doi: 10.15389/agrobiology.2020.1.66rus
УДК 633.15:581.132
Работа выполнена по гранту РФФИ 20-04-00465 А.
ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ И ФОТОХИМИЧЕСКАЯ АКТИВНОСТЬ ФОТОСИНТЕТИЧЕСКОГО АППАРАТА ЛИСТЬЕВ КУКУРУЗЫ (Zea mays L.) ПОД ВЛИЯНИЕМ ТИАМЕТОКСАМА
Д.А. ТОДОРЕНКО1, О.В. СЛАТИНСКАЯ1, J. HAO2, Н.Х. СЕЙФУЛЛИНА1,
Č.N. RADENOVIĆ3, Д.Н. МАТОРИН1, Г.В. МАКСИМОВ1, 4
В последнее десятилетие неоникотиноидные инсектициды активно используются для защиты растений от вредителей. При этом мало изучено их воздействие на сами растения, в частности на состояние фотосинтезирующих пигментов. В настоящей работе впервые показано, что обработка листьев кукурузы (Zea mays L.) инсектицидом тиаметоксамом (TMX) приводит к снижению функциональной активности фотосистемы II и уменьшению энергизации мембран тилакоидов. Кроме того, эффект воздействия тиаметоксама зависит от генотипа кукурузы. Цель работы заключалась в изучении действия пестицида тиаметоксама на фотосинтетические пигменты и фотохимическую активность фотосинтетического аппарата листьев кукурузы двух генотипов. Опыты проводили в 2018-2019 годах. Объектом исследования служили образцы листьев кукурузы инбредной линии zppl 225 и гибрида zp 341 с высокими показателями всхожести, качества зерна и урожайности (Institut za kukuruz «Zemun Polje»,г. Белград, Сербия). Семена проращивали до появления корешков (длина не менее 5 мм), после чего высаживали в грунт (смесь вермикулит: чернозем — 1:1) и выращивали в условиях 16-часового светового дня при постоянной температуре 25 °С. При появлении 3-го настоящего листа (длина более 4 см) растения опрыскивали раствором TMX в концентрации 0,2 мг/л. В качестве контроля использовали растения, выращенные в аналогичных условиях без опрыскивания TMX. Измерения проводили при достижении 5-м настоящим листом размера 12-14 см. Содержание фотосинтетических пигментов (хлорофилла a, bи каротиноидов) определяли спектрофотометрически в 100 % экстракте ацетона и рассчитывали по формулам Хольма-Веттштейна. Изменение конформации молекул каротиноидов регистрировали методом спектроскопии комбинационного рассеяния (КР). Светоиндуцированные кинетики быстрой флуоресценции (БФ), замедленной флуоресценции (ЗФ) и модулированного отражения при λ = 820 нм (МР) регистрировали одновременно с помощью мультифункционального анализатора растений M-PEA-2 («Hansatech Instruments», Великобритания). Индукционные кривые БФ (OJIP-кривые) анализировали с помощью стандартного JIP-теста. Установлено, что в фазу 5-го настоящего листа у инбредной линии zppl 225 содержание хлорофиллов в присутствии TMX снижалось с 0,74 до 0,61 мг/г: количество хлорофилла а уменьшилось на 17 %, хлорофилла b — на 24 %. Напротив, в листьях гибрида zp 341 при воздействии TMX изменений в пигментном составе не выявляли. OJIP-кривые у контрольных и обработанных TMX листьев имели типичный вид кривых с характерными фазами O-J, J-I и I-P, которые отражали процессы последовательного восстановления переносчиков в электрон-транспортной цепи фотосинтеза (ЭТЦ) между двумя фотосистемами. Воздействие пестицида на листья двух генотипов кукурузы проявлялось в снижении функционального состояния фотосистемы II, определенного по параметру флуоресценции (PIABS), который был получен на основании анализа OJIP-кривых с помощью JIP-теста. Сравнение PIABS в контроле и при воздействии TMX выявило статистически значимые (p < 0,05) различия: в листьях у образцов zppl 225 и zp 341, обработанных TMX, параметр PIABS снижался соответственно на 29 и 24 %. В листьях кукурузы были обнаружены изменения в быстрой фазе замедленной флуоресценции, связанные с уменьшением энергизации тилакоидной мембраны при воздействии TMX. Анализ максимальных скоростей окисления и восстановления Р700 (кинетика МР) свидетельствует об уменьшении пула акцепторов на акцепторной стороне ФСI в листьях zppl 225 при воздействии TMX. Обнаружено, что реакционные центры (РЦ) ФСI zp 341 проявили устойчивость к воздействию TMX (отсутствие изменений в редокс-превращениях Р700). TMX вызывал изменения конформации молекул каротиноидов, но не менял их содержание в листе. Предложенная комбинация методов быстрой флуоресценции, замедленной флуоресценции, модулированного отражения при λ = 820 нм и спектроскопии комбинационного рассеяния может стать основой для формирования эффективной технологии диагностики ранних дефектов состояния фотосинтетических пигментов при попадании пестицидов в интактное растение.
Ключевые слова: кукуруза, пестициды, тиаметоксам, хлорофилл, каротиноид, флуоресценция, комбинационное рассеяние, Рамановская спектроскопия.
В последнее десятилетие нейроактивные инсектициды на основе никотина (имидаклоприд, ацетамиприд, динотефуран, тиаметоксам) широко используются для защиты растений от вредителей. Неоникотиноиды действуют как агонисты ацетилхолина, связываясь с никотиновыми рецепторами ацетилхолина (nAChR) на постсинаптической мембране, что вызывает блокирование синаптической передачи, торможение проведения возбуждения и гибель насекомого (1). Однако нет единого мнения о влиянии неоникотиноидов на состояние самого растения (2). С одной стороны, неоникотиноиды способны улучшать морфологические и физиологическое характеристики некоторых растений даже в стрессовых условиях, что способствует их росту и повышению урожайности (3), с другой — существуют данные о фитотоксичности неоникотиноидов (4, 5).
