








УДК 635.64:575:631.52
ГЕНОТИПИЧЕСКАЯ ВАРИАБЕЛЬНОСТЬ В ПОТОМСТВЕ ТРАНСГЕННОГО МЕЖВИДОВОГО ГИБРИДА ТОМАТА. СООБЩЕНИЕ I. ПОЛУЧЕНИЕ СЕЛЕКЦИОННОГО МАТЕРИАЛА
Ю.В. ЧЕСНОКОВ1, Н.И. БОЧАРНИКОВА2, Л.В. ЕСАУЛОВА3
Выполняли фенотипический, генетико-селекционный и молекулярный RAPD-анализ генотипов потомства трансгенного межвидового гибрида томата Lycopersicon esculentum Mill. (сорт Факел) x Solanum pennellii Cor. Выявлена высокая генотипическая изменчивость изученных образцов как на молекулярном, так и на фенотипическом уровне. Отобранные формы томата представляют интерес для практической селекции.
Ключевые слова: межвидовой гибрид томата, коадаптированные блоки генов, спектр генотипической изменчивости, чужеродная экзогенная ДНК.
Для повышения общей и специфической адаптивности культивируемых видов растений требуется передать им от диких видов локусы хромосом или блоки коадаптированных генов, определяющие проявление количественных хозяйственно ценных признаков (1). Однако многие гены в этих блоках оказываются сцепленными с генами, обусловливающими нежелательные свой-ства. Потенциал генотипического варьирования в потомстве межвидовых гибридов полностью детерминирован особенностями их мейотической рекомбинации, а также взаимодействием генетических факторов, определяющих различия между компонентами скрещивания. Следовательно, для широкого использования генофонда диких видов необходимы эффективные методы, расширяющие спектр доступной селекционному отбору генетической и генотипической изменчивости, что представляет собой одну из важнейших задач современной селекции. В подобных исследованиях значительные перспективы в настоящее время связывают с применением экзогенной ДНК, что, во-первых, позволяет направленно передавать признаки от донора к реципиенту, во-вторых, способствует повышению изменчивости в потомстве.
Ранее мы получили трансгенный межвидовой гибрид томата Lycopersicon esculentum Mill. (сорт Факел) x Solanum pennellii Cor. (2). При скрещивании в качестве отцовской формы был выбран вид, который является источником ценных для культурного томата адаптивных признаков — холодоустойчивости, а также устойчивости к ряду вирусных и бактериальных заболеваний (3). Отличительная особенность гибрида заключалась в том, что в его генотип в процессе естественного опыления—оплодотворения (в прорастающие пыльцевые зерна) ввели чужеродную ДНК, контролирующую устойчивость к канамицину (2).
Целью настоящей работы была селекционно-генетическая и молекулярная оценка генотипов в расщепляющейся популяции трансгенного межвидового гибрида томата, несущего чужеродную экзогенную ДНК.
Методика. Материалом для исследований служили растения трансгенного межвидового гибрида томата L. esculentum Mill. (сорт Факел) x S. pennellii Cor. с детерминантой устойчивости к канамицину (2). Фенотипические особенности гибридных растений и их потомства оценивали в полевых условиях, сравнивая с общепринятыми описанными генотипами томатов (3, 4).
Тотальную ДНК выделяли из свежих листьев методом, предложенным Д.Б. Дороховым и Э. Клоке (5). ПЦР-анализ проводили в реакционной смеси (объем 25 мкл), содержащей 1xбуфер из коммерческого набора («Biomaster», Россия), 2мМ MgCl2, по 100 мкМ dATP, dCTP, dGTP и dTTP, 2,5 пмоль/мкл соответствующего праймера, 0,25 ед. Taq-полимеразы и 20 нг геномной ДНК. Для RAPD-анализа (random amplified polymorphism DNA) использовали олигонуклеотидные праймеры ОРА-09 (GGGTAACGCC) и ОРА-01 (CAGGCCCTTC) («Operon Technologies», США), праймеры 2-70-1 (GTCTCGTCGG), 4-70-1 (GCCCCTCTTG), 3-70-1 (TCCCTGTGCC), 2-60-1 (GAGTCTGTCG) («Genosys», США) и праймер Р-01 (AGCAGCGTCG), синтезированный в Центре «Биоинженерия» РАН (г. Москва).
Полимеразную цепную реакцию проводили в амплификаторе AM-PLY-250 («Biokom-Service», Россия) в следующем режиме: 3 мин при 94 °С, 1,5 мин при 37 °С, 1,5 мин при 72 °С — 1 цикл; 1 мин при 94 °С, 1,5 мин при 37 °С, 1,5 мин при 72 °С — 33 цикла; 1 мин при 94 °С, 1,5 мин при 37 °С, 15 мин при 72 °С — 1 цикл. Продукты амплификации разделяли электрофорезом в 2 % агарозном геле в 0,5xТВЕ-буфере, окрашивали бромистым этидием и фотодокументировали.
