doi: 10.15389/agrobiology.2022.1.27rus
УДК 633.39:581.144.2
Исследование выполнено при поддержке гранта Российского научного фонда (проект № 19-16-00114).
ОСОБЕННОСТИ ФОРМИРОВАНИЯ КОРНЕВОЙ СИСТЕМЫ У НЕКОТОРЫХ КОРМОВЫХ ПОЛУКУСТАРНИКОВЫХ И КУСТАРНИКОВЫХ ГАЛОФИТОВ В ПРЕДГОРНОЙ ПУСТЫНЕ УЗБЕКИСТАНА
Э.З. ШАМСУТДИНОВА1 ✉, Н.З. ШАМСУТДИНОВ2, И.В. САВЧЕНКО3, О.А. СТАРШИНОВА1, В.И. АГАФОНОВ1,
З.Ш. ШАМСУТДИНОВ1
Отличительные особенности галофитов как кормовых растений — высокая питательная ценность, стабильная сбалансированность корма по питательным веществам по сезонам года, особенно в критические периоды пастбищного содержания животных (осенью и зимой), достаточно высокое содержание незаменимых аминокислот. Кормовые галофитные полукустарники (кохия простертая, солянка восточная), кустарники (саксаул черный, солянка малолистная) в условиях ксеротермической среды среднеазиатской пустыни характеризуются высокой и устойчивой кормовой продуктивностью. Их способность успешно осуществлять полный жизненный цикл в этих условиях реализуется за счет структурных и физиолого-биологических приспособлений. Корневая система играет центральную роль в формировании урожая у культурных растений и в настоящее время рассматриваются как ключевой фактор второй «зеленой революции». Знание характера формирования корневой системы кустарников и полукустарников семейства Маревые (Chenopodiaceae) в условиях предгорной пустыни, в частности скорости роста, глубины проникновения в почву, очень важно не только для познания этих галофитных жизненных форм растений, но и для правильного размещения посевов в аридных зонах. Мы сравнили особенности формирования корневой системы у кустарниковых галофитов — саксаула черного Haloxylon aphyllum (Minkw.) Iljin, солянки малолистной Aellenia subaphylla (С.А. Меу) Aellen и полукустарниковых галофитов — солянки восточной Salsola orientalis S.G. Gmel., кохии простертой Kochia prostrata (L.) Schrad., чтобы выявить их экологическую роль в условиях зоны среднеазиатской предгорной пустыни (Нишанская степь, Кашкадарьинская область, Республика Узбекистан, 2015-2020 годы) у растений 1-го и 5-го года жизни. В процессе эволюции и естественного отбора, формирования более совершенных адаптивных свойств и повышенных продукционных функций у галофитов семейства Маревые важное значение имело развитие мощной и глубоко проникающей в почву корневой системы универсального типа. Показано, что для кохии простертой, солянки восточной, саксаула черного и солянки малолистной характерна высокая скорость роста и развития корневой системы. В 1-й год жизни корневые системы кустарников саксаула черного проникают в почву на глубину до 235 см, солянки малолистной — до 150 см, полукустарников солянки восточной — до 200 см, кохии простертой — до 215-295 см, достигая в 5-летнем возрасте у саксаула черного 1240 см, солянки малолистной — 600 см, солянки восточной — 550 см, кохии простертой — 580 см, что превышает в 1-й год высоту надземной части в 4-4,5 раза, в 5-летнем возрасте — в 6 раз. Способностью к быстрому росту и формированию специфической структуры корневой системы обеспечивается постоянство поглощения воды корнями в условиях засухи и дефицита влаги. На глубину проникновения корневой системы растений разных жизненных форм (кустарники, полукустарники) сильное влияние оказывают водно-физические свойства эдафической среды. В условиях постоянного дефицита почвенной влаги корневая система стремится постоянно углубляться в почвенно-грунтовую среду, пробивая плотные, сцементированные слои почвы. По нашему мнению, полукустарниковым и кустарниковым галофитам присуща способность выполнять функцию так называемого гидравлического лифта, когда растение не только поднимает воду корнями, но и выделяет ее в более сухие горизонты почвы, где этой водой могут воспользоваться растения с неглубокой корневой системой.
Ключевые слова: кормовые галофиты,кустарники, полукустарники, Salsola orientalis S.G. Gmel., кейреук, Kochia prostrata L., кохия простертая, Aellenia subaphylla (С.А. Меу) Aellen, солянка малолистная, Haloxylon aphyllum (Minkw.) Iljin, саксаул черный, корневая система, морфология, гидравлический лифт.
Отличительные особенности галофитов как кормовых растений —высокая питательная ценность, стабильная сбалансированность корма по питательным веществам по сезонам года, особенно в критические периоды пастбищного содержания животных (осенью и зимой), достаточно высокое содержание незаменимых аминокислот(1). В разных районах мира отмечали значение кормовых кустарников и полукустарников как источников протеина. Так, содержание сырого протеина у Kochia brevifolia R. Br. в Западной Австралии составляет 20 %, у Atriplex nummularia L. и Atriplexvesicaria Heward ex Benth. — 14-18 % (2). Важная биологическая особенность кустарников (саксаул черный, солянка малолистная) и полукустарников (кохия простертая, солянка восточная) заключается в быстром росте и развитии в культуре (3, 4).
В процессе галоксерофилизации под влиянием возрастающей аридизации климата кустарники и полукустарники семейства Маревые подвергались глубочайшей приспособительной перестройке различных признаков и функций, в том числе физиолого-биохимических. Такая перестройка в ксеротермических условиях проявлялась прежде всего в редукции испаряющих вегетативных органов и усилении поглощающей (всасывающей) функции корневой системы (5, 6).
У кустарниковых и полукустарниковых галофитов в процессе совершенствования адаптивных свойств, обеспечивших им успешное функционирование и репродуцирование в жестких ксеротермических условиях среднеазиатской пустыни, очевидно, большое значение имеют глубоко проникающие в почву корневые системы (7). Известно, что корни очень важны для закрепления и поглощения водно-минеральных ресурсов, но не все исследователи рассматривают корни как важный орган, принимающий активное участие в формировании фитомассы (урожая) (8). Традиционно большинство исследователей основное внимание уделяли изучению надземной части растений (стебли, листья, цветки, плоды, семена) и упускали из виду корневую систему (9). В настоящее время многие исследователи начинают понимать, что корни растений играют центральную роль в формировании урожая у культурных растений. В обзоре V. Gewin (10), опубликованном в 2010 году в журнале «Nature», отмечается, что успех первой «зеленой революции» связывают с селекцией карликовых короткостебельных сортов пшеницы, у которых энергия и метаболиты в большей степени расходуются на формирование не стебля, а зерна. По прогнозам (10), ключевой фактор второй «зеленой революции» — корневая система, улучшение ее архитектоники и эколого-физиологических функций.
С углубленным пониманием вопросов, касающихся корневой системы растений, связывают решение практических задач в растениеводстве, в частности более эффективное использование удобрений и воды, а также обеспечение устойчивой продуктивности при различных биотических и абиотических стрессах (11). Засоление земель создает неблагоприятные условия для сельскохозяйственного производства, приводя к глобальным ежегодным потерям продукции на сумму, превышающую 12 млрд долларов США (12). В Китае засоленно-щелочные почвы составляют 25 % сельскохозяйственных угодий и используются недостаточно. Одна из устойчивых стратегий более эффективного вовлечения засоленных земель в сельскохозяйственное производство — разведение галофитов, которые могут выжить и завершить свой жизненный цикл в почвенных средах, содержащих более 200 мМ NaCl (13). Недавно в проведенных в Иране исследованиях установлено, что галофитные виды рода Suaeda sp. содержат в корнях азотфиксирующие эндофитные бактерии, которые могут внести существенный вклад в обеспечение растений азотом (14).
Изучение галофитов дополнительно актуализируется в условиях изменяющегося климата и необходимости обеспечивать продуктами питания растущее население Земли (15, 16).
По данным И.И. Судницына (17), скорость поглощения воды растением прямо пропорциональна глубине проникновения и плотности размещения корней в почвенном слое. Поэтому знание характера формирования корневой системы кустарников и полукустарников семейства Маревые (Chenopodiaceae) в условиях предгорной пустыни, в частности скорости роста, глубины проникновения в почву, очень важно не только для познания этих галофитных жизненных форм растений, но и для правильного размещения посевов в аридных зонах.
