерна, зараженного токсигенным А. flavus,колбы со стерильными зернами инфицировали штаммом А11 (107 конидий/мл, 1 мл суспензии/20 г зерна) и распределяли на четыре партии. В первую партию колб вносили компактин до конечной концентрации 0,05 мг/г. Во второй партии зерно через 6 сут после инокуляции конидиальной суспензией той же концентрации обрабатывали супернатантом культуральной жидкости (КЖ) штамма AC-884 (0,25 мл/г). Зерно в третьей партии колб сначала обрабатывали компактином (0,05 мг/г), а через 6 сут вносили указанное выше количество супернатанта. В контрольные колбы добавляли 20 % этанол, стерильную воду или и этанол, и воду.
Колбы с зерном, искусственно контаминированным токсином, выдерживали в термостате ТСО-1/80 (ОАО «Смоленское СКТБ СПУ», Россия) при 30 °С в течение 3 сут после добавления биодеструкторов, а колбы с зараженным зерном — при 27 °С в течение 7 сут после инокуляции. Контрольные колбы инкубировали 3 или 7 сут в тех же условиях.
Споры токсигенного A. flavus А11 для инокуляции зерна пшеницы получали, как описано ранее (24). Компактин получали из КЖ P. citrinum, (штамм 18-12), используя метод, изложенный S.N. Ukraintseva с соавт. (28).
Культуры гриба P. glomerata PG-41, секретирующего АФВ1-деградирующие метаболиты, выращивали в жидкой питательной среде, как описано ранее (26), а супернатанты получали из фильтрата КЖ, центрифугируя и стерилизуя его так же, как в опытах с бактериальными культурами. Супернатанты P. glomerata использовали в опытах по деградации АФВ1 и деконтаминации зерна, зараженного A. flavus А11, аналогично описанному выше протоколу для бактериальных штаммов.
Остаточные количества АФВ1, извлеченного из супернатантов КЖ и зерна пшеницы с помощью экстракции хлороформом, определяли методом высокоэффективной жидкостной хроматографии (Waters 1525 Breeze HPLC SYSTEM, «Waters Corp.», США) (26, 28). Предел обнаружения АФВ1 составлял 0,005 мкг/г зерна (ПДК в зерне 0,01-0,5 мкг/г), полнота извлечения из КЖ — не менее 80 %.
Эксперименты (не менее 3 повторностей на вариант в каждом) повторяли трижды. Статистическую обработку проводили в программе STATISTIСA 6.1 («StatSoft, Inc.», США). Рассчитывали значения среднего (M) и стандартного отклонения (±SD). Достоверность различий между контролем и опытными вариантами при p ≤ 0,05 определяли, применяя t-тест для независимых переменных.
Результаты. Все исследованные штаммы Rhodococcus активно росли на среде R1. Их биомасса на 2 сут достигала 1010 бактериальных клеток/мл. Значения рН КЖ к концу культивирования у разных штаммов практически не различались (рН от 8,8 до 9,0). Эти результаты свидетельствуют о том, что культивирование потенциальных биодеструкторов АФВ1 в наших экспериментах было проведено в условиях, которые обеспечивали корректную сравнительную оценку их способности продуцировать метаболиты с целевой активностью и не могли препятствовать реализации этой способности у того или иного штамма.
Анализ остаточных количеств АФВ1 после его инкубации в супернатантах КЖ показал, что среди протестированных культур Rhodococcus наиболее активным продуцентом токсин-деградирующих метаболитов был штамм R. erythropolus AC-884. После инкубации АФВ1 в супернатанте КЖ этого штамма при нейтральном рН в течение 24 ч были обнаружены лишь следовые количества токсина (рис. 1). В супернатантах КЖ трех других штаммов при тех же условиях сохранялось от 15 до 50 % добавленного АФВ1 (см. рис. 1).
