








УДК 635.649:[631.524.7+631.524.84]:575.113.2:631.522/.524
ОБ ИДЕНТИФИКАЦИИ КЛЮЧЕВЫХ АЛЛЕЛЕЙ ХОЗЯЙСТВЕННО ЦЕННЫХ ПРИЗНАКОВ У ОВОЩНОГО ПЕРЦА Capsicum annuum L. РЕГРЕССИОННО-КЛАСТЕРНЫМ АНАЛИЗОМ
О.О. ТИМИНА1, А.C. РЯБОВА2
С использованием родительских форм овощного перца (сорта Добрыня Никитич, Колобок, Прометей, а также линии Л 48 и Л 49, соответственно 1-й, 2-й, 3-й, 4-й и 5-й генотип) и их гибридов F1, полученных в диаллельных скрещиваниях по схеме [х р(р + 1)], дифференцировали изучаемые генотипы на высоко-, средне- и малоидентичные. Полученные данные позволяют определить взаимосвязь между степенью идентичности ключевых аллелей хозяйственно ценных признаков, подбором пар для скрещивания и эффектом гетерозиса.
Ключевые слова: количественные признаки, регрессионно-кластерный анализ, овощной перец.
Известно, что у растений большая часть хозяйственно ценных признаков детерминируется полигенно. Идентификация полигенов с целью получения принципиально новых форм — одна из основных практических задач. Однако при положительном опыте обогащения традиционной парадигмы селекции положениями молекулярно-генетического анализа (1-3) отмечается явная недостаточность как набора уже исследованных полигенов, так и методов их изучения (4). Низкий уровень полиморфизма у Capsicum annuum служит причиной того, что молекулярной маркировке поддается около 25 % изучаемых качественных и количественных признаков (5). Кроме того, стоимость таких анализов достаточно высока. Поэтому общедоступной и актуальной методологической основой изучения количественных признаков в практической селекции остается генетико-статис-тический (6, 7), в частности регрессионно-кластерный (8, 9), анализ, позволяющий оценить материал не только с неаллельными генными взаимодействиями и материнским эффектом, но и дифференцировать доминантные аллели по норме реакции.
Целью наших исследований была идентификация ключевых аллелей хозяйственно ценных признаков у овощного перца Capsicum annuum с применением регрессионно-кластерного анализа.
Методика. На основе пяти генотипов перца (сорта и линии Добрыня Никитич, Колобок, Л 49, Прометей и Л 48 — соответственно 1-й, 2-й, 3-й, 4-й и 5-й генотипы) проводили диаллельные скрещивания по схеме [x р(р + 1)]. Скрещивания выполняли в необогреваемых пленочных теплицах в центральной части Приднестровского региона в 2004-2005 годах. В следующем сезоне растения родительских форм и гибридов F1 выращивали, следуя общепринятым рекомендациям для культуры. В соответствии со схемой случайных рендомизированных блоков 50-суточную рассаду высаживали (в двух повторностях по 10 растений в каждом варианте) в открытый грунт и необогреваемую пленочную теплицу. Вариансы (Vr ) и ковариансы (Wr ) рассчитывали методом Hayman с соавт. (цит. по ссылке 10). Степень варьирования величины Wr + Vr у изучаемых генотипов в зависимости от условий культивирования оценивали трехфакторным дисперсионным анализом по общепринятой методике (10). Регрессионно-кластерный анализ проводили согласно описанию (8, 9), применяя программный пакет Statistica 6.0. Для выравнивания полученных данных использовали линейную нормализацию величин Wr + Vr каждого признака по формуле:
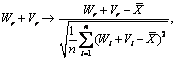
где
Оптимальность кластеризации устанавливали, сравнивая результаты, полученные разными методами — итерационными (определение k-средних по Мак-Кину с использованием программы Statistica 6.0, метод нечеткой кластеризации) (11) и иерархическим. Генотипы идентифицировали внутри кластера, разбивая параметры сопряженных признаков на родственные сгущения. Для большей наглядности полученные сгущения изображали в виде расшифрованных схем с одинаковым обозначением объектов, входящих в один кластер. Число наблюдений (Cases) в кластере подсчитывали по формуле:
C = n • n1 • n2,
где n — число признаков, объединенных в общий кластер, n1— число условий, в которых выращивались растения, n2— число исследуемых генотипов. Степень идентичности (%) подсчитывали по совпадению или несовпадению обозначения соответствующих признаков при наложении схем сравниваемых генотипов. О плейотропном эффекте гена судили по фенотипическому эффекту в благоприятных условиях (защищенный грунт), используя те же расшифрованные схемы.