В представленном сообщении впервые рассматривается роль корневой системы в водоснабжении и поддержании водного баланса галофитных кустарников и полукустарников в ксеротермических условиях среднеазиатской пустыни.
Цель работы — сравнить особенности формирования корневой системы кустарниковых и полукустарниковых видов галофитов и выявить их экологическую роль в условиях среднеазиатской пустыни.
Методика. Исследование проводились в зоне предгорной пустыни (Нишанская степь, Кашкадарьинская область, Республика Узбекистан, 38.62624 с.ш., 65.69219 в.д.) в 2015-2020 годах в интродукционном питомнике кормовых кустарников и полукустарников, заложенном в 2015 году. Район раскопок корневых систем растений разных жизненных форм расположен в нижнем поясе предгорной пустыни на высоте 354 м над уровнем моря. Климатические условия района исследований характеризуются высокой сухостью и континентальностью климата.
В опытах с полукустарником кохией простертой исследовали три экотипа: каменистый (семена собраны в Ошской области Киргизии), песчаный (семена собраны в пустынях Кызылкум в Узбекистане, Муюнкум в Казахстане и в Прикаспийской полупустыне) и солонцовый (Ачикулакская научно-исследовательская лесная опытная станция, Россия).
Фенологические наблюдения по каждому исследуемому виду проводили на 75 растениях в трех повторностях.
Раскопку корневых систем кустарников (саксаул черный, солянка малолистная) и полукустарников (кохия простертая, солянка восточная) осуществляли траншейным методом (18). Раскопку корневых систем кустарников и полукустарников в возрасте 1 года проводили у трех растений каждого вида в разные фазы развития (всходы, настоящие листья, ветвление, цветение, плодообразование). Для раскопки корневых систем саксаула черного, солянки малолистной, солянки восточной, кохии простертой в 5-летнем возрасте выбирали растение, внешне соответствующее среднеразвитому представителю каждого из исследуемых видов.
Статистическую обработку полученных данных проводили в программе Microsoft Excel 2010. Результаты измерений представлены как средние (M) и их стандартные ошибки (±SEM).
Результаты. В зоне предгорной пустыни Кашкадарьинской области вегетационный сезон состоит из мезотермических и ксеротермических периодов, характеризующихся определенным режимом температуры и увлажнения (5). Мезотермический (прохладный и влажный) период приходится на ноябрь-апрель. В это время в районе исследований в год выпадает в среднем 224 мм осадков Ксеротермический (засушливый) период охватывает май-октябрь. Летом почва иссушается из-за физического испарения и транспирации растений под действием высоких температур, инсоляции и постоянно дующих ветров. Среднегодовая температура воздуха 14,8 °С , максимальная — +47°С, минимальная — -27°С. Переход температуры через 0 °С в среднем приходится на конец февраля-начало марта. Средняя температура воздуха в феврале 3,6 °С, в марте — 9,4 °С, в апреле — 15,7 °С. Относительная влажность в среднем за год — 30 %, летом — 10 %.
Почвенный покров, где были проведены раскопки корневых систем, как и весь нижний пояс предгорной пустыни, представлен в основном светлыми сероземами. Характерная особенность почвенного профиля — его слоистое строение (чередование горизонтов легкого, среднего и тяжелого суглинка, а также супеси).
Почвы в значительной степени засолены, их можно отнести к солончаковым. Не засолен только верхний 8-сантиметровый слой, ниже засоление слабое — 0,25-0,45 %, а на глубине 94-610 см содержание водорастворимых солей доходит до 1,35-2,77 %. Содержание гипса в почве этих разрезов небольшое и по горизонтам колеблется в пределах 0,5-35,8 %, гумуса в корнеобитаемом слое — 1,86-2,02 %, общего азота по горизонтам — 0,006-0,12 %, общего фосфора — 0,02-0,19 %. Калий присутствует в достаточном количестве во всем корнеобитаемом горизонте (в верхних слоях — 936 мг/кг, в нижних — 30 мг/кг).
Солянка восточная (Salsola orientalis S.G. Gmel.) — многолетнее растение высотой 40-60 см, имеет стволик высотой 5-10 см, от которого отходят 3-6 скелетных осей (19). Солянка восточная по жизненной форме полукустарник, по экологии — галоксерофит, характеризуется высокой выносливостью к засухе и устойчивостью к солевому стрессу (6). Способность солянки восточной успешно осуществлять полный жизненный цикл в ксеротермических аридных условиях при высокой засоленности почвы реализуется за счет структурных приспособлений и суккулентной организации листьев (20), многослойности эпидермиса, утолщения кутикулы (21) и С4-типа фотосинтеза, обеспечивающего более экономное и эффективное использование воды на транспирацию, чем у С3-растений (22, 23).
Данные, характеризующие рост корневой системы солянки восточной в первый год жизни, приведены в таблице 1. В конце апреля при высоте надземной части растения 0,9-1,0 см корни солянки восточной проникают на глубину 44±5,3 см, в конце мая углубляются до 80±4,1 см, в конце вегетации (2.XII.2016) — до 200±11,2 см.
Интересно отметить, что глубина проникновения корней солянки восточной больше высоты ее надземной части в 4,1-4,7 раза, а во второй половине вегетации — 2,5-3,5 раза. Корневой коэффициент (произведение максимальной глубины проникновения в почву корневой системы на максимальный диаметр ее распространения в сантиметрах) (24) составляет в первой половине вегетации 726-7600, а во второй — 10625-29000. Во второй и последующие годы жизни корневая система продолжает развиваться. По нашим наблюдениям, в мае корневая система солянки восточной 5-летнего возраста была мощной, проникающей в почву на глубину 550 см (рис. 1).
Главный корень на глубине 8 см делится на два крупных корня, идущих вниз под небольшим углом друг к другу. Один из них на глубине 12 см, другой — на глубине 17 см, в свою очередь, делятся на два, образуя четыре довольно крупных корня. Один из них на глубине 25 см сворачивает под углом 45° в сторону на 65 см, постепенно углубляясь в грунт. На глубине 140 см один из корней резко сворачивает в сторону (горизонтально примерно на 1 м в длину), затем снова направляется вниз и на глубине 330 см, войдя в плотный дресвяный горизонт, уходит в сторону горизонтально. Другой, самый тонкий из трех корней, направляется вертикально вниз, сильно ветвится на мелкие белые корешки. Третий корень на глубине 340 см, сделав петлю, идет немного вверх и в сторону на 30 см, затем резко поворачивает вниз. Главный корень солянки восточной, дойдя до плотного горизонта, сворачивает в сторону, делится на несколько корней, которые расходятся по сторонам, некоторые поднимаются даже вверх, затем разветвляются на многочисленные тонкие корни, которые, в свою очередь, делясь на более мелкие, заканчиваются в рыхлом среднесуглинистом влажном горизонте. Быстрый рост и формирование мощной корневой системы солянки восточной имеют решающее значение в рациональном использовании водно-минеральных ресурсов почвенно-грунтовой среды и формировании высокой кормовой и семенной продуктивности в засушливых условиях предгорной пустыни.
Кохия простертая [Kochia prostrata (L.) Schrad.] — многолетнее растение, по экологии ксерогалофит, по жизненной форме полукустарник из семейства Маревые, по нашим данным, имеет высоту 75-110 см, формирует 16-25 ц/га сухой кормовой массы, содержит 16-18 % протеина. Предназначен для выращивания без орошения в аридных районах со слабой и средней степенью засоления для создания долголетних высокопродуктивных пастбищ в засушливых районах. В опытах, проведенных в засушливых районах США (штаты Айдахо и Юта), кохия простертая оказалась лучшей по кормовой продуктивности, питательной ценности и переваримости полученного из нее корма (25-27).
Мы сравнили развитие растений кохии простертой разных экотипов в 1-й год жизни (табл. 2). Как оказалось, глубина проникновения корневой системы у растений в значительной мере изменяется в зависимости от экотипической принадлежности.