При анализе зерна, в которое был внесен коммерческий АФВ1, статистически значимое снижение его содержания по сравнению с контролем (при p ≤ 0,05) было обнаружено, только если обработку проводили, используя штамм AC-884. Через 3 сут после внесения бактериальной суспензии количество АФВ1 в зерне снижалось по сравнению с контролем почти в 2 раза (эффективность деградации 47,6 %), если конечная концентрация токсина составляла 1,0 мкг/г зерна. Статистически значимые различия с контролем после обработки клеточной суспензией (p = 0,003) сохранялись и при концентрации АФВ1 2,5 мкг/г. В то же время обработка супернатантом КЖ AC-884 зерна, контаминированного максимальной из использованных доз токсина (5,0 мкг/г), приводила к удалению в течение 3 сут от 60 до 70 % АФВ1 (рис. 2). Кроме того, деградация АФВ1 в супернатанте КЖ R. erythropolus AC-884 происходила быстрее, чем в аналогично полученном экстрацеллюлярном супернатанте культуры гриба P. glomerata PG-41 — другого биодеструктора АФВ1, который активно удалял токсин из модельных инкубационных сред (26), но в отличие от супернатанта КЖ AC-884 оказался малоэффективен при использовании на загрязненном зерне (рис. 3).
Высокая токсин-деградирующая активность метаболитов, секретируемых штаммом AC-884 в КЖ, была подтверждена экспериментами с инфицированием зерен пшеницы продуцирующим АФВ1 штаммом А. flavus (табл.). Через 1 сут после обработки супернатантом образцов зерна, на котором гриб развивался в течение 6 сут, содержание токсина в экстрактах, полученных из этих образцов, сокращалось почти в 3 раза по сравнению с контролем, а использование ингибитора приводило к 5-кратному уменьше-нию загрязненности.
В результате последовательного применения компактина и супернатанта КЖ AC-884 концентрация АФВ1 в экстрактах снижалась более чем в 200 раз (см. табл.), то есть комбинирование двух обработок обеспечивало почти 100 % деконтаминацию зерна. Поскольку в этих опытах применяли супернатант, освобожденный от бактериальных клеток, снижение содержания АФВ1 в зерне происходило в результате действия метаболитов, выделяемых бактериями в КЖ, а не из-за часто встречающейся адсорбции токсина клетками биодеструктора (29), которая может быть обратимой (30), из-за чего такой способ деконтаминации не всегда сохраняет свою эффективность (31).
Результаты наших предыдущих исследований (26), чувствительность обнаруженной АФВ1-деградирующей активности к протеолизу и анализ данных литературы позволяют предполагать, что активность супернатанов AC-884 имеет энзиматическую природу. Так, ферменты, которые катализируют деградацию токсинов грибов, например алкалоидов спорыньи (гидролаза ErgA, амидаза ErgB) (32), или участвуют в катаболизме токсичных полиароматических соединений (например, бифенилдиоксигеназы и дегидрогеназы), идентифицированы у целого ряда штаммов R. erythropolus. Установлено, что гены, кодирующие эти ферменты, сгруппированы в кластеры, а энзиматическая деградация происходит через каскад реакций, включающих расщепление ароматических колец. Поскольку AФB1 — это также полиароматическое соединение, предполагается, что он может быть мишенью подобных ферментов и разлагаться аналогичным образом (33).
Высокий деконтаминирующий потенциал супернатанта КЖ AC-884 в наших опытах согласуются с полученными ранее данными о способности экстрацеллюлярных супернатантов других шаммов R. erythropolus деградировать АФВ1, значительно снижая при этом его гентоксичность (33). По-видимому, синтез АФВ1-деградирующих ферментов относится к конститутивным свойствам многих штаммов R. erythropolus, хотя, в отличие от AC-884, не все они способны секретировать такие фермены КЖ (22).