Статистическую обработку данных выполняли по Б.А. Доспехову (6).
Результаты. В предварительных исследованиях мы отобрали 9 трансгенных генотипов F1 L. esculentum x S. рennellii, устойчивых к канамицину (2, 7). Контролем служил межвидовой гибрид, полученный без использования экзогенной ДНК. Уже в F1 трансгенные гибридные растения по разнообразию окраски, массе и осемененности плодов не только различались между собой, но и отличались от контрольных. Окраска плодов варьировала от оранжево-красной до желтой и желто-зеленой. Для дальнейшей работы отобрали два генотипа F1, контрастных по окраске и массе плода. В последующих поколениях в расщепляющихся популяциях F2 наблюдалась вариабельность по форме и размеру куста, стерильности, завязываемости, окраске и массе плодов (рис. 1, А). Наряду с признаками, присущими исследуемым популяциям, у некоторых трансгенных генотипов проявились характерные индивидуальные особенности.
Так, если в контроле окраска плодов у всей популяции была желто-зеленой, то у трансгенных генотипов F1 — красной, оранжево-красной, оранжевой, желто-оранжевой, желтой и желто-зеленой. Два отобранных генотипа — 2-83(2) и 2-83(4) (F1) имели соответственно красный и желтый цвет плодов. При самоопылении в последующих четырех поколениях цвет плодов не изменялся, оставаясь красным и желтым, но при реципрокном скрещивании генотипы 2-83(2) и 2-83(4) в F2 давали плоды только красного цвета.
В красноплодной популяции 010 F2 гибрида 2-83(2) был обнаружен генотип 010/37 с очень сложной кистью, на которой насчитывалось до 300 цветков. Изучение характера наследования этого признака представляло очевидный интерес, однако при самоопылении растения в следующем поколении признак не проявился: сложная кисть «рассыпалась» на более мелкие (до 50-60 цветков). Нестабильность указанного признака прослеживалась и в дальнейшем, что проявлялось в расщеплении в последующих поколениях. Кроме того, появление такой кисти сопровождалось частичной стерильностью цветков (отмечалась гетеростилия и отчасти стерильность пыльцы), что затрудняло анализ.
В той же популяции (010) у генотипа 010/1, имевшего крупные красные плоды, при самоопылении в потомстве (F2) наблюдалось расщепление, близкое к 3:1 (χ2 = 2,84; 0,05 < Р < 0,20): примерно 3/4 растений формировали нормальные кисти, в которых завязывались плоды, 1/4 — давали соцветия с редуцированными или видоизмененными цветками, которые, как и следовало ожидать, были стерильны. При анализе потомства, полученного от самоопыления растений F2 с нормальными цветками, оказалось, что 1/3 генотипов F3 не расщеплялись и образовывали нормальные фертильные соцветия, тогда как 2/3 — давали растения как с нормальными, так и с редуцированными или видоизмененными соцветиями. Причем соотношение числа форм с нормальными и видоизмененными либо редуцированными соцветиями составляло примерно 3:1 (χ2= 1,93; 0,05 < Р < 0,20). Этот признак стабильно наследовался в поколениях (F4 и F5). Кроме того, удалось создать линии с видоизмененной и с редуцированной кистью, разделив генотипы. Оба указанных признака — моногенные рецессивы. Вследствие того, что в гомозиготной (рецессивной) форме генотипы, проявляющие эти признаки, стерильны, поддерживать и размножать такие мутантные формы можно только через гетерозиготу. Вновь приобретенный признак «видоизмененная кисть» морфологически несколько напоминает известную мутацию fa (8).
А |
Б |
|
|
В |
|
|
Рис. 1. Морфологические варианты у растений томата, полученные межвидовой гибридизацией Lycopersicon esculentum Mill. (сорт Факел) x Solanum pennellii Cor. с введением в генотип экзогенной ДНК, определяющей устойчивость к канамицину: А — различия по форме, размеру и окраске плодов в F2-F5; Б — «картофельный» тип листовой пластинки; В — образец 2-82(6) с сильным опушением (гены Wo или Ln). |



В желтоплодной популяции 011 F2 гибрида 2-83(4) были обнаружены генотипы 011(9) и 011(17), у которых на стадии проростков проявлялся признак, напоминающий известную мутацию с («картофельный» тип листа) (см. рис. 1, Б) (8). Установлено, что среди потомков F2 генотипа 011(9) растения с фенотипом «картофельный лист» составляли меньшинство (5 из 82), а полученное расщепление оказалось статистически недостоверным. То же отмечали и в последующих поколениях (F3 и F4). Признак «картофельный лист» наследовался случайным образом, не подчиняясь статистическим критериям и законам.