Известно, что производство кукурузы осложняется воздействием различных абиотических и биотических факторов, а также способностью фитофагов повреждать растение на самых ранних стадиях развития. Перспективно исследовать изменения состояния пигментного состава растений кукурузы при действии неоникотиноидов как в лабораторных, так и полевых условиях.
Фотосинтетический аппарат (ФСА) высших растений, состоящий из пигмент-белковых комплексов, включает два типа пигментов — хлорофиллы и каротиноиды. После поглощения квантов света молекулы пигментов антенны переносят энергию на реакционные центры (РЦ) двух фотосистем, в возбужденном состоянии отдающих электроны акцепторам, которые далее восстанавливают НАДФ+ до НАДФН, синтезируют АТФ и впоследствии органические вещества. Состояние ФСА — один из чувствительных индикаторов стресса у растений (6). При его изучении эффективны флуоресцентные методы, обладающие высокой чувствительностью и позволяющие выявить нарушения состояния пигментов задолго до появления морфологических изменений (7, 8). При исследовании состояния молекул каротиноидов широко используется метод спектроскопии комбинационного рассеяния света (КР), который позволяет контролировать изменения конформации молекулы каротиноидов антенны фотосистемы II (ФСII), оценивая вклад валентных колебаний С=С-связей (I1520/I1006), переходы из плоской конфигурации молекулы в изогнутую (I960/I1006), а также изменения длины полиеновой цепи (I1520/I1160) (9).
В настоящей работе впервые показано, что обработка листьев кукурузы инсектицидом тиаметоксамом (TMX) приводит к снижению функциональной активности фотосистемы II и уменьшению энергизации мембран тилакоидов. Кроме того, эффект воздействия тиаметоксама зависит от генотипа кукурузы. Так, у инбредной линии zppl 225 показано уменьшение содержания хлорофилла и пула акцепторов на акцепторной стороне ФСI. Тиаметоксам вызывает противоположные изменения конформации молекул каротиноидов в антенне, но не меняет их содержание.
Цель работы заключалась в изучении действия пестицида тиаметоксама на фотосинтетические пигменты и фотохимическую активность фотосинтетического аппарата листьев кукурузы двух генотипов.
Методика. Опыты проводили в 2018-2019 годах. Исследовали образцы листьев кукурузы (Zea mays L.) инбредной линии zppl 225 и гибрида zp 341 с высокими показателями всхожести, качества зерна и урожайности (Institut za kukuruz «Zemun Polje», г. Белград, Сербия) (10). Семена проращивали до появления корешков (длина не менее 5 мм), после чего высаживали в грунт (смесь вермикулита с черноземом 1:1) и выращивали в условиях 16-часового светового дня при постоянной температуре 25 °С. При появлении 3-го настоящего листа длиной более 4 см (10-е сут после проращивания) растения опрыскивали раствором тиаметоксама [5-метил-3-(2-хлортиазол-5-илметил)-1,3,5-оксадиазинан-4-илиден-N-нитроамин] в концентрации 0,2 мг/л. В качестве контроля использовали листья кукурузы, выращенные в аналогичных условиях без опрыскивания тиамет-оксамом. Измерения проводили при достижении 5-м настоящим листом размера 12-14 см.
Содержание фотосинтетических пигментов (хлорофиллов a, b и каротиноидов) определяли спектрофотометрически в 100 % экстракте ацетона на приборе Hitachi-557 («Hitachi», Япония) и рассчитывали по формулам Хольма-Веттштейна (11). Изменение конформации молекул каротиноидов регистрировали методом спектроскопии КР при помощи КР-спектрометра ДФС 24 («ЛОМО», Россия) с лазером 473 нм (Ciel, «Euro-lase», Германия) и системой регистрации МОРС 1/3648 («ЛОМО», Россия) на базе линейной ПЗС-матрицы TCD1304DG («Toshiba», Япония) с фильтром LPO2-473RS-50 («Shemrock», США). В эксперименте фрагмент листовой пластинки фиксировали на предметном столике и регистрировали сигнал КР в течение 5 с, мощность лазера на образце 3 мВт.
Светоиндуцированные кинетики быстрой флуоресценции (БФ), замедленной флуоресценции (ЗФ) и модулированного отражения при λ = 820 нм (МР) регистрировали одновременно с помощью мультифункционального анализатора растений M-PEA-2 («Hansatech Instruments», Великобритания). Интенсивность действующего света и продолжительность освещения составляли соответственно 3000 мкмоль квантов·м-2·с-1 и 60 с. Измерения проводили на адаксиальной стороне интактных листьев, помещенных в измерительный блок в специальной клипсе-прищепке. Перед измерением растения адаптировали к темноте в течение 15 мин, чтобы реакционные центры (РЦ) фотосистем перешли в «открытое» состояние с окисленным QA. Три сигнала регистрировали при чередовании световых и темновых интервалов, длительность которых подробно описана R.J. Strasser с соавт. (12).
Индукционные кривые БФ (OJIP-кривые) анализировали с помощью стандартного JIP-теста, описывающего потоки энергии через различные участки электрон-транспортной цепи фотосинтеза (13) при 20 мкс (FO), 270 мкс (F270мкс), 2 мс (FJ), 30 мс (FI) и максимальной степени флуоресценции (FM). Эти характеристики использовали для расчета максимального квантового выхода фотохимии ФСII как отношения переменной флуоресценции (FV) к максимальной (FM) FV/FM = 1 - FO/FM, вероятности транспорта электронов за пределы QA- ψEo = 1 - (FJ -FO)/(FM - FO), потока энергии (ABS), поглощаемой одним активным реакционным центром (RC), как ABS/RC = 4 × (F270мкс - FO) × (FM - FO)-3 × (FJ - FO)-1 × FM и показателя функциональной активности ФCII, отнесенной к поглощаемой энергии (индекс производительности ФCII): PIABS = (ABS/RC)-1 × FV/FM × (1 - FV/FM)-1 × ψEo × (1 - ψEo)-1 (13).