Результаты. Х а р а к т е р и с т и к а и о п т и м а л ь н о с т ь
к л а с т е р и з а ц и и. Диаллельный анализ пяти генотипов перца по 18 изучаемым признакам показал, что по большинству отмечались как неаллельные взаимодействия, так и зависимость экспрессии от условий выращивания (табл. 1). Полученные результаты совпадают с опубликованными данными (5), согласно которым у генотипов перца проявление хозяйственно ценных признаков не описывалось аддитивно-доминантной моделью по основным параметрам, однако отмечалась очень хорошая корреляция между молекулярными и биометрическими показателями. Поэтому в дальнейшем использовали кластерный анализ значений Wr + Vr, показывающих наличие у генотипов доминантных аллелей (10), для 16 признаков, изученных одновременно в двух различных условиях.
| 1. Значения объединенных коэффициентов регрессии и дисперсионного анализа экспрессии хозяйственно ценных признаков у изученных генотипов овощного перца (сорта и линии Добрыня Никитич, Колобок, Л 49, Прометей и Л 48) в разных условиях выращивания (по данным диаллельных скрещиваний) | ||||||||||
Признак |
Защищенный грунт |
Открытый грунт |
||||||||
b1±m |
t1 отличие от 0 |
t2 отличие от 1 |
Wr - Vr |
b1±m |
t1 отличие от 0 |
t2 отличие от 1 |
Wr - Vr |
|||
F |
P |
F |
P |
|||||||
Продолжительность: |
|
|
|
|
|
|
|
|
|
|
I фенофазы |
0,9705±0,2303 |
4,214 |
0,128 |
1,850 0,008 |
> 0,05 |
1,3060±0,3813 |
3,425 |
-0,800 |
5,697 |
< 0,05 |
II фенофазы |
0,6242±0,1510 |
4,134 |
2,489 |
9,376 |
< 0,05 |
0,6790±0,0960 |
7,073 |
3,340 |
1,383 |
> 0,05 |
III фенофазы |
1,0062±0,1637 |
6,147 |
-0,038 |
3,677 |
> 0,05 |
1,0480±0,1704 |
6,150 |
-0,282 |
3,471 |
> 0,05 |
вегетационного периода |
1,2249±0,2496 |
4,907 |
-0,901 |
5,813 |
< 0,05 |
0,9021±0,2350 |
3,839 |
0,417 |
5,432 |
< 0,05 |
Высота куста |
0,7454±0,1416 |
5,264 |
1,798 |
16,750 |
< 0,05 |
0,6900±0,1724 |
4,000 |
1,800 |
8,134 |
< 0,05 |
Размер плода: |
|
|
|
|
|
|
|
|
|
|
длина |
0,9210±0,0596 |
15,453 |
1,326 |
6,224 |
< 0,05 |
0,8190±0,0560 |
14,625 |
3,232 |
1,058 |
> 0,05 |
диаметр |
0,2062±0,1976 |
1,043 |
4,017 |
74,340 |
< 0,05 |
0,9492±0,0801 |
11,850 |
0,634 |
1,435 |
> 0,05 |
Индекс плода |
0,9524±0,0654 |
14,563 |
0,728 |
11,046 |
< 0,05 |
0,7320±0,0580 |
12,620 |
4,620 |
2,260 |
> 0,05 |
Число камер в плоде |
0,8766±0,0692 |
12,514 |
1,936 |
3,107 |
> 0,05 |
0,4120±0,0920 |
4,478 |
6,390 |
0,675 |
> 0,05 |
Толщина стенки плода |
0,7208±0,0861 |
8,372 |
3,243 |
0,078 |
> 0,05 |
0,9160±0,0504 |
18,170 |
1,667 |
1,757 |
> 0,05 |
Масса плода: |
|
|
|
|
|
|
|
|
|
|
средняя с растения |
0,2608±0,1219 |
2,139 |
6,064 |
21,100 |
< 0,05 |
0,9022±0,0690 |
13,070 |
1,420 |
4,945 |
> 0,05 |
товарного |
0,2603±0,1120 |
2,341 |
6,652 |
22,856 |
< 0,05 |
0,9300±0,0650 |
14,310 |
1,077 |
0,477 |
> 0,05 |
Число плодов с растения |
0,1450±0,1100 |
1,318 |
7,770 |
3,890 |
> 0,05 |
0,8113±0,1208 |
6,716 |
1,562 |
0,856 |
> 0,05 |
Урожай с растения: |
|
|
|
|
|
|
|
|
|
|
общий |
-0,0230±0,0547 |
0,420 |
17,860 |
6,769 |
< 0,05 |
0,7500±0,2510 |
2,988 |
0,996 |
0,153 |
> 0,05 |
товарный |
0,0655±0,0488 |
1,340 |
19,150 |
5,270 |
< 0,05 |
0,4490±0,3190 |
1,407 |
1,727 |
0,840 |
> 0,05 |
Увядание в монокультуре |
0,6066±0,1790 |
3,389 |
2,198 |
5,148 |
< 0,05 |
0,3030±0,2487 |
1,218 |
2,800 |
2,279 |
> 0,05 |
Гниль плодов |
0,6520±0,2180 |
2,990 |
1,596 |
1,616 |
> 0,05 |
0,6130±0,1511 |
4,057 |
2,560 |
4,069 |
> 0,05 |
Прорастание при низких температурах. |
0,9080±0,2900 |
3,131 |
0,317 |
2,326 |
> 0,05 |
|
|
|
|
|
Поражение вертициллезом при искусственном заражении |
0,7740±0,1086 |
7,127 |
2,081 |
1,595 |
> 0,05 |
|
|
|
|
|