В наших исселдованиях использование ингибитора биосинтеза АФВ1 совместно с последующим удалением остаточных количеств токсина с помощью их биодеградации приводило к практически полной деконтаминации загрязненного зерна. Возможность практической реализации такого подхода будет проверена в дальнейших экспериментах посредством обработки образцов закладываемого на хранение зерна, собранного в зонах риска загрязнения продуцентами афлатоксинов, и контаминированного силоса. Весьма перспективно и то, что некоторые штаммы R. erythropolus способны разлагать микотоксины Т-2 (20) и зеараленон (18), а также использовать охратоксин А в качестве источника фенилаланина (34). Поскольку недавно мы обнаружили, что аналог компактина может тормозить образование зеараленона (35), представляется целесообразной проверка способности AC-884 деградировать этот фузариотоксин и другие микотоксины. В случае обнаружения такой способности сфера применения разрабатываемого подхода может быть существенно расширена. Доступность источников целевых метаболитов, а именно афлатоксин-деградирующая активность в супернатанте КЖ R. erythropolus AC-884 и способность P. citrinum 18-12 к секреции компактина in vitro (26), служит фактором, который в будущем может способствовать разработке относительно несложной технологии получения двухкомпонентного биопрепарата для деконтаминации фуражного зерна и других кормов.
Таким образом, в результате исследования токсин-деградирующей способности четырех штаммов Rhodococcus (Rhodococcus sp. AC-1260, R. ruber AC1801, R. erythropolus AC-1269 и АС-884) и ее сравнения в опытах с обработкой зерна супернатантом культуральной жидкости (КЖ) Phomaglomerata PG-41 был отобран наиболее активный биодеструктор афлатоксина В1 (АФВ1) — штамм R. erythropolus AC-884. Эффективность деконтаминации зерна, искусственно загрязненного препаратом токсина, после обработки супернатантом культуральной жидкости AC-884 могла достигать 70 %, а после применения на зерне, зараженном токсигенным Aspergillus flavus, в среднем составляла 65 %. Обработка такого зерна компактином снижала содержание АФВ1 с эффективностью почти 79 %, комбинированная обработка, то есть внесение вслед за компактином супернатанта КЖ AC-884, фактически полностью предотвращала накопление микотоксина в зараженном A. flavus зерне пшеницы.
Накопление афлатоксина В1 (АФВ1) в зараженном Aspergillus flavus А11 зерне пшеницы (Triticum aestivum L.) сорта Дарьяпосле раздельной или совместной обработки зерна компактином и метаболитами Rhodococcus erythropolus AC-884 (n = 9, M±SD) |
||
Вариант |
Содержание АФВ1, мкг/мл |
Эффективность деконтаминации, % |
Компактин, 0,05 мг/г зерна |
7,45±1,34 |
78,5 |
Супернатант культуральной жидкости (КЖ) AC-884, 0,25 мл/г зерна |
12,10±1,27 |
65,1 |
Компактин, 0,05 мг + супернатант КЖ AC-884, 0,25 мл/г зерна |
0,13±0,09 |
99,6 |
Контроль (без обработки) |
34,66±3,07 |
|
Примечание. Отличия от контроля и между обработками статистически значимы при р < 0,05. |
||
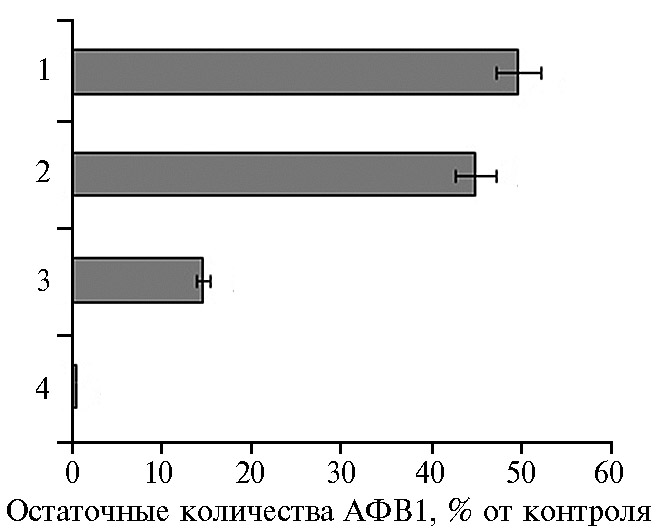
Рис. 1. Способность различных штаммов Rhodococcus spp. снижать содержание афлатоксина В1 (АФВ1), добавленного в супернатант их культуральной жидкости. Средние значения для трех экспериментов: 1 — R. ruber AC1801, 2 — Rhodococcus sp. AC-1260, 3 — R. erythropolus AC-1269, 4 — R. erythropolus АС-884 (n = 9, M±SD).