Кинетики модулированного отражения/поглощения света при λ = 820 нм (МР) характеризуют окислительно-восстановительные превращения пигмента ФСI (Р700) и пластоцианина (12). В нашем исследовании сигнал МР был нормирован по МР0 и представлен как МР/МР0 - 1, где МР0 — интенсивность сигнала рассеяния при 0,7 мс.
Статистическую обработку полученных данных проводили, используя непараметрический критерий для множества независимых переменных Краскела-Уоллиса (Kruskal-Wallis test) с помощью пакета Statistica v.10 («StatSoft, Inc.», США). Значения p < 0,05 считались статистически значимыми. В таблице представлены значения среднего (M) и стандартные ошибки среднего (±SEM).
Результаты. Пигментный состав листьев мало различался у двух генотипов кукурузы (zppl 225 и zp 341), но после действия тиаметоксама были обнаружены явные изменения, зависящие от генотипа. Так, в листьях у инбредной линии zppl 225 в присутствии TMX выявили снижение содержания хлорофиллов с 0,74 до 0,61 мг/г: количество хлорофилла а уменьшилось на 17 %, хлорофилла b — на 24 % (p < 0,05). Напротив, при воздействии TMX на листья гибрида zp 341 изменений в пигментном составе не обнаружили.
Индукционные кривые БФ (OJIP) как у контрольных, так и у обработанных TMX листьев имели типичный вид кривых с характерными фазами O-J, J-I и I-P, которые отражали процессы последовательного восстановления переносчиков в электрон-транспортной цепи фотосинтеза (ЭТЦ) между двумя фотосистемами (13) (рис. 1, А, Г). Исходный уровень O (origin) соответствует интенсивности флуоресценции при «открытых» РЦ ФСII (FO), когда все акцепторы в фотосистеме II (ФСII) находятся в окисленном состоянии и способны принять электрон. Нарастание флуоресценции в фазе O-J обусловлено светоиндуцированным восстановлением состояний QA-. Последующие фазы J-I-P отражают дальнейший рост флуоресценции, связанный с накоплением состояний QA- в результате восстановления акцепторов QB, пула хинонов и первичных акцепторов фотосистемы I (ФСI) (14).
При воздействии TMX у инбредной линии zppl 225 и гибрида zp 341 наблюдались небольшие изменения в форме OJIP (см. рис. 1, А, Г). Для более детального анализа индукционные кривые были дважды нормированы (на уровни O и P) и представлены в виде относительной переменной флуоресценции (Vt) (см. рис. 1, Б, Д) и как разность значений функций Vt между контролем и образцами, обработанными TMX (ΔVt) (см. рис. 1, В, Е). Воздействие пестицида на листья двух генотипов кукурузы проявлялось в увеличении ΔVt в фазе O-J, что свидетельствует о накоплении восстановленного QA в результате увеличения доли QB-невосстанав-ливающих центров ФСII, то есть центров, не способных к передаче электрона по электрон-транспортной цепи (ЭТЦ) (15). В листьях инбредной линии zppl 225 изменения в фазе O-J оказались более выражены, чем у гибрида zp 341. Кроме того, в листьях zppl 225 эффект воздействия TMX сопровождался увеличением амплитуды фазы J-I: ΔVt увеличивалось в фазах O-J и J-I, что свидетельствует о накоплении восстановленных форм QA и молекул пластохинонов, которые не способны к передаче электронов для темновых реакций фотосинтеза (8).
Индукционные кривые БФ анализировали с помощью JIP-теста (13) (табл.). Известно, что наиболее адекватный параметр JIP-теста — величина отношения FV/FM, которое коррелирует с максимальным квантовым выходом первичной фотохимической реакции ФСII и используется как показатель эффективности фотосинтеза (6). Величина FV/FM в контроле в фазу 5-го листа у zppl 225 и zp 341 составила 0,74±0,01. Полученные значения фотосинтетической активности, определенные по FV/FM, были сопоставимы со значениями FV/FM, полученными для растений C4-фотосинтеза (16). При воздействии TMX в листьях zppl 225 и zp 341 статистически значимых различий по параметру FV/FM не выявляли (р > 0,05).
В отличие от параметра FV/FM, индекс производительности PIABS существенно менялся при воздействии TMX. Известно, что PIABS коррелирует с жизнеспособностью растения и отражает текущее состояние работы ФСА в условиях стресса (17). Сравнение PIABS в контроле и при воздействии TMX выявило статистически значимые (p < 0,05) различия: в листьях у образцов zppl 225 и zp 341, обработанных TMX, параметр PIABS снижался соответственно на 29 и 24 %.
PIABS — интегральный показатель, который включает в себя три независимых параметра: долю активных РЦ (ABS/RC), эффективность переноса электронов захваченным экситоном в цепи переноса электронов за пределы QA (ψEo) и вероятность того, что экситон будет связан в РЦ (FV/FM) (13). Снижение PIABS в листьях zppl 225 в присутствии TMX было обусловлено значительным уменьшением эффективности электронного транспорта на акцепторной стороне ФСII (ψEo), что подтверждается изменениями в индукционных кривых БФ, представленных выше (p < 0,05). Напротив, снижение PIABS в листьях zp 341 могло быть связано с незначительным уменьшением доли активных РЦ (увеличение ABS/RC) и фотохимии ФСII (FV/FM).
Окислительно-восстановительные превращения реакционных центров молекул ФСI (P700) в присутствии TMX мы оценивали по поглощению света при λ = 820 нм (МР) (18) (рис. 2, А, В). Известно, что кинетика светоиндуцированного спада сигнала МР в первые 15-20 мс отражает процесс окисления Р700 (быстрая фаза) и достигает минимума при ~ 20 мс (МРмин)· МРмин — переходное устойчивое состояние с равными скоростями окисления и ревосстановления Р700. Впоследствии скорость ре-восстановления преобладает над скоростью окисления за счет донирования электронов от ФСII, приводя к уменьшению величины поглощения при λ = 820 нм и росту сигнала МР (медленная фаза) до максимума при ~ 200 мс (МРмакс) (12).