На среднесуглинистых светлых сероземах предгорной пустыни у кохии простертой к концу 1-го года вегетации формируется довольно глубоко (до 215-295 см) проникающая корневая система. В строении корневой системы между экотипами имелись следующие различия. Так, корневая система кохии простертой каменистого экотипа характеризуется ярко выраженным главным корнем. У песчаных экотипов корневые системы достаточно мощные и глубоко проникающие. Однако на глубине 80-100 см их главный корень по размеру и характеру развития становится похож на боковые корни 1-го порядка. Слабее всех развита корневая система у кохии простертой солонцового экотипа.
У 5-летней кохии простертой (каменистый экотип) корневая система в условиях Нишанской степи проникает на глубину 580 см (рис. 2). Главный корень вертикально направляется вниз. Диаметр корневой шейки 4 см. На глубине 10 см от главного корня отходит довольно крупный корень, идущий в горизонтальном направлении, он не углубляется в почву и сильно ветвится. Главный корень углубляется, образуя при этом небольшие изгибы и повороты. На глубине 50-75 см на главном корне появляется много мелких и несколько крупных боковых корней. Мелкие корни в этих слоях заканчиваются, а крупные направляются вниз. На глубине 75 см главный корень заметно утончается, на глубине 120-125 см — сильно ветвится и образует много тонких корней, направленных вниз. До глубины 300 см крупные боковые корни несут на себе большое число живых тонких корешков. Большинство корней, направляющихся вниз, на глубине 400 см проникают в плотный дресвяный горизонт, здесь сильно утончаются, распадаясь на большое число мелких, и в слое 450-470 см от поверхности заканчиваются, образуя густую сеть корневых волосков. Лишь один корень кохии простертой проходит дресвяный горизонт и заканчивается в рыхлом среднесуглинистом слое на глубине 580 см.
В морфологии корневой системы кохии простертой, произрастающей в условиях предгорной пустыни (Нишанская степь), можно выделить три яруса: первый — эфемерные корни, которые располагаются в слое 0-25-30 см; второй ярус находится на глубине 120-150 см в зоне сильного бокового ветвления корней, третий — в зоне окончания корневой системы, где корни сильно разветвляются и несут на себе много живых всасывающих корешков.
Таким образом, корневая система кохии простертой, культивируемой в предгорной пустыне, может быть охарактеризована как мощная и глубоко проникающая, способная утилизировать водно-минеральные ресурсы довольно большого объема почвенно-грунтовой среды.
Солянка малолистная [Aellenia subaphylla (С.А. Меу) Aellen] — многолетнее растение, галоксерофитный кустарник со стволиками, покрытыми светло-серой корой (19). Сильно ветвистое растение, с ветвями большей частью отклоненными, имеющими светло-зеленую окраску, иногда с сизоватым оттенком. Высота растений может достигать 1,5-2,0 м. Исключительно полиморфный вид, встречающийся в различных экологических условиях аридной зоны. Отличительная особенность солянки малолистной — высокая засухоустойчивость и способность произрастать на засоленных почвах.
В начальные фазы роста солянки малолистной корневая система энергично развивается и углубляется в почву (табл. 3). В фазу семядольных листочков при высоте растения 2,0±0,2 см корни углубляются в почву на 21,0±2,0 см, при высоте 2,5±0,5 см — на 30,4±1,8 см, при 8,0±1,2 см — на 43,0±2,8 см. В конце периода вегетации в 1-й год жизни (в фазу плодообразования) корни проникают на глубину 150,0±3,7 см.
В мае мы провели раскопку подземной части растения солянки малолистной 5-го года вегетации. Главный корень, толщина которого составляла 6 см, идет вертикально вниз. На глубине 12-15 см два мощных боковых корня, разойдясь в стороны, направляются вертикально вниз (рис. 3). Главный корень на глубине 37 см делится на два корня. Последние, в свою очередь, по мере углубления в почву еще несколько раз делились на более мелкие. Все корни солянки малолистной 1-го порядка, за исключением некоторых мелких, в слое 0-140 см имеют очень мало боковых ответвлений и только с глубины 140 см число боковых корней увеличивается. Во всех сильно уплотненных горизонтах, например на глубине 170-180 и 220-230 см, встречаются рыхлые прослойки толщиной 5-15 см. Корни, попав в эти прослойки, стелются в горизонтальном направлении, удаляясь от главного корня в стороны (на расстояние до 2 м и более). Они имеют много ответвлений в вертикальном направлении, которые, в свою очередь, разветвляются на множество мелких корешков. На глубине 300-320 см на корнях имеется очень много живых и мертвых корневых волосков. На глубине 500 см в почве часто встречаются пустые или содержащие рыхлые породы щели. Корни, попадая в них, образуют пучки тонких корешков, заполняющих эти пространства. Ниже 600 см располагается плотный, как бы сцементированный гравийный горизонт. Корни солянки малолистной, дойдя до этого горизонта, не проникают в него, а, сильно ветвясь, стелются над ним, образуя густую сеть из мелких корешков.
Саксаул черный [Haloxylon aphyllum (Minkw.) Iljin] — это безлистный кустарник (или полудерево) высотой 3-4 м. Ассимилирующая функция принадлежит ежегодно опадающим веточкам, суккулентный галофит (19). Способность саксаула черного успешно осуществлять полный жизненный цикл при высоких концентрациях солей в почвенно-грунтовой среде во многом реализуется за счет суккулентности благодаря превращению пластинчатых листьев в цилиндрические фотосинтезирующие органы, многослойности эпидермиса и утолщению кутикулы. Растение с широкой экологической устойчивостью к почвенному засолению: произрастает как на песчаных, так и на глинистых и щебенистых почвах с разной степенью засоленности. Выдерживает минерализацию грунтовых вод до 40 г/л. Корневая система мощная, глубоко проникающая в почву. Встречается преимущественно на тер-риториях с близким залеганием грунтовых вод, но может произрастать и в автоморфных условиях. Поедаемыми частями саксаула черного в осенне-зимний период служат годичные побеги, прошлогодние веточки, плоды. В побегах содержится 10-12 % протеина (в плодах — до 20 %), 2,2-2,7 % жира, 21,2-38,6 % зольных веществ, 39,3 % безазотистых экстрактивных веществ, 14,9 % клетчатки.
В 1-й год жизни корни саксаула черного в начале вегетации (25.IV.) проникают на глубину 29-36 см и в 4-5 раз превосходят по длине надземную часть. К концу вегетации они распространяются на глубину до 235 см, а в горизонтальном направлении — до 160 см. В предгорной пустыне (Нишанская степь) саксаул черный формирует в 5-летнем возрасте мощную и глубоко проникающую в почву корневую систему (рис. 4). Главный корень на глубине 30-40 см разветвляется на три корня диаметром 8-10 см, а на глубине 50 см один из корней делится, в свою очередь, еще на три части. Почва в слое 0-75 см довольно плотная, затем она становится рыхлее, а на глубине 300-360 см опять уплотняется. Дойдя до уплотненного горизонта, корни сильно разветвляются и по трещинам проходят вглубь почвы. На глубине 400-500 см наблюдается некоторое повышение влажности грунта, здесь строение почвы слоистое. В этих слоях корни мало ветвятся, уходя вглубь почвы. Корневая система в целом стремится вертикально вниз. На глубине 800 см залегает очень плотный слой толщиной 30 см. В этом слое корни утончаются, сплющиваются, пройдя плотный слой, принимают округлую форму. С глубины 860 см начинается дресвяный слой, переходящий в однородный песчаный горизонт. Здесь грунт влажен, при сжатии образуется комок. На глубине 1130-1200 см грунт супесчаный и очень влажный. Здесь корень сильно ветвится и образует большое число живых белых корешков. На глубине 1240 см грунт становится очень влажным, в комках при разламывании обнаруживаются капельки воды. На этой глубине в течение некоторого времени накапливалась вода (соленая).
Таким образом, у саксаула черного в 5-летнем возрасте формируется мощная и глубоко проникающая в почву корневая система универсального типа, приспособленная к использованию атмосферных осадков, конденсационной влаги и грунтовых вод.