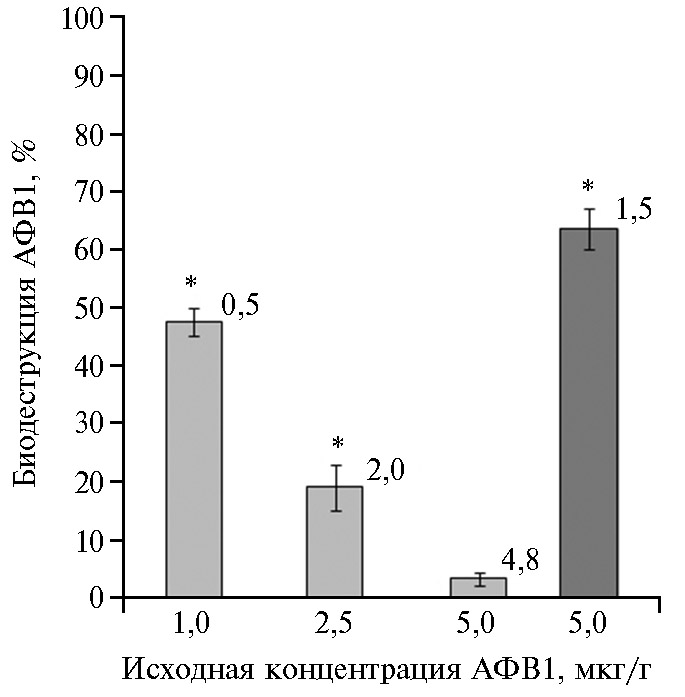
Рис. 2. Эффективность деградации афлатоксина В1 (АФВ1), добавленного в зерно пшеницы (Triticum aestivum L.) сорта Дарья, с помощью суспензии клеток (светло-серые столбцы) или супернатанта культуральной жидкости (темно-серый столбец) Rhodococcus erythropolus AC-884 в зависимости от исходной концентрации токсина (n = 9, M±SD). Цифры рядом со столбцами показывают остаточные количества АФВ1 в зерне (мкг/г) через 3 сут после обработки.
*Отличия от контроля статистически значимы при р ≤ 0,05 (отмечены звездочкой).
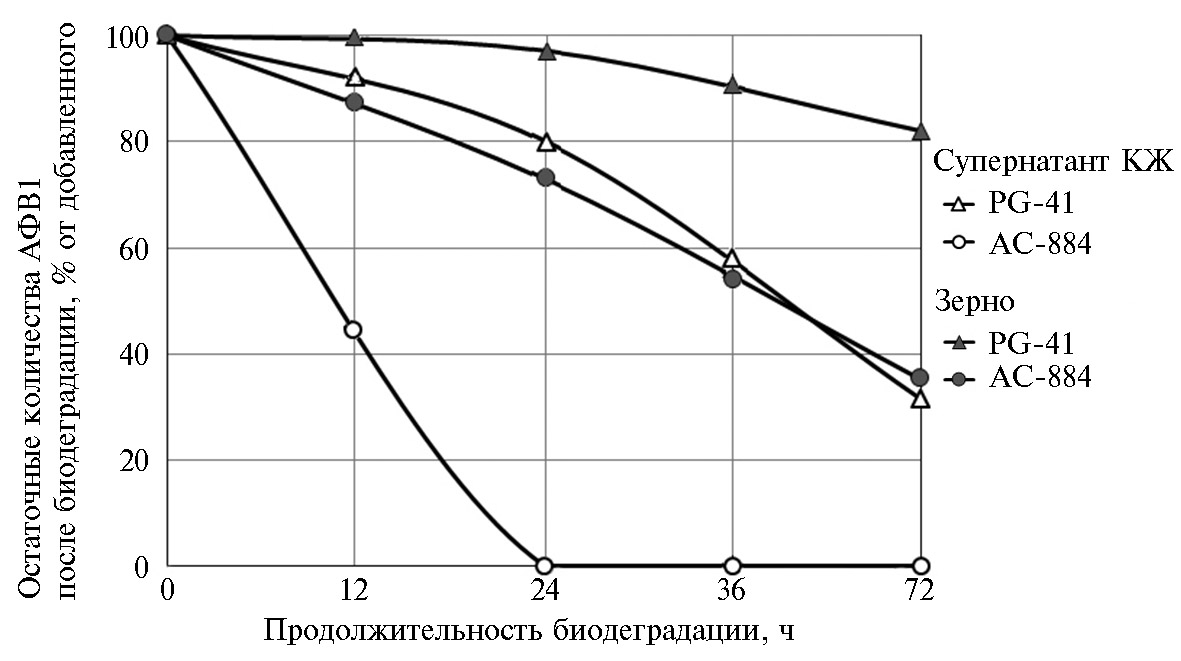
Рис. 3. Динамика деградации афлатоксина В1 (АФВ1) в течение 3 сут в супернатанте культуральной жидкости Rhodococcus erythropolus AC-884 или Phoma glomerata PG41 и в обработанном ими зерне пшеницы (Triticum aestivum L.) сорта Дарья, искусственно загрязненном токсином.
ЛИТЕРАТУРА
- Борутова Р., Айдинян Г. Микотоксины: беспечность недопустима. Исследования образцов кормового сырья растительного происхождения (урожай 2020 г.). Животноводство России, 2021, 4: 54-59.
- Йылдырым Е.А., Ильина Л.А., Филиппова В.А., Солдатова В.В., Никонов И.Н., Лаптев Г.Ю., Соколова О.В., Новикова Н.И. Изучение распространения микотоксинов в силосе и разработка стратегии борьбы с ними. Кормопроизводство, 2016, 3: 41-45.
- Танасева С.А., Босяков И.В. Мониторинг афлатоксинов в кормах из Рязанской области. Мат. межд. науч.-практ. конф., посвященной 90-летию со дня рождения профессора В.А. Киршина. Казань, 2018: 191-193.