В присутствии TMX в листьях кукурузы инбредной линии zppl 225 и гибрида zp 341 были выявлены изменения в окислительно-восстановительных превращениях Р700 (см. рис. 2, А, В). Отметим, что у хлорофилла в листьях zppl 225 были обнаружены изменения в быстрой и медленной фазах МР, связанные с уменьшением амплитуды при ~ 20 мс (см. рис. 2, А). Анализ максимальных скоростей окисления и восстановления Р700 (кинетика МР) свидетельствует об уменьшении образования окисленных и увеличении — восстановленных форм акцепторов ФСI (P700) при воздействии TMX на листья zppl 225. Вероятно, это связано с уменьшением пула акцепторов, таких как P700, на акцепторной стороне ФСI. Напротив, РЦ ФСI листьев гибрида zp 341 проявляли резистентность к воздействию TMX (отсутствие изменений в МР кинетиках) (см. рис. 2, В).
Степень энергизации тилакоидных мембран мы оценивали по пикам I1 и I2 замедленной флуоресценции, измеренной в микросекундном временнóм диапазоне (0-0,09 мс). Известно, что ЗФ пропорциональна скорости реакций рекомбинаций в ФСII, на которые влияет энергизация тилакоидной мембраны (6). В процессе работы мы анализировали быструю фазу ЗФ, связанную с формированием потенциала на мембране. Известно, что быстрая фаза ЗФ имеет два пика I1 (~ 7 мс) и I2 (~ 100 мс), после чего падает до минимума D2 (~ 200 мс) (19). На рисунке 2 (Б, Г) представлены индукционные кривые ЗФ листьев zppl 225 и zp 341, обработанных TMX, нормированные на уровень минимальной флуоресценции D2. При действии ТМХ в листьях zppl 225 и zp 341 снижалась амплитуда пика I1 по сравнению с контролем соответственно на 23 и 21 %. Уменьшение пика I1 в микросекундном временнóм диапазоне могло быть вызвано снижением скорости реокисления QA (нарушение на акцепторной стороне ФСII) и/или уменьшением восстановления Z+ от 4MnCa кластера кислород-выделяющего комплекса (КВК) (19).
Состояние КВК и его способность донировать электрон для Р680+ через Z+ мы контролировали по появлению дополнительного пика на индукционной кривой БФ. Для этого рассчитывали относительную флуоресценцию между пиками O и J как WOJ = (Ft - FO)/(FJ - FO) (см. рис. 3, А, В) и разность значений функций между контролем и образцами, обработанными TMX как ΔWOJ = WO(TMX) - WOJ (контроль) (см. рис. 3, Б, Г). В листьях zppl 225 и zp 341 при действии TMX был обнаружен дополнительный пик между O и J при ~ 300 мкс (пик К) (см. рис. 3, Б, Г), который характерен для высокотемпературного стресса за счет ограничения переноса электронов к РЦ из-за нарушений в работе КВК (13).
В спектрах комбинационного рассеивания (КР) листьев кукурузы были выявлены характерные для каротиноидов полосы (960, 1006, 1156, 1190, 1200 и 1520 см-1), обусловленные валентными колебаниями электронов в полиеновой цепочке молекулы (рис. 4). Максимальные изменения амплитуды полос спектра КР наблюдались для соотношений I1520/I1006 (вклад валентных колебаний –С=С– связей полиеновой цепи каротиноида), I1520/I1006 (изменение вклада –С=С– связей по отношению к –С–С– связям полиеновой цепи) и I1006/I960 (расположение полиеновой цепи каротиноида относительно пиррольных колец) (20). Наличие в спектрах КР пиков на 1190 и 1200 см-1 указывало на состояние каротиноидов, характерное для 15-cis конформации (данные не представлены).
После опрыскивания листьев кукурузы TMX на-блюдалось увеличение соотношения полос I960/I1006 (см. рис. 4), что может указывать на изменение конформации молекул каротиноидов в антенне ФСА. При действии TMX в листьях инбредной линии zppl 225 менялась кон-формация полиеновой цепи молекулы каротиноидов, что проявлялось в снижении вклада валентных колебаний –С=С– на 26 % (соотношение I1520/I1006). В листьях гибрида zp 341 в присутствии TMX было отмечено увеличение величины соотношений I1520/I1160 и I1520/I1006 соответственно на 16 и 43 %, что свидетельствует об изменении в листьях гибрида zp 341 конформации каротиноидов, но не их содержания.
В настоящей работе мы исследовали влияние пестицида TMX на фотосинтетические пигменты и фотохимическую активность фотосинтетического аппарата листьев кукурузы двух генотипов с помощью современных спектральных и флуоресцентных методов.
Инбредная линия кукурузы zppl 225 была подвержена большим изменениям при опрыскивании листьев TMX, чем гибрид zp 341. У инбредной линии zppl 225 выявлено уменьшение содержания хлорофиллов, которое, вероятно, свидетельствует о том, что при действии TMX снижается синтез этих пигментов. Отметим, что в листьях zppl 225 выявлено уменьшение соотношения хлорофиллы (a + b)/каротиноиды, что, как правило, коррелирует с увеличением содержания каротиноидов в условиях стресса (21). Напротив, у листьев гибрида zp 341 изменений в пигментном составе не выявлено. Ранее на примере хлопка показано, что неоникотиноиды не влияют на содержание хлорофиллов, тогда как у листьев бамии при воздействии ацетамиприда зарегистрировано постепенное увеличение общего содержания хлорофилла (22). В то же время TMX может выступать в качестве биоактиватора, увеличивая физиологическую и метаболическую активность такой культуры, как рис (23).