Анализируя полученные результаты, следует отметить, что растения разных жизненных форм, у которых мы исследовали особенности формирования корневых систем, имеют разные типы адаптивной стратегии по классификации Раменского-Грайма. Саксаул черный, солянка малолистная, кохия простертая, будучи виолентами по Л.Г. Раменскому (28), или конкурентами по J. Grime (29), обладают высокой конкурентной способностью, характеризующейся быстрым ростом, умением захватывать и длительно удерживать территорию, подавляя соперника, и в полном объеме использовать ресурсы среды. Солянка восточная по типу адаптивной стратегии, согласно этой классификации, относится к патиентам. Растения с патиентными свойствами (28), или стресс-толеранты (29), — это виды, существующие не за счет высокой энергии жизнедеятельности, а благодаря выносливости при воздействии стрессовых факторов среды. Поэтому патиенты в зависимости от условий произрастания обладают устойчивостью к низкой обеспеченности водой, к засоленности почвы или другим неблагоприятным экологическим факторам среды. В формировании виолентных свойств саксаула черного, солянки малолистной, кохии простертой, патиентных свойств у солянки восточной в ксеротермических условиях среднеазиатской пустыни существенную роль играет способность экономно расходовать влагу на транспирацию. По нашим данным (4), полученным в среднеазиатской пустыне Карнабчуль, в апреле среднедневная интенсивность транспирации у саксаула черного, солянки малолистной, солянки восточной была в пределах 301,6-492,0 мг/ч. В этих же условиях полынь солелюбивая (Ar-temisiahalophileKrasch.) — типичный представитель пустынной флоры расходует на транспирацию в 2 раза больше воды — 957,7 мг/ч.
Таким образом, наши данные и результаты других исследователей подтверждают положение о том, что для кормовых кустарников (саксаул черный, солянка малолистная) и полукустарниковых галофитов (кохия простертая, солянка восточная) характерно экономное расходование воды на транспирацию. В процессе длительной эволюции растений от настоящих мезофитов до ксерогалофитов произошла глубочайшая адаптивная пере-стройка их морфологии, что прежде всего выразилось в редукции испаряющих органов (30). В результате у кохии простертой и солянки малолистной пластинчатые листья превратились в мелкие опушенные листочки, а у солянки восточной и саксаула черного — в цилиндрические ассимиляционные суккулентные мясистые листья с многослойной кутикулой (31), что обеспечивает сокращение поверхности испарения и снижает интенсивности транспирации (20).
В формировании различных типов адаптивных стратегий в процессе эволюции в ксеротермических условиях среднеазиатской пустыни, наряду с выработкой галоксерофилизированных свойств фотосинтезирующих органов, у кормовых кустарников и полукустарников важную роль играла способность к образованию быстро растущей и глубоко проникающей в почву корневой системы. Полученные нами результаты (см. рис. 1, рис. 2, рис. 3, рис. 4) показывают, что в условиях дефицита влаги, чрезмерно высокой температуры и сухости воздуха солянка восточная, кохия простертая (полукустарники), саксаул черный и солянка малолистная (кустарники) способны сохранять нормальную оводненность тканей (4), что достигается благодаря усиленному поглощению воды корнями (32). Отсюда следует, что способность растений к бесперебойному поглощению воды связана с активацией роста их корней (10). Оказалось, что недостаток почвенной влаги обусловливает усиление роста корней, увеличивая тем самым возможность поглощения воды (33). Таким образом, способность корней к непрерывному росту имеет определяющую роль в адаптации к дефициту воды, поскольку благодаря именно этому процессу растение получает ее в необходимом количестве (34).
Полученные данные показывают, что солянка восточная, кохия простертая, солянка малолистная, саксаул черный характеризуются высокой скоростью роста корневой системы, которая глубоко проникает в почву. В 1-й год жизни глубина корневой системы этих полукустарников и кустарников более чем в 4-5 раз превышает высоту их надземной части. Быстрорастущая и глубоко проникающая в почву корневая система полукустарниковых и кустарниковых галофитов обеспечивает их успешное функционирование в суровых условиях среднеазиатской пустыни и позволяет пережить длительный летний засушливый период. Мы обнаружили, что корни полукустарников и кустарников в период роста и развития проникают через очень плотные слои грунта, сравнимые по плотности с бетоном. В научной литературе обсуждается способность растений пробивать плотные слои почвы, которая связана с особенностями строения кончика корня, обеспечивающего преодоление сопротивления плотных сухих почв. Предполагается, что одним из механизмов этого может быть формирование корневых волосков, выступающих якорем при продвижении корня через сухие плотные слои почвы (35). Важную роль в поддержании непрерывного роста корней в плотных сухих слоях почвы может играть транспорт воды по флоэме к кончику корня. Его роль была установлена с помощью трехмерного моделирования распределения воды в зависимости от расположения флоэмных окончаний (36).
В.Г. Онипченко (37) описал явление так называемого гидравлического лифта, когда растение способно не только поднимать воду корнями, но также выделять ее в более сухие горизонты почвы. Явление гидравлического лифта широко распространено в аридных регионах (38). В условиях пустыни растения своими глубоко проникающими в почву корнями подни-мают воду в поверхностные слои, где этой водой могут пользоваться растения с неглубокой корневой системой. Подобные результаты получены и в других исследованиях в лесах сахарного клена (Acer saccharum Marshall), где растения нижних ярусов получали воду за счет обеспечиваемого кленом гидравлического лифта. Показано, что за ночь взрослое дерево сахарного клена может перекачивать из нижних горизонтов почвы в верхние около 100 л воды (39). Одно дерево зонтичной акации [Acaciatortilis (Forssk.) Hayne] в Африке поднимает за ночь от 70 до 235 л воды (40). В настоящее время описано более 90 видов растений с такой способностью (41, 42).
Гидравлический подъем — это пассивное перемещение воды от корней в слои почвы с более низким водным потенциалом, в то время как другие части корневой системы в более влажных слоях почвы, обычно на глубине, поглощают воду (43, 44). Гидравлическим перераспределением обеспечивается пассивное перемещение воды между различными частями почвы через корневые системы растений, обусловленное градиентами водного потенциала на границе раздела почва—растение. Гидравлическое перераспределение может иметь важные последствия в масштабах сообщества, влияя на чистую первичную производительность, а также на динамику водных запасов и развития растительности. В глобальном масштабе оно способно повлиять на гидрологические и биогеохимические циклы и, в конечном счете, на климат. Полученные нами результаты сравнения особенностей формирования корневой системы кустарников и полукустарников дают основание предполагать, что саксаул черный, солянка малолистная, солянка восточная и кохия простертая способны осуществлять функцию гидравлического лифта благодаря формированию глубоко проникающей в почву корневой системы. Выделение воды в почве за счет гидравлического лифта обеспечивает лучшее поглощение элементов минерального питания растениями из верхних сухих горизонтов почвы и усиление активности почвенных микроорганизмов (37).
Выявлено важное значение корневых систем галофитов для их устойчивости к засолению. Способность растений переносить засоленную среду определяется множеством физиологических и биохимических процессов, которые способствуют удержанию и/или поглощению воды, защищают функции хлоропластов и поддерживают гомеостаз ионов. У галофитов синтезируются осмотически активные метаболиты, специфические белки и определенные ферменты, улавливающие свободные радикалы. Многие галофиты накапливают метилированные метаболиты, которые играют решающую роль в качестве осмопротекторов и нейтрализуют свободные радикалы (45).
Разнообразие микроорганизмов, связанных с корнями растений галофитов, огромно. Это сложное микробное сообщество, которое называют вторым геномом растения, имеет решающее значение для его стрессоустойчивости. Растения способны формировать свой ризосферный микробиом, о чем свидетельствует тот факт, что разные виды растений являются хозяевами определенных микробных сообществ на одной и той же почве (46).
Недавние исследования показали, что использование ризобактерий галофитов благоприятно влияет на рост сельскохозяйственных растений и повышает их урожайность. Пять солеустойчивых бактерий были выделены из корней галофита Arthrocnemum indicum. В условиях солевого стресса инокулированные проростки арахиса сохраняли ионный гомеостаз, накапливали меньше активных форм кислорода и демонстрировали усиленный рост по сравнению с неинокулированными проростками (47). Инокуляция ризосферной бактерией AzospirillumbrasilenseNH, первоначально выделенной из засоленной почвы на севере Алжира, значительно усилила рост твердой пшеницы (Triticumdurumvar. waha) в условиях засоленной почвы. У инокулированных растений такие важные параметры, как скорость прорастания, высота стебля, длина колоса, сухая масса корней и побегов, содержание хлорофиллов a и b, масса 1000 семян, число семян в колосе и масса семян были значительно выше, чем у неинокулированных (48).