П р и м е ч а н и е. При Fфакт. < Fстат. Р > 0,05; при Fфакт. > Fстат. Р < 0,05; b1±m — объединенный коэффициент регрессии. Для F и Р приведены значения для вариантов и повторностей (соответственно над и под чертой). |
||||||||||
Иерархическая кластеризация нормализованных величин параметра Wr + Vr по всему массиву данных (табл. 2, рис. 1, А) свидетельствовала о
|
Рис. 1. Дендрограммы, ха-рактеризующие идентичность изученных генотипов в разных условиях выращивания (А) и 16 хозяйственно ценных признаков (Б) у овощного перца, построенные на основе нормализованных значений Wr + Vr (по результатам диаллельных |
скрещиваний, см. табл. 2; критерий объединения — полная связь; функция расстояния — 1-коэффициент парной корреляции Пирсона). |
|
следующей группировке генотипов по кластерам: 1-й кластер — Прометей (защищенный грунт), Прометей (открытый грунт), Колобок (открытый грунт); 2-й — Л 49 (защищенный грунт), Колобок (защищенный грунт); 3-й — Добрыня Никитич (открытый грунт), Л 49 (открытый грунт); 4-й — Добрыня Никитич (защищенный грунт), Л 48 (защищенный грунт), Л 48 (открытый грунт). Аналогичная кластеризация по 16 признакам (см. табл. 2, рис. 2, Б) выявила следующие сгущения: 1-е — длина плода, индекс плода; 2-е — диаметр плода; 3-е — высота куста; 4-е — гниль плода; 5-е — средняя масса плода, товарная масса плода; 6-е — продолжительность III фенофазы, продолжительность II фенофазы; 7-е — увядание в монокультуре, толщина стенки плода; 8-е — число камер в плоде, продолжительность I фенофазы; 9-е — товарный урожай с растения, общий урожай с растения, число плодов с растения.
По условию кластеризации объекты из одного кластера имеют сход-
| 2. Итоговые и нормализованные значения величины Wr + Vr для 16 хозяйственно ценных признаков у изученных генотипов овощного перца (сорта и линии Добрыня Никитич, Колобок, Л 49, Прометей и Л 48) в разных условиях выращивания (по данным диаллельных скрещиваний) | ||||||||||
Признак |
Генотип |
|||||||||
1-й |
2-й |
3-й |
4-й |
5-й |
||||||
1 |
2 |
1 |
2 |
1 |
2 |
1 |
2 |
1 |
2 |
|
И т о г о в ы е з н а ч е н и я |
||||||||||
Продолжительность: |
|
|
|
|
|
|
|
|
|
|
I фенофазы |
-0,850 |
0,175 |
7,750 |
24,750 |
5,375 |
0,275 |
5,125 |
6,350 |
5,775 |
16,575 |
II фенофазы |
0,725 |
0,825 |
6,200 |
4,875 |
0,775 |
3,050 |
4,750 |
7,850 |
1,400 |
2,000 |
III фенофазы |
1,025 |
3,200 |
3,300 |
5,500 |
0,775 |
3,050 |
7,575 |
11,625 |
1,525 |
5,300 |
Высота куста |
345,360 |
347,550 |
67,081 |
44,425 |
53,058 |
1,075 |
25,539 |
59,250 |
137,467 |
212,050 |
Размер плода: |
|
|
|
|
|
|
|
|
|
|
длина |
3,059 |
3,295 |
0,716 |
2,084 |
3,866 |
4,810 |
4,353 |
4,831 |
4,138 |
5,102 |
диаметр |
0,786 |
0,870 |
0,361 |
0,447 |
0,460 |
0,897 |
0,810 |
0,406 |
0,632 |
0,436 |
Индекс плода |
0,191 |
0,156 |
0,037 |
0,021 |
0,191 |
0,305 |
0,156 |
0,194 |
0,201 |
0,233 |
Число камер в плоде |
0,195 |
0,050 |
0,082 |
0,236 |
0.103 |
0,115 |
0,119 |
0,022 |
0,068 |
0,154 |
Толщина стенки плода |
0,014 |
0,014 |
0,028 |
0,013 |
0,021 |
0,017 |
0,016 |
0,015 |
0,009 |
0,008 |
Масса плода: |
|
|
|
|
|
|
|
|
|
|
средняя с растения |
0,0004 |
0,0006 |
0,0002 |
0,0010 |
0,0002 |
0,0008 |
0,0007 |
0,0006 |
0,0003 |
0,0004 |
товарного |
0,0006 |
0,0006 |
0,0003 |
0,0010 |
0,0004 |
0,0008 |
0,0008 |
0,0006 |
0,0003 |
0,0003 |
Число плодов с растения |
0,162 |
3,776 |
0,423 |
1,026 |
1,257 |
3,269 |
0,282 |
1,317 |
1,477 |
2,283 |
Урожай с растения: |
|
|
|
|
|
|
|
|
|
|
общий |
0,0062 |
0,0582 |
0,0030 |
0,0300 |
0,0010 |
0,0680 |
0,0068 |
0,0089 |
0,0007 |
0,0149 |
товарный |
0,0033 |
0,0627 |
0,0015 |
0,0355 |
0,0009 |
0,0564 |
0,0024 |
0,0080 |
0,0057 |
0,0163 |
Увядание в монокультуре |