Влияние TMX на состояние ФСII, ФСI, электронных переносчиков между фотосистемами в листьях кукурузы zppl 225 и zp 341 оценивали, исходя из набора параметров, полученных после анализа кинетических кривых БФ, ЗФ и МР. Анализ кинетических кривых БФ с помощью JIP-теста, который описывает потоки энергии через различные участки электрон-транспортной цепи фотосинтеза (13), позволил выявить, что TMX оказывал незначительное (р > 0,05) воздействие на фотохимию ФСII (FV/FM) в листьях двух изученных генотипов. Обнаружены изменения на акцепторной стороне ФСII, связанные со снижением вероятности электронного транспорта после акцептора QA- (ψEo), в листьях инбредной линии zppl 225. Снижение эффективности электронного транспорта на акцепторной стороне ФСII (ψEo) вызывало уменьшение функциональной активности ФСII (PIABS). Другие неоникотинодные пестициды, такие как ацетамиприд, также не оказывают воздействия на FV/FM и приводят к значительному снижению функциональной активности ФСII за счет уменьшения размера пула и снижения электронного транспорта на акцепторной стороне ФСII у цианобактерий Synechocystis sp. (24).
Таким образом, при опрыскивании растений кукурузы пестицидом тиаметоксамом (TMX) было выявлено снижение содержания хлорофилла в листьях инбредной линии zppl 225 по сравнению с гибридом zp 341. Кроме того, в листьях zppl 225 обнаружено уменьшение пула акцепторов на акцепторной стороне фотосистемы I (ФСI). Установлено, что TMX воздействует на функциональную активность фотосистемы II (ФСII), уменьшая параметр PIABS, и снижает формирование потенциала на тилакоидной мембране (I1) как у zppl 225, так и у zp 341. TMX вызывал противоположные изменения конформации молекул каротиноидов в антенне, но не менял их содержание, что, по-видимому, связано с более эффективным рассеянием избыточной энергии через систему каротиноидов в листьях zp 341 по сравнению с zppl 225. Предложенная комбинация методов позволяет эффективно оценивать функциональное состояние фотосинтетического аппарата в листьях в полевых и лабораторных условиях как при биомониторинге, так и при сравнении различных линий сельскохозяйственных культур.
Параметры JIP-теста, полученные после анализа индукционных кривых OJIP, для листьев растений кукурузы (Zea mays L.) двух генотипов при действии тиаметоксама (TMX) (M±SEM) |
||||
Параметр |
zppl 225 |
zppl 225 + TMX |
zp 341 |
zp 341 + TMX |
FV/FM |
0,740±0,004 (100 %) |
0,740±0,004 (99 %) |
0,740±0,010 (100 %) |
0,710±0,010 (97 %) |
ψEo |
0,69±0,01 (100 %) |
0,65±0,01 (93 %)* |
0,67±0,01 (100 %) |
0,66±0,02 (100 %) |
ABS/RC |
2,62±0,09 (100 %) |
2,93±0,13 (112 %) |
2,57±0,12 (100 %) |
2,95±0,18 (115 %) |
PIABS |
2,50±0,10 (100 %) |
1,77±0,15 (71 %)* |
2,24±0,23(100 %) |
1,70±0,09 (76 %)* |
Примечание. FV/FM — максимальный квантовый выход фотохимии ФСII, ψEo — вероятность транспорта электронов за пределы QA- , ABS/RC — поток энергии, поглощаемый одним активным рекционным центром (РЦ), PIABS — показатель функциональной активности ФCII, отнесенной к поглощаемой энергии. В скобках представлены значения в процентах от контроля. |
||||
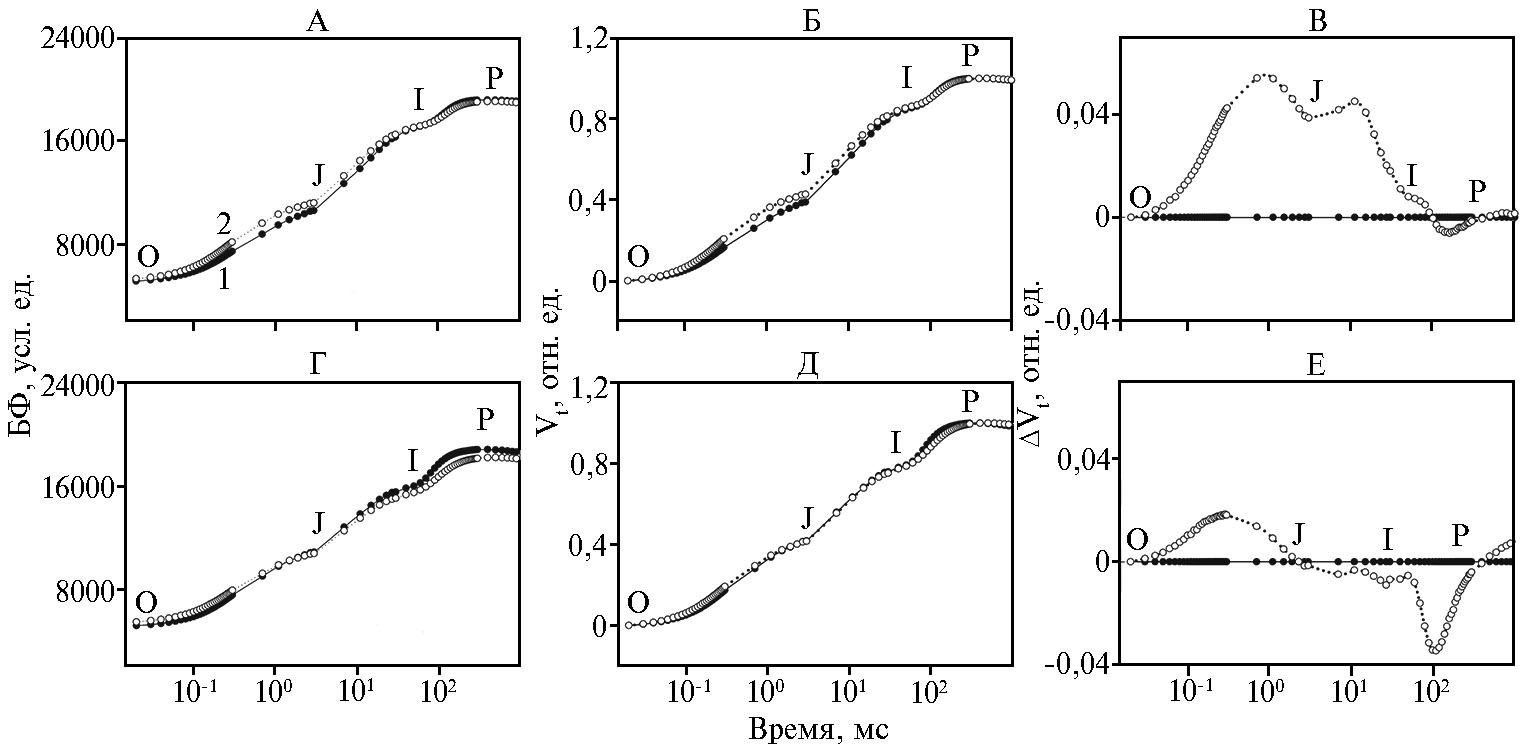
Рис. 1. Индукционные кривые (OJIP) быстрой флуоресценции (БФ) листьев кукурузы (Zea mays L.) (А, Г), кривые в виде относительной переменной флуоресценции Vt = (Ft- FO)/(FM - FO) после нормирования на уровни O и P (Б, Д) и разности значений функций ΔVt = Vt(TMX) - Vt(контроль) (В, Е) для инбредной линии zppl 225 (А, Б, В) и гибрида zp 341 (Г, Д, Е) в контроле (1) и при опрыскивании тиаметоксамом (2). O, J, I и P — пики на кривой индукции, наблюдаемые соответственно при 20 мкс, 2 мс, 30 мс и 300 мс.