- Седова И.Б., Захарова Л.П., Киселева М.Г., Чалый З.А., Тутельян В.А. Фузариотоксины и афлатоксин В1 в продовольственном зерне кукурузы в Российской Федерации. Научные труды СКФНЦСВВ, 2018, 21: 129-137.
- Мирошниченко П.В., Панфилкина Е.В., Околот М.В. Мониторинговые исследования содержания микотоксинов в кормах для крупного рогатого скота Краснодарского края. Сборник научных трудов ФГБНУ КНЦЗВ, 2020, 9(2): 109-111 (doi: 10.34617/85s8-np65).
- Коваленко А., Солдатенко Н., Фетисов Л., Сухих Е. Микотоксикологический монторинг кормов в Северо-Кавказском регионе. Комбикорма, 2011, 3: 98-99.
- Семёнов Э.И., Папуниди К.Х., Тремасов М.Я. Микотоксикозы в АПК: распространение, диагностика, профилактика. Режим доступа: http://soyanews.info/news/mikotoksikozy_v_apk-_rasprostranenie-_diagnostika-_profilaktika.html. Дата обращения: 14.10.2021.
- Aflatoxins — biochemistry and molecular biology /R.G. Guevara-González (ed.). InTech, London, 2000 (doi: 10.5772/896).
- Molecular and applied aspects of oxidative drug metabolizing enzymes /E. Arinç, J.B. Schenkman, E. Hodgson (eds.). Springer, New York, 1999 (doi: 10.1007/978-1-4615-4855-3).
- Yang C., Song G., Lim W. Effects of mycotoxin-contaminated feed on farm animals. Journal of Hazardous Materials,2020, 389: 122087 (doi: 10.1016/j.jhazmat.2020.122087).