0,484 |
0,104 |
0,700 |
0,211 |
1,375 |
0,076 |
0,830 |
0,074 |
0,300 |
0,126 |
Гниль плодов |
0,327 |
0,354 |
0,333 |
0,491 |
0,206 |
0,300 |
1,452 |
0,088 |
0,055 |
0,113 |
Н о р м а л и з о в а н н ы е з н а ч е н и я |
||||||||||
Продолжительность: |
|
|
|
|
|
|
|
|
|
|
I фенофазы |
-0,875 |
-1,004 |
2,217 |
0,078 |
-0,862 |
-0,221 |
-0,098 |
-0,252 |
1,188 |
-0,171 |
II фенофазы |
0,952 |
0,991 |
0,641 |
0,162 |
0,077 |
0,972 |
1,812 |
0,592 |
0,490 |
0,726 |
III фенофазы |
0,324 |
0,972 |
0,361 |
0,294 |
0,369 |
1,046 |
2,186 |
0,979 |
0,302 |
0,823 |
Высота куста |
1,689 |
1,672 |
0,657 |
0,482 |
0,992 |
0,590 |
0,542 |
0,803 |
0,641 |
0,063 |
Размер плода: |
|
|
|
|
|
|
|
|
|
|
длина |
0,239 |
0,410 |
1,116 |
2,107 |
0,858 |
0,174 |
0,873 |
0,527 |
1,069 |
0,371 |
диаметр |
1,224 |
0,828 |
-0,771 |
-1,177 |
1,352 |
-0,710 |
-0,965 |
0,941 |
-0,823 |
0,101 |
Индекс плода |
-0,147 |
0,265 |
-1,737 |
-1,549 |
1,608 |
0,265 |
0,300 |
-0,147 |
0,760 |
0,383 |
Число камер в плоде |
-0,980 |
1,226 |
1,850 |
-0,493 |
0,009 |
-0,173 |
-1,406 |
0,070 |
0,602 |
-0,706 |
Толщина стенки плода |
-0,222 |
-0,326 |
-0,430 |
2,171 |
0,264 |
0,957 |
-0,083 |
0,090 |
-1,297 |
-1,124 |
Масса плода: |
|
|
|
|
|
|
|
|
|
|
средняя с растения |
0,301 |
-0,451 |
1,806 |
-1,204 |
1,053 |
-1,204 |
0,301 |
0,677 |
-0,451 |
-0,828 |
товарного |
0,122 |
0,122 |
1,754 |
-1,101 |
0,938 |
-0,693 |
0,122 |
0,938 |
-1,101 |
-1,101 |
Число плодов с растения |
1,825 |
-1,108 |
-0,407 |
-0,896 |
1,414 |
-0,219 |
-0,171 |
-1,011 |
0,613 |
-0,041 |
Урожай с растения: |
|
|
|
|
|
|
|
|
|
|
общий |
1,569 |
-0,554 |
0,418 |
-0,685 |
1,969 |
-0,766 |
-0,444 |
-0,53 |
-0,199 |
-0,779 |
товарный |
1,835 |
-0,675 |
0,686 |
-0,751 |
1,569 |
-0,776 |
-0,476 |
-0,713 |
-0,126 |
-0,573 |
Увядание в монокультуре |
-0,758 |
0,131 |
-0,508 |
0,636 |
-0,824 |
2,216 |
-0,828 |
0,941 |
-0,707 |
-0,300 |
Гниль плодов |
-0,044 |
-0,111 |
0,295 |
-0,096 |
-0,178 |
-0,411 |
-0,704 |
2,677 |
-0,642 |
-0,785 |
П р и м е ч а н и е. Обозначения генотипов (1-й, 2-й, 3-й, 4-й и 5-й) см. в разделе «Методика»; 1 и 2 — соответственно открытый и защищенный грунт. |
||||||||||
ную норму реакции и, соответственно, тождественный набор доминантных аллелей. Следовательно, сравнив состав кластеров, можно определить сопряженность конкретных признаков у генотипов, идентичность генотипов по доминантным аллелям, детерминирующим признаки, и стабильность
|
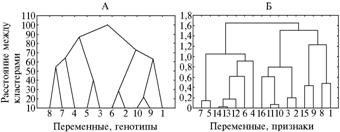
Рис. 2. Дендрограммы идентичности доминантных аллелей хозяйственно ценных признаков у изученных генотипов овощного перца в разных условиях выращивания |
(построены на основе нормализованных значений Wr + Vr, полученных при диаллельных скрещиваниях, см. табл. 2). А-Д — соответственно кластеры длина плода—индекс плода (для 20 случаев, критерий средней арифметической простой, евклидово расстояние); общий урожай с растения—товарный урожай с растения—число плодов с растения (для 30 случаев, критерий дальнего соседа, полная связь, евклидово расстояние); диаметр плода (для 10 случаев, критерий дальнего соседа, полная связь, евклидово расстояние); высота куста (для 10 случаев, критерий средней арифметической простой, евклидово расстояние); гниль плодов (для 10 случаев, критерий средней арифметической простой, евклидово рас- стояние). Описание признаков, контролируемых аллелями, см. на рисунке 4. |
|
их экспрессии, а также общую функцию, что не всегда возможно при использовании других методов, включая молекулярные. При этом конкурентные взаимоотношения генотипов не учитывали ввиду заведомой нивелировки плотности посадки растений на выбранных фонах.