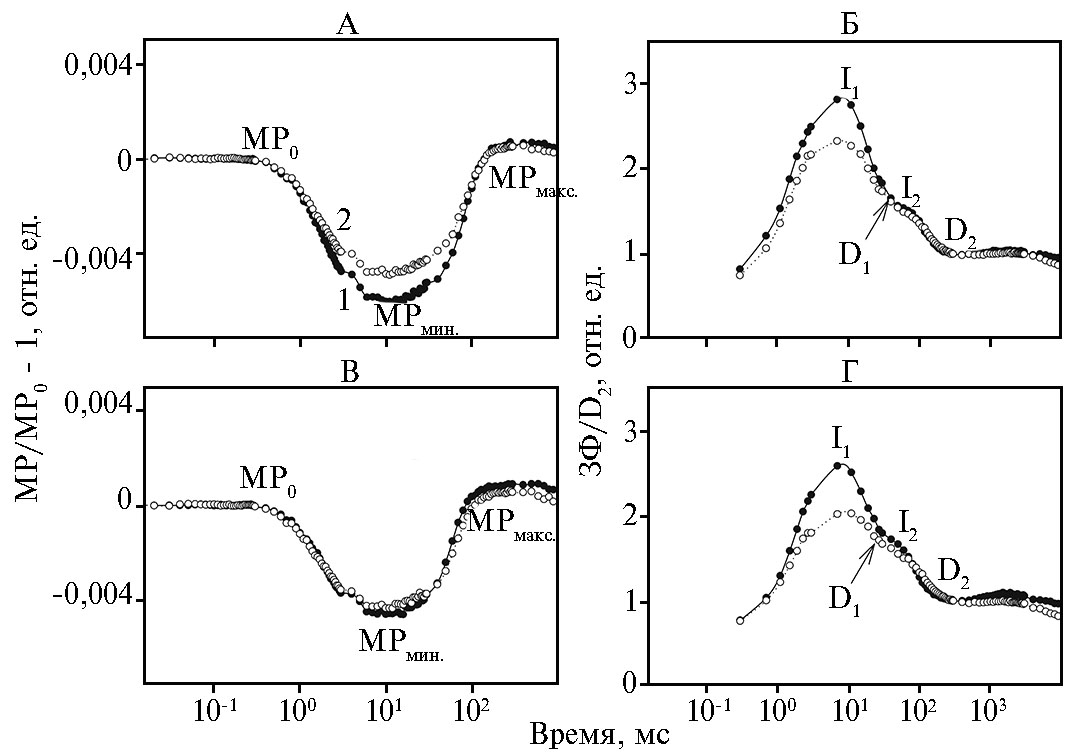
Рис. 2. Светоиндуцированные кинетики модулированного отражения при λ = 820 нм (MР) (А, В) и замедленной флуоресценции (ЗФ), нормированные на минимальный уровень сигнала D2 при 200 мс (Б, Г), у листьев кукурузы (Zea mays L.) инбредной линии zppl 225 (А, Б) и гибрида zp 341 (В, Г) в контроле (1) и при опрыскивании тиаметоксамом (2). Кинетики МР нормированы и представлены в виде МР/МР0 – 1, где МР0 — интенсивность сигнала при 0,7 мс. I1, I2, D1, D2 — максимумы и минимумы на индукционной кривой ЗФ.