Галотолерантные бактерии способны адаптироваться к повышенной солености среды и поддерживать нормальное функционирование благодаря эффективным осморегуляторным механизмам. Ризобактерии галофитов стимулируют рост корней растений в условиях высокой засоленности, используя такие механизмы, как синтез индолуксусной кислоты, гиббереллинов, цитокининов, абсцизовой кислоты, солюбилизация нерастворимого фосфата, синтез 1-аминоциклопропан-1-карбоксилат-дезаминазы (АЦК-дезаминаза), снижающей в растениях содержание этилена, образуемого при солевом стрессе (49).
Итак, кормовые галофитные полукустарники кохия простертая, солянка восточная, кустарники саксаул черный, солянка малолистная при выращивании в ксеротермических условиях формируют мощные и проникающие глубоко в почву корневые системы: в первые годы жизни — на глубину 200-295 см, в 5-летнем возрасте на светлых сероземах предгорной среднеазиатской пустыни (Нишанская степь) — на глубину от 500-600 см у солянки восточной, кохии простертой и солянки малолистной до 1200 см (то есть в 2 раза глубже) у саксаула черного. Эти культуры формируют корневые системы универсального типа, приспособленные к использованию атмосферных осадков, конденсационной влаги и неглубоко залегающих грунтовых вод. Для растений характерен быстрый рост и развитие корневых систем: в 1-й год жизни они по длине больше надземной части в 4-4,5 раза, в возрасте 5 лет — в 6 раз. Благодаря такой способности корней обеспечивается поглощение необходимого растению количества воды в условиях дефицита влаги и при засухе. На глубину проникновения корневой системы растений разных жизненных форм (кустарники, полукустарники) сильное влияние оказывают водно-физические свойства эдафической среды. При постоянном недостатке почвенной влаги корневая система непрерывно стремится углубляться, пробивая плотные, сцементированные слои почвы. Мы полагаем, что полукустарниковым и кустарниковым галофитам присуща функция так называемого гидравлического лифта, когда растение способно не только поднимать воду корнями, но также выделять ее в более сухие горизонты почвы. В условиях среднеазиатской пустыни кормовые полукустарники (кохия простертая, солянка восточная) и кустарники (саксаул черный, солянка малолистная) быстрорастущими и глубоко проникающими корнями поднимают воду в поверхностные слои почвы, где этой водой могут пользоваться растения с неглубокой корневой системой.
1. Показатели роста и развития корневой системы солянки восточной (Salsola orientalis S.G. Gmel.) в 1-й год вегетации (n = 9, M±SEM; интродукционный питомник, зона предгорной пустыни, Нишанская степь, Кашкадарьинская область, Республика Узбекистан, 2016 год) |
|||
Дата и фаза развития |
Глубина проникновения корневой системы, см |
Ширина горизонтального распространения корня, см |
Отношение длины корней к высоте надземной части |
24.IV. Всходы |