| 3. Результаты кластеризации хозяйственно ценных признаков и генотипов овощного перца (сорта и линии Добрыня Никитич, Колобок, Л 49, Прометей и Л 48) в зависимости от условий выращивания, выполненной разными статистическими методами на основе нормализованных значений параметра Wr + Vr для 16 признаков | ||
Номер кластера |
Показатель |
Состав кластера |
П р и з н а к и, м е т о д н е ч е т к о й
(параметр останова = 0,001; экспоненциальный вес = 1,300; |
||
1-й |
0,999824а |
Длина плода |
2-й |
1а |
Диаметр плода |
3-й |
1а |
Высота куста |
4-й |
1а |
Гниль плодов |
5-й |
0,999954а |
Масса товарного плода |
6-й |
0,999487а |
Продолжительность II фенофазы |
7-й |
0,995813а |
Толщина стенки плода |
8-й |
0,991196а |
Продолжительность I фенофазы |
9-й |
0,999940а |
Товарный урожай с растения |
Г е н о т и п ы, м е т о д k-м е т р и к и п о М а к-К и н у |
||
1-й |
0,777730б |
Сорт Колобоко |
2-й |
0,410360б |
Сорт Колобокз |
3-й |
0,486175б |
Сорт Добрыня Никитичо |
4-й |
0,628852б |
Добрыня Никитичз |
П р и з н а к и, м е т о д k-м е т р и к и п о М а к-К и н у |
||
1-й |
0,255483б |
Длина плода |
2-й |
0 |
Диаметр плода |
3-й |
0 |
Высота куста |
4-й |
0 |
Гниль плодов |
5-й |
0,176441б |
Масса товарного плода |
6-й |
0,300459б |
Продолжительность II фенофазы |
7-й |
0,445215б |
Толщина стенки плода |
8-й |
0,465017б |
Продолжительность I фенофазы |
9-й |
0,185290б |
Товарный урожай с растения |
П р и м е ч а н и е. а, б — соответственно степень принадлежности и расстояние от центра кластера; о, з — соответственно открытый и защищенный грунт. |
||
Для определения оптимальности кластеризации не удалось использовать дискриминан-тный анализ (11, 12), так как были получены кластеры, содержащие по одному объекту, и экспериментальная величина толерантности выходила за пределы, принятые по умолчанию. Поэтому оптимальность кластеризации оценивали, дополнительно разбивая полученный массив данных итерационными методами. Сопоставление результатов выявило очень хорошее совпадение данных (100 %), свидетельствующее об относительной оптимальности разбивки на кластеры разными способами — при применении метрики k-средних Мак-Кина для генотипов с учетом разделения на 4 кластера и иерархической кластеризации, а также при разбивке параметра Wr + Vr для 16 признаков на 9 кластеров с помощью ал-горитма нечеткой кластеризации, метрики k-средних Мак-Кина и иерархического метода (см. рис. 1, см. табл. 2, 3). Следовательно, полученные кластеры достаточно устойчивы.
Результаты кластеризации параметра Wr + Vr генотипов продемонстрировали, что у Л 48 и сорта Прометей значения параметров экспрессии для большинства признаков совпадают или очень близки в условиях как открытого, так и защищенного грунта. Вероятно, эти генотипы можно рассматривать в качестве универсальных доноров стабильной экспрессии комплекса хозяйственно ценных признаков.