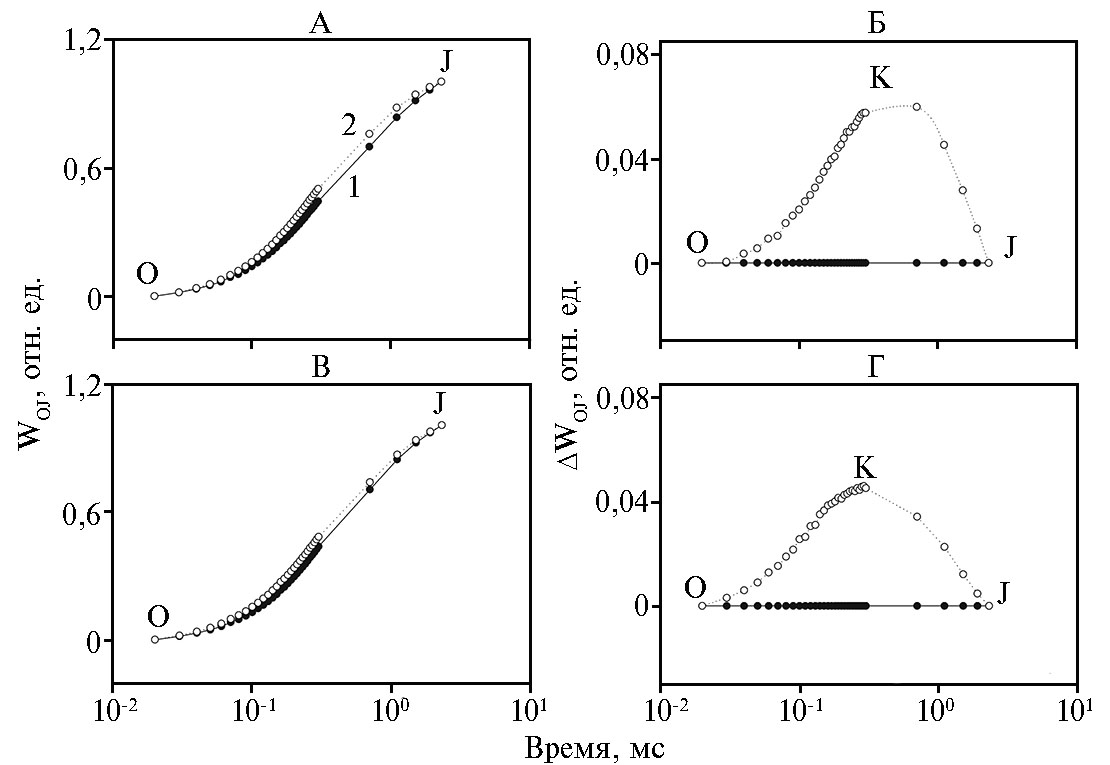
Рис. 3. Относительная переменная флуоресценция WOJ = (Ft - FO)/(FJ - FO) между уровнями O и J (А, В) и разность значений функций ΔWOJ = WOJ(TMX) - WOJ(контроль) (Б, Г) у листьев кукурузы (Zea mays L.) инбредной линии zppl 225 (А, Б) и гибрида zp 341 (В, Г) в контроле (1) и при опрыскивании тиаметоксамом (2). O, K, J — пики на кривой индукции, наблюдаемые соответственно при 20 мкс, 300 мкс и 2 мс.
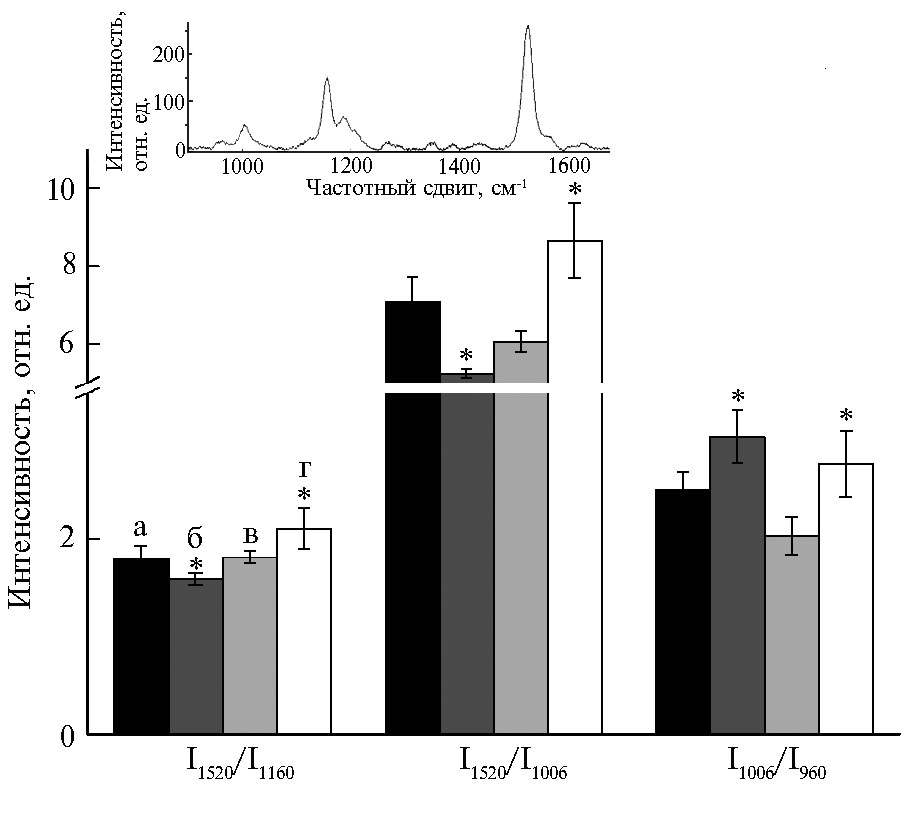
Рис. 4. Изменение спектров комбинационного рассеивания (КР) каротиноидов в листьях кукурузы (Zea mays L.) двух генотипов при опрыскивании тиаметоксамом (TMX): а — инбредная линия zppl 225, б — zppl 225 + ТМХ, в — гибрид zp 341, г — гибрид zp 341 + ТМХ. Звездочкой обозначены статистически значимые различия (p < 0,05). На вставке представлен спектр КР каротиноида листа кукурузы инбредной линии zppl 225.
ЛИТЕРАТУРА
- Ensley S.M. Neonicotinoids. Veterinary toxicology (third edition). Basic and clinical principles /R.C. Gupta (ed.). Academic Press, 2018: 521-524 (doi: 10.1016/C2016-0-01687-X).
- Georgieva M., Tsenov B., Dimitrova A. Dual effects of N-nitroguanidine neonicotinoids on plants. Genetics and Plant Physiology, 2017, 7(3-4): 107-120.