44,0±5,3 |
16,5±3,3 |
4,7 |
31.V. Настоящие листья |
80,0±4,1 |
95,0±2,7 |
4,1 |
5.VII. Ветвление |
105,0±7,4 |
75,0±5,2 |
2,6 |
3.IX. Цветение |
125,0±5,7 |
85,0±7,3 |
2,5 |
2.XII. Плодообразование, конец вегетации |
200,0±11,2 |
145,0±6,4 |
3,4 |
2. Рост и развитие растений кохии простертой [Kochia prostrata (L.) Schrad.] разных экотипов в 1-й год вегетации (n = 75, M±SEM; интродукционный питомник, зона предгорной пустыни, Нишанская степь, Кашкадарьинская область, Республика Узбекистан, 2016 год) |
||||||
Экотип |
ВР |
ШК |
ЧП |
ГПК |
ГРК |
ЧК1 |
Каменистый |
92,0±1,4 |
100,0±6,3 |
32,0±4,2 |
240,0±12,6 |
115,0±13,6 |
29,0±2,1 |
Песчаный |
67,0±0,8 |
65,0±5,2 |
23,0±3,0 |
237,0±18,7 |
130,0±15,8 |
25,0±1,8 |
Песчаный |
73,0±2,7 |
36,0±2,5 |
18,0±3,6 |
270,0±10,5 |
100,0±19,6 |
19,0±2,3 |
Песчаный |
50,0±4,2 |
47,0±2,8 |
10,0±2,7 |
295,0±22,3 |
90,0±16,1 |
17,0±2,6 |
Солонцовый |
54,0±1,8 |
67,0±0,4 |
25,0±3,4 |
215,0±17,3 |
135,0±10,5 |
21,0±1,6 |
Примечание. ВР — высота растений, см; ШК — ширина кроны, см; ЧП — число побегов, шт.; ГПК — глубина проникновения корней, см; ГРК — горизонтальное распространение корней, см; ЧК1 — число корней 1-го порядка, шт. |
||||||
3. Рост и развитие растений солянки малолистной [Aelleniasubaphylla (С.А. Меу) Aellen] в 1-й год вегетации (n = 75, M±SEM; интродукционный питомник, зона предгорной пустыни, Нишанская степь, Кашкадарьинская область, Республика Узбекистан, 2016 год) |
|||
Дата и фаза развития |
Высота всходов и растений, см |
Глубина проникно- вения корней, см |
Число корней 1-го порядка |
24.IV. Всходы; семядольные листочки |
2,0±0,2 |
21,0±2,0 |
|
30.V. Появление настоящих листьев: |
|
|
|
двух |
2,5±0,5 |
30,4±1,8 |
5,6±1,4 |
четырех (5.VI.) |
6,4±0,4 |
36,0±4,3 |
7,4±1,1 |
шести (13.VI.) |
8,0±1,2 |
43,0±2,8 |
11,8±2,0 |
восьми (20.VI.) |
9,6±1,6 |
46,0±3,3 |
12,8±1,5 |
5.VII. Начало ветвления |
20,0±2,6 |
46,0±2,8 |
21,0±2,7 |
3.IX. Начало цветения |
55,0±1,4 |
53,0±1,5 |
50,0±5,8 |
2.XII. Плодообразование |
77,0±3,3 |
150,0±3,7 |
58,0±3,6 |
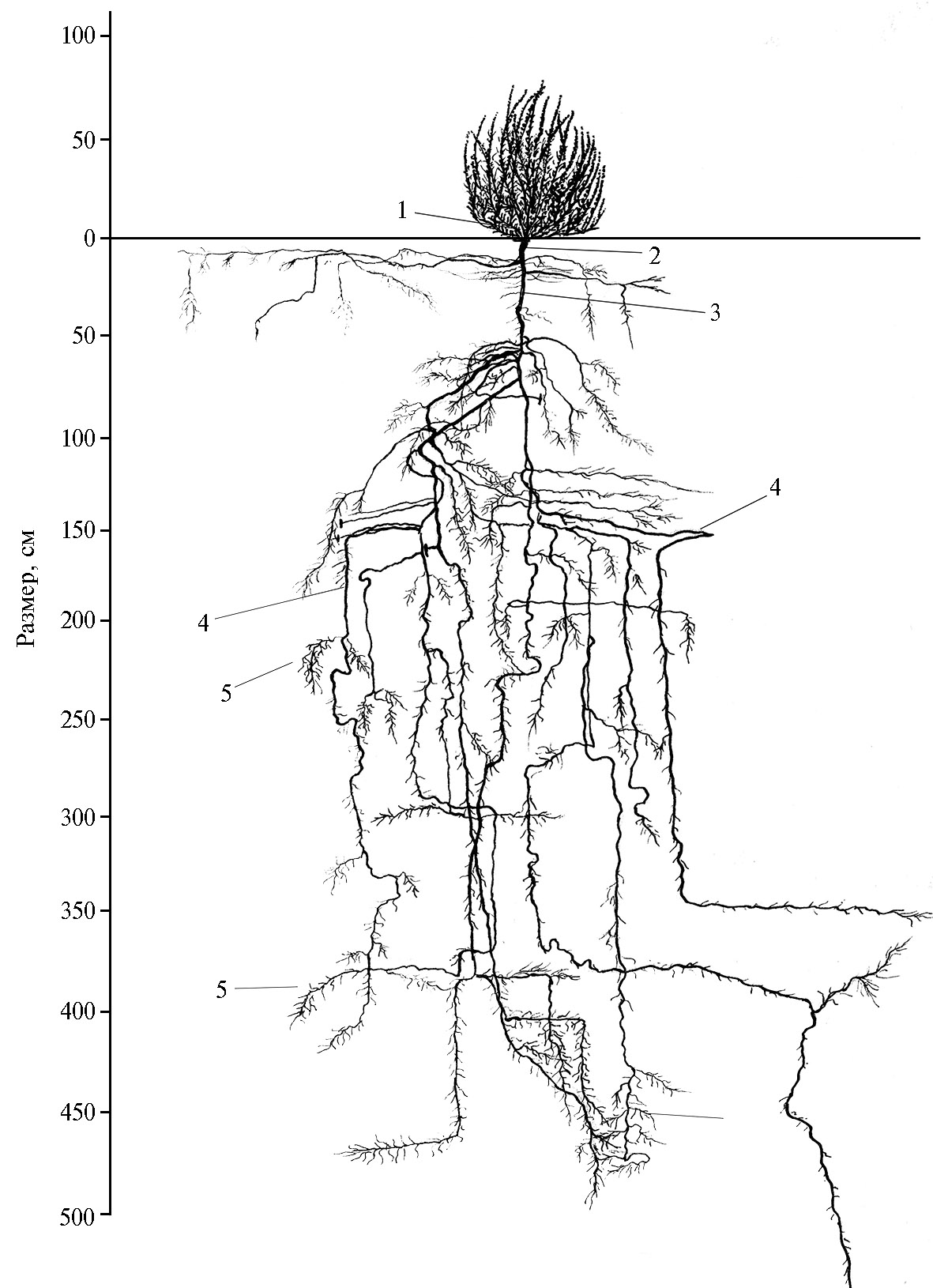
Рис. 1. Корневая система солянки восточной (Salsola orientalis S.G. Gmel.) в возрасте 5 лет на среднесуглинистых сероземах: 1 — почки возобновления, 2 — корневая шейка, 3 — главный корень, 4 — боковые корни, 5 — корневые волоски (интродукционный питомник, зона предгорной пустыни, Нишанская степь, Кашкадарьинская область, Республика Узбекистан, 2020 год).
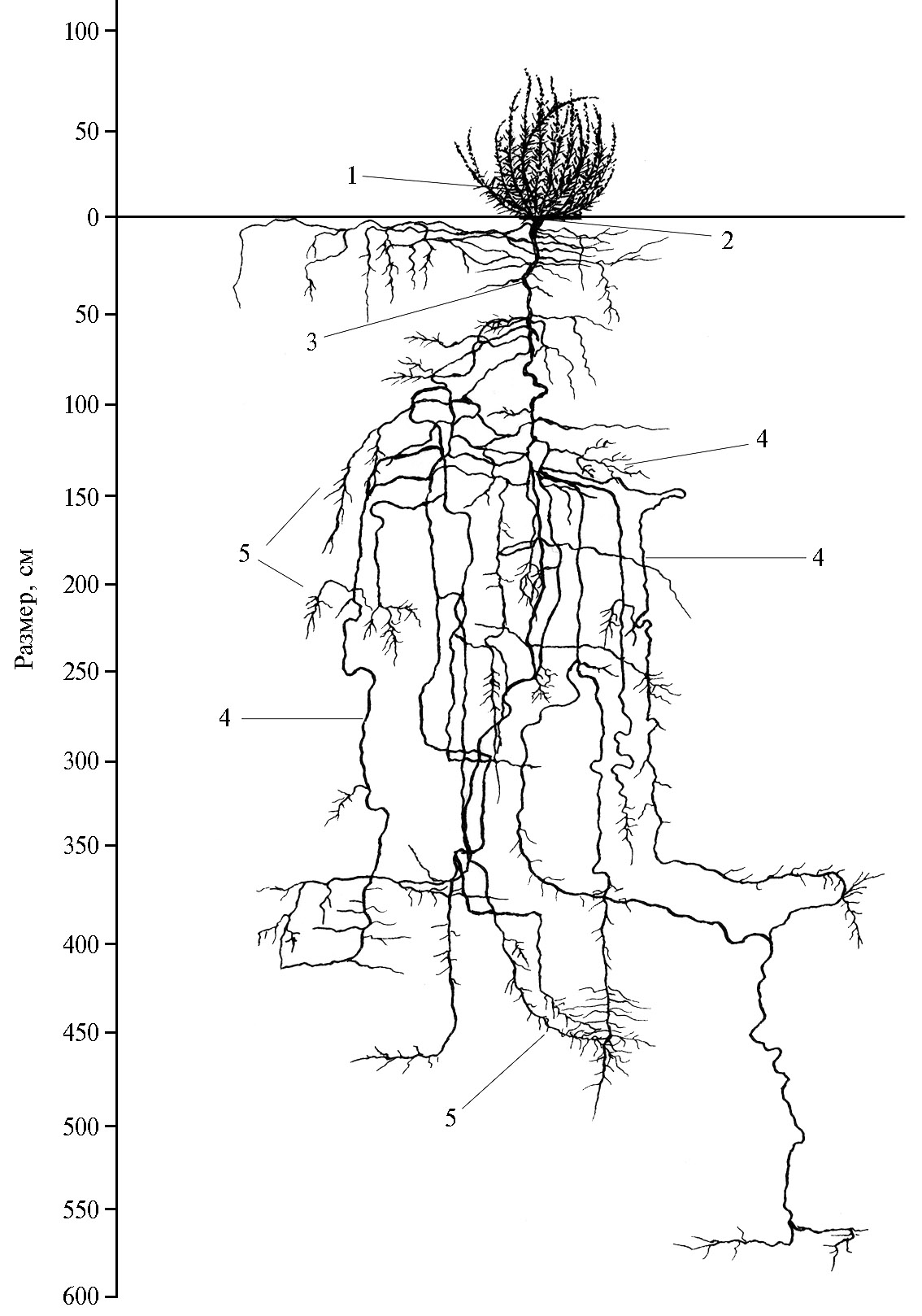
Рис. 2. Корневая система кохии простертой [Kochia prostrata (L.) Schrad.] каменистого экотипа в возрасте 5 лет на среднесуглинистых сероземах: 1 — почки возобновления, 2 — корневая шейка, 3 — главный корень, 4 — боковые корни, 5 — корневые волоски (интродукционный питомник, зона предгорной пустыни, Нишанская степь, Кашкадарьинская область, Республика Узбекистан, 2020 год).