Совершенно иной характер наследования этого признака наблюдали у генотипа 011(17). Как и у генотипа 011(9), в популяции F2, полученной после самоопыления растения 011(17), были выявлены проростки как с нормальным, так и с «картофельным» типом листа при расщеплении примерно 3:1 (χ2 = 1,23; 0,20 < Р < 0,50). В популяциях, полученных в результате самоопыления растений каждого типа, формы с фенотипом «картофельный лист» в F2 дали в F3 растения только с этим признаком, а генотипы, имеющие нормальный лист в F2, в F3 разделились: у 1/3 не наблюдали расщепления (получены растения только с нормальным типом листа), у 2/3 вновь происходило расщепление по типу листа (нормальный и «картофельный»), причем в соотношении 3:1. В F4 самоопыленного потомства гибрида 011(17) наблюдались те же закономерности, что и в F2 и F3. Таким образом, по обнаруженному признаку «картофельный» тип листа отмечали разный характер наследования: у генотипа 011(17) — стабильный, рецессивный, моногибридный, гомозиготный, у 011(9) — нестабильный и случайный, что, на наш взгляд, указывает на различную природу указанного фенотипа у этих образцов.
Морфологический анализ последующих поколений (F5-F8) выявил высокую фенотипическую гетерогенность растений томата в популяциях и позволил выделить формы с признаками, не характерными для обоих родителей. Так, в F5 в расщепляющейся популяции 011 на стадии проростков был обнаружен генотип 2-82(6) с сильным опушением (см. рис. 1, В), внешне напоминающим проявление известных доминантных мутаций Wo или Ln (woolly и lanata — соответственно густое и чрезмерное опушение). На протяжении всей вегетация признак густого опушения сохранялся, причем на фоне других признаков. В то же время у образца 2-82(6) взрослое растение имело длинный стелющийся стебель (признак дикого вида), сложное (до 30-40 цветков) соцветие и красного цвета плоды с массой 50-60 г (признаки культурного вида). При самоопылении растения ни в F4, ни в последующих поколениях (вплоть до F8) признак густого опушения не изменялся и не исчезал. Полученные линии, имеющие такое опушение, стабильно его наследовали.
В то же время в поколениях F4-F8 происходило изменение ряда других признаков. Так, уже в F4 появились растения с компактным типом куста, свойственным культурному виду, в том же поколении — растения с желто-оранжевыми и фиолетовыми плодами, хотя масса плода при этом практически не изменилась. Произошло также разделение по типу соцветия: первый характеризовался 30-40 цветками на кисти, второй — только 8-10. В F5 растения с компактным типом куста его сохранили, и в последующих поколениях не давая расщепления по длине стебля. Растения с длинным стеблем в F5 дали расщепление на формы с густым опушением и растения обычного типа. В F6-F8 растения с густым опушением не утратили этот признак, а растения с обычным типом куста в F6 вновь дали растения без густого опушения, однако в F7 самоопыленного потомства густое опушение появилось опять и наследовалось в F8. Окраска плодов в поколениях F5-F8 также не изменялась и сохранялась у самоопыленного потомства как красноплодных, так и желто-оранжевых генотипов. Этот признак наследовался независимо от других.
Наибольшие изменения по степени фенотипического проявления наблюдались для признака «тип соцветия». По нему оба класса образцов (с 30-40 и 8-10 цветками на кисти) в F5-F8, несмотря на строгое самоопыление, давали самое разнообразное потомство — генотипы как с простой (4-5 цветков), так и со сложной (до 40 цветков и более) кистью. Причем признак был нестабильным, а его проявление — непредсказуемым даже внутри одной семьи.
Качественные характеристики цветка (структура и морфология) также варьировали. В частности, в самоопыленном потомстве F5 появились генотипы с редуцированной кистью, в F6 и F8 — формы с кожистым типом цветка, у которого все части, включая чашелистики, имели плотный эпидермис. Кроме того, такие цветки несли недоразвитые тычинки и (в отличие от обычных) были стерильны.
Разная степень стерильности цветков на кисти отмечалась и у других генотипов той же семьи. Так, завязываемость плодов варьировала от 0 до 100 % практически во всех поколениях (F4-F8) исследованных форм с признаком «густое опушение».
Нам не удалось обнаружить корреляционные связи между густым опушением, типом куста и соцветия, стерильностью и завязываемостью плодов, а также определить характер наследования: проведенный статистический анализ не выявил каких-либо закономерностей и подчиненности законам Менделя либо другим биологическим законам наследования признаков.