- Min L., Fink-Gremmels J., Li D., Tong X., Tang J., Nan X., Yu Z., Chen W., Wang G. An overview of aflatoxin B1 biotransformation and aflatoxin M1 secretion in lactating dairy cows. Animal Nutrition, 2020, 7(1): 42-48 (doi: 10.1016/j.aninu.2020.11.002).
- Джавахия В.Г., Стацюк Н.В., Щербакова Л.А., Поплетаева С.Б. Афлатоксины: ингибирование биосинтеза, профилактика загрязнения и деконтаминация агропродукции. М., 2017.
- Sipos P., Peles F., Brassó D.L., Béri B., Pusztahelyi T., Pócsi I., Győri Z. Physical and chemical methods for reduction in aflatoxin content of feed and food. Toxins, 2021, 13(3): 204 (doi: 10.3390/toxins13030204).
- Lyagin I., Efremenko E. Enzymes for detoxification of various mycotoxins: origins and mechanisms of catalytic action. Molecules, 2019, 24(13): 2362 (doi: 10.3390/molecules24132362).
- Loi M., Fanelli F., Zucca P., Liuzzi V.C., Quintieri L., Cimmarusti M.T., Monaci L., Haidukowski M., Logrieco A.F., Sanjust E., Mulè G. Aflatoxin B1 and M1 degradation by Lac2 from Pleurotus pulmonarius and redox mediators. Toxins, 2016, 8(9): 245 (doi: 10.3390/toxins8090245).
- Wang J., Ogata M., Hirai H., Kawagishi H. Detoxification of aflatoxin B1 by manganese peroxidase from the white-rot fungus Phanerochaete sordida YK-624. FEMS Microbiology Letters, 2011, 314(2): 164-169 (doi: 10.1111/j.1574-6968.2010.02158.x).
- Cao H., Liu D., Mo X., Xie C., Yao D. A fungal enzyme with the ability of aflatoxin B1 conversion: purification and ESI-MS/MS identification. Microbiological Research, 2011, 166(6): 475-483 (doi: 10.1016/j.micres. 2010.09.002).
- Wu Y.Z., Lu F.P., Jiang H.L., Tan C.P., Yao D.S., Xie C.F., Liu D.L. The furofuran-ring selectivity, hydrogen peroxide-production and low Km value are the three elements for highly effective detoxification of aflatoxin oxidase. Food and Chemical Toxicology, 2015, 76: 125-131 (doi: 10.1016/j.fct.2014.12.004).
- Krifaton C., Kriszt B., Szoboszlay S., Cserháti M., Szűcs A., Kukolya J. Analysis of aflatoxin-B1-degrading microbes by use of a combined toxicity-profiling method. Mutation Research, 2011, 726(1): 1-7 (doi: 10.1016/j.mrgentox.2011.07.011).
- Cserháti M., Kriszt B., Krifaton C., Szoboszlay S., Háhn J., Tóth S., Nagy I., Kukolya J. Mycotoxin-degradation profile of Rhodococcus strains. International Journal of Food Microbiology, 2013, 166(1): 176-185 (doi: 10.1016/j.ijfoodmicro.2013.06.002).
- Eshelli M., Harvey L., Edrada-Ebel R., McNeil B. Metabolomics of the bio-degradation process of aflatoxin B1 by Actinomycetes at an initial pH of 6.0. Toxins, 2015, 7(2): 439-456 (doi: 10.3390/toxins7020439).
- Risa A., Divinyi D.M., Baka E., Krifaton C. Aflatoxin B1 detoxification by cell-free extracts of Rhodococcus strains. Acta Microbiologica et Immunologica Hungarica, 2017, 64(4): 423-438 (doi: 10.1556/030.64.2017.023).
- Prettl Z., Dési E., Lepossa A., Kriszt B., Kukolya J., Nagy E. Biological degradation of aflatoxin B1 by a Rhodococcus pyridinivorans strain in by-product of bioethanol. Animal Feed Science and Technology, 2017, 224: 104-114 (doi: 10.1016/J.ANIFEEDSCI.2016.12.011).