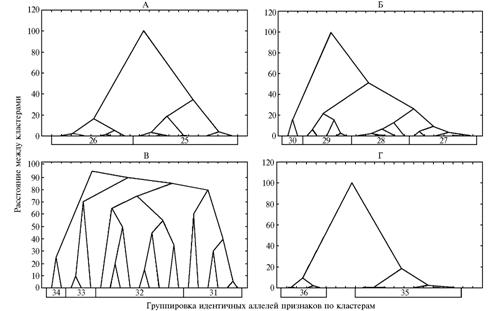 |
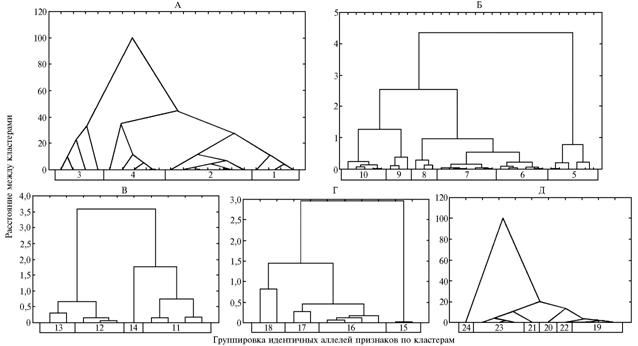
Рис. 3. Дендрограммы идентичности доминантных аллелей хозяйственно ценных признаков у изученных генотипов овощного перца в разных условиях выращивания (построены на основе нормализованных значений Wr + Vr, полученных при диаллельных скрещиваниях, см. табл. 2). А-Г — соответственно кластеры средняя масса—товарная масса плода (для 20 случаев, метод Уорда, евклидово расстояние); продолжительность II и III фенофаз (для 20 случаев, критерий средней арифметической простой, евклидово расстояние); увядание в условиях монокультуры—толщина стенки плода (для 20 случаев, критерий среднего соседа без учета числа объектов в кластере, степенное); число камер в плоде—продолжительность I фенофазы (для 20 случаев, метод Уорда, квадрат евклидовой метрики). Описание признаков, контролируемых аллелями, см. на рисунке 4. |
Изучение проявления 16 признаков для указанного генетического фона выявило сопряженность следующих из них: длина плода и индекс плода; продолжительность II и III фенофаз; увядание и толщина стенки плода; средняя и товарная масса плода; число камер в плоде и продолжительность I фенофазы; товарный, общий урожай и число плодов на растении. Обнаруженная сопряженность признаков позволила провести их дальнейшую внутрикластерную идентификацию на основе соответствующих сгущений.
Х а р а к т е р и с т и к а к л а с т е р о в. 1-й к л а с т е р (д л и н а
п л о д а—и н д е к с п л о д а).При дальнейшей группировке объектов этого кластера (см. табл. 3, рис. 1, Б) проявились четыре более мелких устойчивых сгущения, характеризующих экспрессию признаков (см. рис. 2, А). Согласно схеме состава доминантных аллелей в этом кластере (см. рис. 4), наиболее дифференцированная экспрессия признаков отмечалась у сорта Прометей, наиболее стабильная — у сорта Колобок. В контроле признаков длина плода и индекс плода участвуют четыре различных аллеля с плейотропным фенотипическим эффектом. Плейотропизм проявился у трех (2-й, 3-й и 5-й) из пяти проанализированных генотипов. Степень функциональной идентичности с учетом экспрессии в разных условиях произрастания у 1-го и 2-го генотипов составила 0 %; 1-го и 3-го — 25,0 %; 1-го и 4-го — 0 %; 1-го и 5-го — 25,0 %; 2-го и 3-го — 0 %; 2-го и 4-го — 0 %; 2-го и 5-го — 0 %; 3-го и 4-го — 75,0 %; 3-го и 5-го — 100 %; 4-го и 5-го — 50,0 %.
2-й к л а с т е р (д и а м е т р п л о д а). Кластер формировал четыре устойчивых сгущения (см. рис. 2, В, рис. 4). Стабильная экспрессия признака отмечалась только у сорта Добрыня Никитич. Диаметр плода контролируется четырьмя аллелями. Степень функциональной идентичности с учетом экспрессии в разных условиях произрастания: 1-го и 2-го генотипов — 0 %; 1-го и 3-го — 50,0 %; 1-го и 4-го — 50,0 %; 1-го и 5-го — 0 %; 2-го и 3-го — 0 %; 2-го и 4-го — 0 %; 2-го и 5-го — 50,0 %; 3-го и 4-го — 0 %; 3-го и 5-го — 0 %; 4-го и 5-го — 0 %.
3-й к л а с т е р (в ы с о т а к у с т а). В кластере выявлялись четыре устойчивых сгущения (см. рис. 2, Г, рис. 4). Стабильную экспрессию признака отмечали у сортов Добрыня Никитич, Колобок и линии Л 48. Высота куста контролируется четырьмя аллелями. Степень функциональной идентичности с учетом экспрессии в разных условиях произрастания: 1-го и 2-го генотипов — 0 %; 1-го и 3-го — 0 %; 1-го и 4-го — 0 %; 1-го и 5-го — 0 %; 2-го и 3-го — 50,0 %; 2-го и 4-го — 50,0 %; 2-го и 5-го — 0 %; 3-го и 4-го — 0 %; 3-го и 5-го — 0 %; 4-го и 5-го — 0 %.
4-й к л а с т е р (г н и л ь п л о д о в). Кластер преобразовывался в шесть устойчивых сгущений (см. рис. 2, Д, рис. 4). Стабильную экспрессию признака выявили у сорта Добрыня Никитич и линии Л 48. Признак контролируется шестью аллелями. Степень функциональной идентичности с учетом экспрессии в разных условиях произрастания: 1-го и 2-м генотипов — 50,0 %; 1-го и 3-го — 0 %; 1-го и 4-го — 0 %; 1-го и 5-го — 0 %; 2-го и 3-го — 0 %; 2-го и 4-го — 0 %; 2-го и 5-го — 0 %; 3-го и 4-го — 0 %; 3-го и 5-го — 0 %; 4-го и 5-го — 50,0 %.