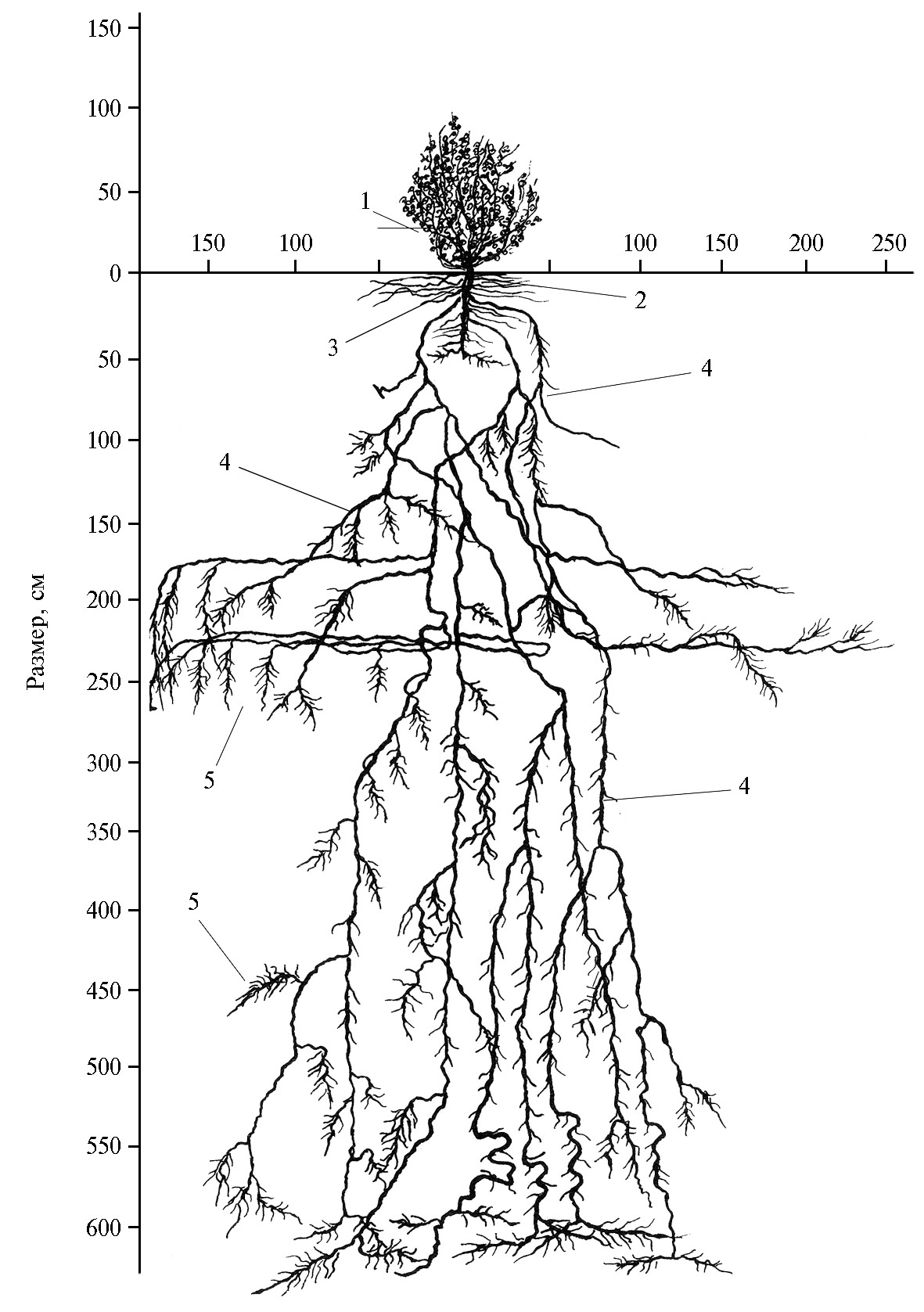
Рис. 3. Корневая система солянки малолистной [Aellenia subaphylla (С.А. Меу) Aellen] в возрасте 5 лет на среднесуглинистых сероземах: 1 — почки возобновления, 2 — корневая шейка, 3 — главный корень, 4 — боковые корни, 5 — корневые волоски (интродукционный питомник, зона предгорной пустыни, Нишанская степь, Кашкадарьинская область, Республика Узбекистан, 2020 год).
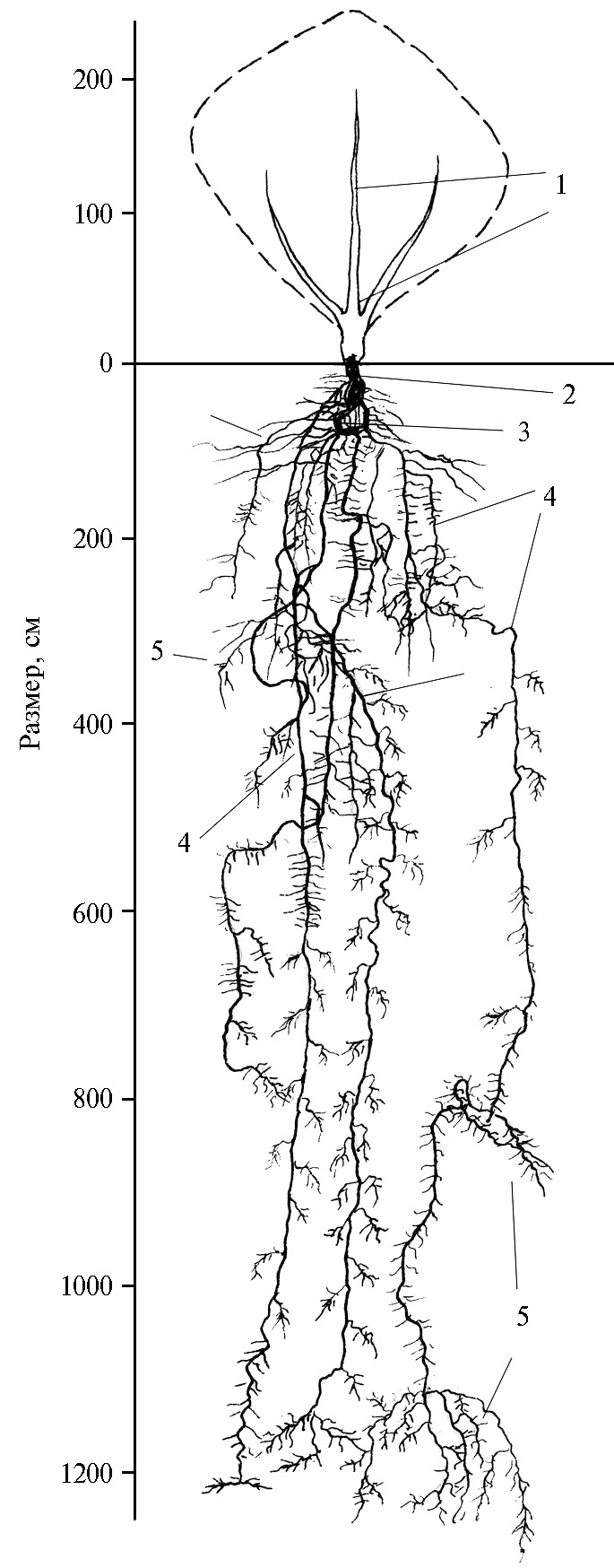
Рис. 4. Корневая система саксаула черного [Haloxylon aphyllum (Minkw.) Iljin] в возрасте 5 лет на среднесуглинистых сероземах: 1 — почки возобновления, 2 — корневая шейка, 3 — главный корень, 4 — боковые корни, 5 — корневые волоски (интродукционный питомник, зона предгорной пустыни, Нишанская степь, Кашкадарьинская область, Республика Узбекистан, 2020 год).
ЛИТЕРАТУРА
- Нечаева Н.Т., Николаев В.Н. Химический состав, питательность и биологическая полноценность пастбищных кормов подгорной равнины Туркменистана. Ашхабад, 1985.
- Barret-Lennard E.G., Malcolm C.V. Saltland Pastures in Australia: a practical guide. Bulletin 4312. Dept. ofAgriculture. WesternAustralia. SouthPerth, 1995.
- Нечаева Н.Т., Василевская В.К., Антонова К.Г. Жизненные формы растений пустыни Каракумы. М., 1973.
- Шамсутдинов З.Ш., Шамсутдинов Н.З. Галофитное растениеводство (эколого-биологические основы). М., 2005.
- Коровин Е.П. Растительность Средней Азии и Казахстана. Ташкент, 1961.
- Акжигитова Н.И. Галофильная растительность Средней Азии и ее индикационные свойства. Ташкент, 1982.
- Shamsutdinov N.Z., Shamsutdinova E.Z., Orlovsky N.S., Shamsutdinov Z.Sh. Halophytes: ecological features, global resources, and outlook for multipurpose use. Herald of the Russian Academy of Sciences, 2017, 87: 1-11 (doi: 10.1134/S1019331616060083).
- Bazzaz F.A., Ackerly D.D., Reekie E.G. Reproductive allocation in plants. In: Seeds: the ecology of regeneration in plant communities /M. Fenner (ed.). CAB International, Oxford, 2000.
- Waines J.G., Ehdaie B. Domestication and crop physiology: roots of green-revolution wheat. Annals of Botany, 2007, 100(5): 991-998 (doi: 10.1093/aob/mcm180).
- Gewin V. Food: an underground revolution. Nature, 2010, 466: 552-553 (doi: 10.1038/466552a).