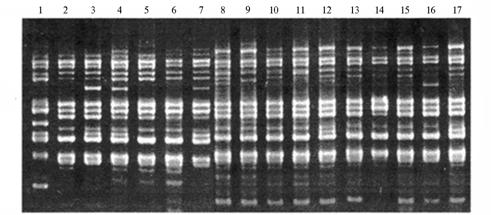
Необходимо отметить, что генотип F1 исходно содержал в геноме последовательности активной трансгенной ДНК, что было установлено блот-дот-гибридизацией с последующим определением активности NPTII (неомицинфосфотрансфераза, определяющая устойчивость к канамицину как селективному маркеру) (2). Проведенный дополнительный сравнительный RAPD-анализ изучаемых популяций выявил, что в потомстве трансгенного гибрида преобладали RAPD-спектры, близкие к таковым у L. escu-lentum, и лишь у некоторых генотипов имелись зоны амплифицированной ДНК, характерные для S. pennellii, что подтверждает их гибридную природу. У ряда генотипов отсутствовали некоторые зоны ДНК, типичные для L. esculentum, и регистрировались зоны, несвойственные для родительских форм (рис. 2). К тому же у значительной части гибридов были обнаружены новые зоны ампликонов, что свидетельствует о значительной перестройке генома.
|
Рис. 2. RAPD-спектры в потомстве у томата, полученного при межвидовой гибридизации Lycopersicon esculentum Mill. (сорт Факел) x Solanum pennellii Cor. с введением экзогенной ДНК, определяющей устойчивость к канамицину: 1 — S. pennellii; 2 — L. esculentum; 3-17 — F5 самоопыленного полового потомства (праймер 4-70-1, фирма «Genosys», США). |
Итак, использование чужеродной экзогенной ДНК при межвидовом скрещивании приводит к расширению спектра генотипической вариабельности у томата и позволяет в короткие сроки получить гибридные генотипы с ценными хозяйственными признаками (на достижение подобного результата обычными гибридологическими методами необходимо затратить несколько лет) (9-12). Выделенный генетический материал, несомненно, представляет большой интерес для генетики и селекции, поскольку в популяциях, помимо обычных рекомбинантных, имеются новые трансгрессивные и необычные мутантные формы со свойствами, не присущими обоим родителям.
Л И Т Е Р А Т У Р А
1. Ж у ч е н к о А.А. Адаптивная система селекции растений (эколого-генетические основы). Т. I-II. М., 2001.
2. Ч е с н о к о в Ю.В. Наследственные изменения, вызванные переносом экзогенной ДНК в высшие растения посредством прорастающей пыльцы. Автореф. докт. дис. СПб, 2000.
3. Ж у ч е н к о А.А., Г л у щ е н к о Е.Я., А н д р ю щ е н к о В.К., Б а л а ш о в а Н.Н., С а м о в о л А.П., М е д в е д е в В.В. Дикие виды и полукультурные разновидности томатов и их использование в селекции. Кишинев, 1974.
4. Ж у ч е н к о А.А. Генетика томатов. Кишинев, 1973.
5. Д о р о х о в Д.Б., К л о к е Э. Быстрая и экономичная технология RAPD анализа растительных геномов. Генетика, 1997, 33(4): 443-450.
6. Д о с п е х о в Б.А. Методика полевого опыта. М., 1973.
7. С е д о в Г.И., Ч е с н о к о в Ю.В. Отбор трансформантов in vitro на стадии незрелых зародышей. Генетика, 1995, 31(4): 499-503.
8. Б о ч а р н и к о в а Н.И., К о з л о в а В.М. Мутантные формы томатов. Кишинев, 1992.
9. Ж у ч е н к о А.А., К о р о л ь А.Б. Рекомбинация в эволюции и селекции. М., 1985.
10. Ж у ч е н к о А.А. Эколого-генетические основы адаптивной системы селекции растений. С.-х. биол., 2000, 3: 3-29.
11. Г е р ш е н з о н С.М., А л е к с а н д р о в Ю.Н., М а л ю т а С.С. Мутагенное действие ДНК и вирусов у дрозофилы. Киев, 1975.
12. M o r t o n R., H o o y k a a s P.J.J. Gene replacement. Mol. Breeding, 1995, 1(2): 123-132.
Yu.V. Chesnokov1, N.I. Bocharnikova2, L.V. Esaulova3
The authors made the phenotypic, genetic-selective and molecular RAPD-analysis of genotypes of progeny of transgenic interspecific hybrid of tomato of Lycopersicon esculentum Mill. (the Fakel variety) x Solanum pennellii Cor. The high genetic variability was revealed in studied variants both on molecular and phenotypic level. Selected tomato forms are of interest for practical breeding.
Keywords: interspecific hybrid of tomato, coadapted blocks of genes, spectrum genotypic variability, alien exogenous DNA.
1ГНУ Всероссийский НИИ растениеводства |
Поступила в редакцию |