5-й к л а с т е р (
с р е д н я я м а с с а п л о д а—т о в а р н а я
м а с с а п л о д а). Объекты этого кластера группировались в два устойчивых сгущения (см. рис. 3, А, рис. 4). Стабильную экспрессию признаков регистрировали у сорта Прометей и линии Л 48. Кластеризуемые признаки контролируются двумя аллелями с плейотропным эффектом, который отмечался у четырех (2-й, 3-й, 4-й и 5-й) из пяти изучаемых генотипов. Степень функциональной идентичности с учетом экспрессии в разных условиях произрастания: 1-го и 2-го генотипов — 50,0 %; 1-го и 3-го — 75,0 %; 1-го и 4-го — 75,0 %; 1-го и 5-го — 25,0 %; 2-го и 3-го — 100 %; 2-го и 4-го — 50,0 %; 2-го и 5-го — 50,0 %; 3-го и 4-го — 50,0 %; 3-го и 5-го — 50,0 %; 4-го и 5-го — 0 %.
6-й к л а с т е р (п р о д о л ж и т е л ь н о с т ь II -III ф е н о ф а з) (см. рис. 3, Б, рис. 4). Параметры этого кластера формировали четыре устойчивых сгущения. Стабильная экспрессия одного признака (продолжительность II фенофазы) была характерна для двух генотипов (1-го и 2-го), двух — только для одного (2-го). Эти признаки контролируются четырьмя различными аллелями с плейотропным эффектом, отмеченным у четырех (1-й, 3-й, 4-й и 5-й) из пяти генотипов. Степень функциональной идентичности в разных условиях произрастания: 1-го и 2-го генотипов — 25,0 %; 1-го и 3-го — 75,0 %; 1-го и 4-го — 0 %; 1-го и 5-го — 50,0 %; 2-го и 3-го — 25,0 %; 2-го и 4-го — 25,0 %; 2-го и 5-го — 0 %; 3-го и 4-го — 0 %; 3-го и 5-го — 75,0 %; 4-го и 5-го — 0 %.
7-й к л а с т е р (у в я д а н и е в у с л о в и я х м о н о к у л ь т у р ы —т о л щ и н а с т е н к и п л о д а). Кластер формировал четыре устойчивых сгущения (см. рис. 3, В, рис. 4). Ни у одного из генотипов не отмечалась стабильная экспрессия относящихся к этому кластеру признаков, контролируемых четырьмя аллелями с плейотропным эффектом, который наблюдали у одного (1-го) из пяти генотипов. Степень функциональной идентичности: 1-го и 2-го генотипов — 25,0 %; 1-го и 3-го — 50,0 %; 1-го и 4-го — 75,0 %; 1-го и 5-го — 50,0 %; 2-го и 3-го — 25,0 %; 2-го и 4-го — 50,0 %; 2-го и 5-го — 0 %; 3-го и 4-го — 50,0 %; 3-го и 5-го — 25,0 %; 4-го и 5-го — 25,0 %.
|
Рис. 4. Схемы состава доминантных аллелей кластеров хозяйственно ценных признаков у изученных генотипов овощного перца в зависимости от условий выращивания. Кластеры: длина плода—индекс плода; общий урожай с растения—товарный урожай с растения—число плодов с растения; диаметр плода; высота куста; гниль плодов; средняя масса плода—товарная масса плода; продолжительность II фенофазы—продолжительность III фенофазы; увядание в монокультуре—толщина стенки плода; число камер в плоде—продолжительность I фенофазы; А и Б — соответственно открытый и защищенный грунт; а, б, в, г, д — 1-й, 2-й, 3-й, 4-й и 5-й генотипы (описание см. в разделе «Методика»). |

8-й к л а с т е р (ч и с л о к а м е р в п л о д е—п р о д о л ж и т е л ь-
н о с т ь I ф е н о ф а з ы). Кластер образовал два устойчивых сгущения (см. рис. 3, Г, рис 4). По продолжительности I фенофазы генотипы практически не различались, однако дифференцировались по стабильности проявления этого признака. У двух из пяти генотипов наблюдали стабильную экспрессию по обоим признакам, которые контролируются двумя аллелями с плейотропным эффектом. Такой эффект регистрировали у четырех (2-й, 3-й, 4-й и 5-й) из пяти генотипов. Степень функциональной идентичности генотипов оказалась высокой и составила: 1-го и 2-го — 25,0 %; 1-го и 3-го — 75,0 %; 1-го и 4-го — 75,0 %; 1-го и 5-го — 25,0 %; 2-го и 3-го – 50,0 %; 2-го и 4-го — 50,0 %; 2-го и 5-го — 100 %; 3-го и 4-го — 100 %; 3-го и 5-го — 50,0 %; 4-го и 5-го — 50,0 %.