- Dzhavakhiya V.G., Voinova T.M., Popletaeva S.B., Statsyuk N.V., Limantseva L.A., Shcherbakova L.A. Effect of various compounds blocking the colony pigmentation on the aflatoxin B1 production by Aspergillus flavus. Toxins, 2016, 8(11): 313 (doi: 10.3390/toxins8110313).
- Дудчик Н.В., Дроздова Е.В., Трейлиб В.В., Будкина Е.А., Бурая В.В., Козлова Т.О., Ушкова Л.Л. Оценка интегральной токсичности объектов окружающей среды методами биотестирования (инструкция по применению). Минск, 2012.
- Shcherbakova L.A., Statsyuk N.V., Mikityuk O.D., Nazarova T.A., Dzhavakhiya V.G. Aflatoxin B1 degradation by metabolites of Phoma glomerata PG41 isolated from natural substrate colonized by aflatoxigenic Aspergillus flavus. Jundishapur Journal of Microbiology, 2015, 8(1): e24324 (doi: 10.5812/jjm.24324).
- Vankayalapati V.K. Aflatoxins: properties, toxicity and detoxification. Nutrition and Food Science International Journal, 2018, 6(5): 555696 (doi: 10.19080/NFSIJ.2018.06.555696).
- Ukraintseva S.N., Voinova T.M., Dzhavakhiya V.G. Obtaining the highly productive mutants Penicillium citrinum producing compactin and optimization of fermentation process in shaken flasks. In: Biotechnology in biology and medicine /A.M. Egorov, G. Zaikov (eds.). Nova Science Publishers, New York, 2006.
- Bueno D.J., Casale C.H., Pizzolitto R.P., Salvano M.A., Oliver G. Physical adsorption of aflatoxin B1 by lactic acid bacteria and Saccharomyces cerevisiae: a theoretical model. Journal of Food Protection, 2007, 70(9): 2148-2154 (doi: 10.4315/0362-028X-70.9.2148).
- Peltonen K., El-Nezami H., Haskard C., Ahokas J., Salminen S. Aflatoxin B1 binding by dairy strains of lactic acid bacteria and bifidobacteria. Journal of Dairy Science, 2001, 84(10): 2152-2156 (doi: 10.3168/jds.S0022-0302(01)74660-7).
- Ondiek W., Wang Y., Sun L., Zhou L., On S.L., Zheng H., Ravi G. Removal of aflatoxin b1 and t-2 toxin by bacteria isolated from commercially available probiotic dairy foods. Food Science and Technology International, 2021, 28(1): 15-25 (doi: 10.1177/1082013220987916).
- Hahn I., Thamhesl M., Apfelthaler E., Klingenbrunner V., Hametner C., Krska R., Schatzmayr G., Moll W.-D., Berthiller F., Schwartz-Zimmermann H.E. Characterisation and determination of metabolites formed by microbial and enzymatic degradation of ergot alkaloids. World Mycotoxin Journal, 2015, 8(4): 393-404 (doi: 10.3920/WMJ2014.1807).
- Alberts J.F., Engelbrecht Y., Steyn P.S., Holzapfel W.H., van Zyl W.H. Biological degradation of aflatoxin B1 by Rhodococcus erythropolis cultures. International Journal of Food Microbiology, 2006, 109(1-2): 121-126 (doi: 10.1016/j.ijfoodmicro.2006.01.019).
- Rodriguez H., Reveron I., Doria F., Costantini A., De Las Rivas B., Munoz R., Garcia-Moruno E. Degradation of ochratoxin A by Brevibacterium species. Journal of Agricultural and Food Chemistry, 2011, 59(19): 10755-10760 (doi: 10.1021/jf203061p).
- Микитюк О.Д., Воинова Т.М., Джавахия В.Г. Подавление образования фузариотоксина зеараленона 6-деметилмевинолином — ингибитором биосинтеза афлатоксина В1. Мат. межд. науч. конф. «Актуальные вопросы органической химии и биотехнологии». Екатеринбург, 2020, т. 1: 659-660.