- Geldner N., Salt D.E. Focus on roots. Plant Physiol., 2014, 166(2): 453-454 (doi: 10.1104/pp.114.900494).
- Shabala S. Learning from halophytes: physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot., 2013, 112(7): 1209-1221 (doi: 10.1093/aob/mct205).
- Liu L., Wang B. Protection of halophytes and their uses for cultivation of saline-alkali soil in China. Biology (Basel), 2021, 10(5): 353 (doi: 10.3390/biology10050353).
- Alishahi F., Alikhani H.A., Khoshkholgh-Sima N.A., Etesami H. Mining the roots of various species of the halophyte Suaeda for halotolerant nitrogen-fixing endophytic bacteria with the potential for promoting plant growth. Int. Microbiol., 2020, 23(3): 415-427 (doi: 10.1007/s10123-019-00115-y).
- Nikalje G.C., Nikam T.D., Suprasanna P. Looking at halophytic adaptation to high salinity through genomics landscape. Curr. Genomics, 2017, 18(6): 542-552 (doi: 10.2174/1389202918666170228143007).
- Flowers T.J., Muscolo A. Introduction to the Special Issue: halophytes in a changing world. AoB Plants, 2015, 7: plv020 (doi: 10.1093/aobpla/plv020).
- Судницын И.И. Движение почвенной влаги и водопотребление растений. М., 1979.
- Шалыт М.С. Методика изучения морфологии подземной части растений и растительных сообществ. В кн.: Полевая геоботаника /Под ред. Е.М. Лавренко, А.А. Корчагина. М.-Л., 1960, т. 2: 369-447.
- Ботаническая география Казахстана и Средней Азии (в пределах пустынной области) /Под ред. Е.И. Рачковской, Е.А. Волковой, В.Н. Храмцова. СПб, 2003.
- Ogburn R.M., Edwards E.J. The ecological water-use strategies of succulent plants. Advances in Botanical Research, 2010, 55: 179-225 (doi: 10.1016/S0065-2296(10)55004-3).
- Sage R.F., Christin P.A., Edwards E.J. The C4 plant lineages of planet earth. Journal of Experimental Botany, 2011, 62(9): 3155-3169 (doi: 10.1093/jxb/err048).
- Пьянков В.И., Мокроносов А.Т. Физиолого-биохимические основы дифференциации пустынных растений и проблемы фитомелиорации аридных экосистем. Проблемы освоения пустынь, 1991, 3-4: 161-170.
- Пьянков В.И. Роль фотосинтетической функции в адаптации растений к условиям среды. Автореф. докт. дис. М., 1993.
- Ротмистров В.Г. Районы распространения корней у однолетних культурных растений. Журнал опытной агрономии, 1907, V(VIII): 499-522.
- Waldron B.L., Greenhalgh L.K., ZoBell D.R., Olson K.C., Davenport B.W., Palmer M.D. Forage Kochia (Kochia prostrata) increases nutritional value, carrying capacity, and livestock performance on semiarid rangelands. Forage &Grazinglands, 2011, 9: 1-6 (doi: 10.1094/FG-2011-0301-01-RS).
- Erin C.G., Patricia S.M. Does Kochia prostrata spread from seeded sites? An evaluation from Southwestern Idaho, USA. Rangeland Ecol. Manage, 2013, 66: 191-203 (doi: 10.2111/REM-D-11-00177.1).
- Wang X., Wu J., Yang Z., Zhang F., Sun H., Qiu X., Yi F., Yang D., Shi F. Physiological responses and transcriptome analysis of the Kochia prostrata (L.) Schrad. to seedling drought stress. AIMS Genet., 2019, 6(2): 17-35 (doi: 10.3934/genet.2019.2.17).
- Раменский Л.Г. Избранные работы. Проблемы и методы изучения растительного покрова. Л., 1971.
- Grime J.P. Plants strategies and vegetation processes. John Wiley and Sons Ltd., Chichester, 1979.
- Григорьев Ю.С. О некоторых общих закономерностях ксерофилизации высших растений. Проблемы освоения пустынь, 1968, 5: 3-13.
- Breckle S.W. How do halophytes overcome salinity? In: Biology of salt tolerant plants /M.A. Khan, I.A. Ungar (eds.). Karachi, 1995.
- Кудоярова Г.Р., Холодова В.П., Веселов Д.С. Современное состояние проблемы водного баланса растений при дефиците воды. Физиология растений, 2013, 60(2): 155-165.
- Скобелева О.В., Ктиторова И.Н., Агальцова К.Г. Ускорение роста корней при дефиците нитрата связано с закислением апопласта. Физиология растений, 2010, 57: 520-529.
- Иванов В.Б. Клеточные механизмы роста растений. М., 2011.
- Bengough A.G., McKenzie B.M., Hallett P.D., Valentine T.A. Root elongation, water stress, and mechanical impedance: a review of limiting stresses and beneficial root tip traits. Journal of Experimental Botany, 2011, 62(1): 59-68 (doi: 10.1093/jxb/erq350).
- Wiegers B.S., Cheer A.Y., Silk W.K. Modeling the hydraulics of root growth in three dimensions with phloem water sources. Plant Physiology, 2009, 150: 2092-2103 (doi: 10.1104/pp.109.138198).
- Онипченко В.Г. Функциональная фитоценология: синэкология растений. М., 2013.
- Миркин Б.М., Наумова Л.Г. Введение в современную науку о растительности. М., 2017.
- Emerman S.H., Dawson T.E. Hydraulic lift and its influence on the water content of the rhizosphere: an example from sugar maple, Acer saccharum. Oecologia, 1996, 108(2): 273-278 (doi: 10.1007/BF00334651).
- Ludwig F., Dawson T.E., Kroon H., Berendse F., Prins H.H. Hydraulic lift in Acacia tortilistrees on an east African savanna. Oecologia, 2003, 134(3): 293-300 (doi: 10.1007/s00442-002-1119-x ).
- Ryel R.J. Hydraulic redistribution. In: Progress in botany. 65/K. Esser, U. Lüttge, W. Beyschlag, J. Murata (eds.). Springer, Berlin, Heidelberg, 2004.
- Liste H.H., White J.C. Plant hydraulic lift of soil water — implications for crop production and land restoration. Plant and Soil, 2008, 313(1-2): 1-17 (doi: 10.1007/s11104-008-9696-z).
- Caldwell M., Dawson T., Richards J. Hydraulic lift: consequences of water efflux from the roots of plants. Oecologia, 1998, 113: 151-161 (doi: 10.1007/s004420050363).
- Prieto I., Armas C., Pugnaire F.I. Water release through plant roots: new insights into its consequences at the plant and ecosystem level. New Phytologist, 2012, 193: 830-841 (doi: 10.1111/j.1469-8137.2011.04039.x).
- Asish K.P., Anath B.D. Salt tolerance and salinity effects on plants: a review. Ecotoxicology and Environmental Safety, 2005, 60: 324-349 (doi: 10.1016/j.ecoenv.2004.06.010).
- Berendsen R.L., Pieterse C.M., Bakker P.A. The rhizosphere microbiome and plant health. Trends Plant Sci., 2012, 17(8): 478-486 (doi: 10.1016/j.tplants.2012.04.001).
- Sharma S., Kulkarni J., Jha B. Halotolerant rhizobacteria promote growth and enhance salinity tolerance in peanut. Front.Microbiol., 2016, 7: 1600 (doi: 10.3389/fmicb.2016.01600).
- Nabti E., Sahnoune M., Ghoul M., Fischer D., Hofmann A., Rothballer M., Schmid M., Hartman A. Restoration of growth of durum wheat (Triticum durum var. waha) under saline conditions due to inoculation with the rhizosphere bacterium Azospirillum brasilense NH and extracts of the marine alga Ulva lactuca. J. Plant Growth Regul., 2010, 29: 6-22 (doi: 10.1007/s00344-009-9107-6).
- Nabti E., Schmid M., Hartmann A. Application of halotolerant bacteria to restore plant growth under salt stress. In: Halophiles. Sustainable development and biodiversity, vol. 6 /D. Maheshwari, M. Saraf (eds.). Springer, Cham, 2015: 235-259 (doi: 10.1007/978-3-319-14595-2_9).