9-й к л а с т е р (о б щ и й и т о в а р н ы й у р о ж а й с р а с т е н и я —ч и с л о п л о д о в с р а с т е н и я). Этот кластер образовывал шесть устойчивых сгущений (см. рис. 2, Б, рис. 4). Наиболее дифференцированный тип экспрессии в благоприятных условиях (защищенный грунт) наблюдался у сортов Добрыня Никитич, Прометей и линии Л 48 по трем признакам, относительно стабильный — у сорта Колобок и линии Л 49, причем только по величине общего урожая. Изучаемые признаки этого кластера детерминируются шестью аллелями. Плейотропный эффект отмечался у двух (2-й и 3-й) из пяти генотипов у аллелей, контролирующих общий и товарный урожай с растения. С учетом экспрессии в разных условиях произрастания степень функциональной идентичности 1-го и 2-го генотипов равнялась 33,3 %; 1-го и 3-го — 66,7 %; 1-го и 4-го — 50,0 %; 1-го и 5-го — 0 %; 2-го и 3-го — 33,3 %; 2-го и 4-го — 33,3 %; 2-го и 5-го — 16,7 %; 3-го и 4-го — 16,7 %; 3-го и 5-го — 33,3 %; 4-го и 5-го — 0 %.
Итак, пять изученных форм овощного перца CapsicumannuumL. — сорта и линии Добрыня Никитич, Колобок, Л 49, Прометей и Л 48 (соответственно 1-й, 2-й, 3-й, 4-й и 5-й генотипы) дифференцированы по комплексу хозяйственно ценных признаков и степени идентичности доминантных аллелей, контролирующих эти признаки, на высоко- (1-й и 3-й; 3-й и 5-й; 1-й и 4-й), средне- (1-й и 2-й; 2-й и 3-й; 2-й и 4-й; 2-й и 5-й; 3-й и 4-й) и малоидентичные (4-й и 5-й; 1-й и 5-й). Полученные данные позволяют продолжить исследование взаимосвязи между степенью идентичности ключевых аллелей, подбором пар для скрещивания и эффектом гетерозиса. Кроме того, проведенный анализ выявил потенциальных доноров стабильной экспрессии комплекса хозяйственно ценных признаков и плейотропный эффект ряда полигенов, что может быть использовано в селекции при создании принципиально нового исходного материала.
Л и т е р а т у р а
1. Д р а г а в ц е в В.А. К проблеме генетического анализа полигенных количественных признаков растений. СПб, 2003.
2. П о т о к и н а Е.К., Ч е с н о к о в Ю.В. Современные методы геномного анализа в исследованиях генетики количественных признаков. С.-х. биол., 2005, 3: 3-18.
3. D r a g a v t s e v V.A. Algorithms of an ecology-genetical survey of the genofond and methods of creating the varieties of crop plants for yield, resistance and quality. St. Petersburg, 2002.
4. Ч е с н о к о в Ю.М. Идентификация QTL и скрининг генетических ресурсов растений. Мат. I Межд. науч.-практ. конф «Современные тенденции в селекции и семеноводстве овощных культур. Традиции и перспективы». М., 2008: 416-425.
5. P a r a n I., G r u b e R.C., B e n-C h a i m A. e.a. Biometrical and molecular analysis of quantitative traits in pepper (Capsicum annuum). Proc. Xth EUCARPIA Meeting on genetics and breeding of Capsicum & Eggplant. Avignon, France, 1998: 243-244.
6. Генетика количественных признаков сельскохозяйственных растений /Под ред. Д.К. Беляева. М., 1978.
7. М а з е р К., Д ж и н к с Дж. Биометрическая генетика: Пер. с англ. М., 1985.
8. T i m i n a O.O., T s y k a l i u k R.A., O r l o v P.A. The identification of genotypes quantitative characters by regressive cluster analysis. Capsicum and Eggplant Newsletter, 2004, 23: 37-40.
9. Т и м и н а О.О. Идентификация доноров раннеспелости овощного перца Capsicumannuum L. регрессионно-кластерным анализом. Овочiвництво i баштанництво, 2005, 51: 264-281.
10. Ф е д и н М.А., С и л и с Д.Я., С м и р я е в А.В. Статистические методы генетического анализа. М., 1980: 85-111.
11. Т а р а с к и н а А.С., Ч е р е м у ш к и н Е.С. Обработка микрочиповых данных с помощью алгоритма нечеткой кластеризации. Выч. методы и программирование, 2006, 7: 1-5.
IDENTIFICATION OF KEY ALLELES OF ECONOMIC IMPORTANT DETERMINANTS IN Capsicum annuum L. BY THE REGRESSION-CLUSTERED ANALYSIS
O.O. Timina1, A.S. Ryabova2
With the use of parent’s forms of Capsicum annuum (the Dobrynya Nikitich, Kolobok and Prometei varieties, also the L 48 and L 49 lines of the 1th, 3th, 4th and 5th genotype, respectively) and their F1 hybrids, isolated in diallel crossings on [x р(р + 1)] scheme, the authors differentiate the genotypes on high-, middle- and little-identical. The findings permit to determine a correlation between degree of iden-tity of key alleles of economic important determinant by sorting out the pairs for crossing and heterosis effect.
Key words: quantitative traits, regression cluster analysis, vegetable pepper.
1Приднестровский государственный университет, |
Поступила в редакцию |