- da Silva Almeida A., Villela F.A., Nunes J.C., Meneghello G.E., Jauer A. Thiamethoxam: an inseticide that improve seed rice germination at low temperature insecticides. In: Insecticides — Development of safer and more effective technologies /T. Trdan (ed.). IntechOpen, 2013: 417-426 (doi: 10.5772/53207).
- Shakir S.K., Irfan S., Akhtar B., Rehman S.U., Daud M.K., Taimur N., Azizullah A. Pesticide-induced oxidative stress and antioxidant responses in tomato (Solanum lycopersicum) seedlings. Ecotoxicology, 2018, 27(7): 919-935 (doi: 10.1007/s10646-018-1916-6).
- Çavuşoğlu K., Yalçın E., Türkmen Z., Yapar K., Sağır S. Physiological, anatomical, biochemical, and cytogenetic effects of thiamethoxam treatment on Allium cepa (amaryllidaceae) L. Environmental Toxicology, 2012, 27(11): 623-670 (doi: 10.1002/tox.20680).
- Маторин Д.Н., Рубин А.Б. Флуоресценция хлорофилла высших растений и водорослей. М.-Ижевск, 2012.
- Matorin D.N., Todorenko D.A., Seifullina N.Kh, Zayadan B.K., Rubin A.B. Effect of silver nanoparticles on the parameters of chlorophyll fluorescence and P700 reaction in the green alga Chlamydomonas reinhardtii. Microbiology, 2013, 82(6): 809-814 (doi: 10.1134/S002626171401010X).
- Kalaji H.M., Jajoo A., Oukarroum A., Brestic M., Zivcak M., Samborska I.A., Cetner M.D., Łukasik I., Goltsev V., Ladle R.J., Dąbrowski P., Ahmad P. The use of chlorophyll fluorescence kinetics analysis to study the performance of photosynthetic machinery in plants.In: Emerging Technologies and Management of Crop Stress Tolerance. V. 2 /P. Ahmad, S. Rasool (eds.). Elsevier, USA, 2014: 347-384 (doi: 10.1016/B978-0-12-800875-1.00015-6).
- Merlin J.C. Resonance Raman spectroscopy of carotenoids and carotenoid-containing systems. Pure and Applied Chemistry, 1985, 57(5): 785-792 (doi: 10.1351/pac198557050785).
- Тютяев Е.В., Шутова В.В., Максимов Г.В., Раденович Ч.Н., Гродзинский Д.М. Состояние фотосинтетических пигментов в листьях инбредных линий и гибридов кукурузы. Физиология растений и генетика, 2015, 47(2): 147-159.
- von Wettstein D. Chlorophyll-letale und der submikroskopische Formwechsel der Plastiden. Experimental Cell Research, 1957, 12(3): 427-506 (doi: 10.1016/0014-4827(57)90165-9).
- Strasser R.J., Tsimilli-Michael M., Qiang S., Goltsev V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochimica et Biophysica Acta (BBA) — Bioenergetics,2010, 1797(6-7): 1313-1326 (doi: 10.1016/j.bbabio.2010.03.008).
- Strasser R.J., Tsimilli-Michael M., Strivastava A. Analysis of the chlorophyll a fluorescence transient. In: Chlorophyll a fluorescence: advances in photosynthesis and respiration series, V. 19 /G.C. Papageorgiou, Govindjee (eds.). Springer, Dordrecht, 2004: 321-362 (doi: 10.1007/978-1-4020-3218-9_12).
- Stirbet A., Govindjee. Chlorophyll a fluorescence induction: a personal perspective of the thermal phase, the J-I-P rise. Photosynthesis Research, 2015, 113(1-3): 15-61 (doi: 10.1007/s11120-012-9754-5).
- Lazár D. The polyphasic chlorophyll afluorescence rise measured under high intensity of exciting light. Functional Plant Biology, 2006, 33(1): 9-30 (doi: 10.1071/FP05095).
- Nieva F.J.J., Castellanos E.M., Figueroa M.E., Gil F. Gas exchange and chlorophyll fluorescence of C3 and C4 saltmarsh species. Photosynthetica,1999, 36(3): 397-406 (doi: 10.1023/A:1007024019133).
- Živčák M., Olšovská K., Slamka P., Galambošová J., Rataj V., Shao H.B., Brestič M. Application of chlorophyll fluorescence performance indices to assess the wheat photosynthetic functions influenced by nitrogen deficiency. Plant, Soil and Environment, 2014, 60(5): 210-215 (doi: 10.17221/73/2014-PSE).
- Schansker G., Srivastava A., Govindjee, Strasser R.J. Characterization of the 820-nm transmission signal paralleling the chlorophyll a fluorescence rise (OJIP) in pea leaves. Functional Plant Biology, 2003, 30(7): 785-796 (doi: 10.1071/FP03032).
- Goltsev V., Chernev P., Zaharieva I., Strasser R.J. Delayed fluorescence in photosynthesis. Photosynthesis Research,2009, 101(2-3): 217-232 (doi: 10.1007/s11120-009-9451-1).
- Кэри П. Применение спектроскопии КР и РКР в биохимии /Под ред. Б.В. Локшина. М., 1985.
- Соловченко А.Е., Мерзляк М.Н. Оптическое экранирование как фотозащитный механизм растений. М., 2010.
- Preetha G., Stanley J. Influence of neonicotinoid insecticides on the plant growth attributes of cotton and okra. Journal of Plant Nutrition, 2012, 35(8): 1234-1245 (doi: 10.1080/01904167.2012.676134).
- Macedo W.R., Araújo D.K., de Camargo e Castro P.R. Unravelling the physiologic and metabolic action of thiamethoxam on rice plants. Pesticide Biochemistry and Physiology, 2013, 107(2): 224-249 (doi: 10.1016/j.pestbp.2013.08.001).
- Li L., Chen Xi., Zhang D., Pan X. Effects of insecticide acetamiprid on photosystem II (PSII) activity of Synechocystis sp. (FACHB-898). Pesticide Biochemistry and Physiology, 2010, 98(2): 300-304 (doi: 10.1016/j.pestbp.2010.06.022).