








УДК [633.11+635.656]:576.36:612.014.1
АКТИВНЫЕ ФОРМЫ КИСЛОРОДА И АПОПТОЗ У ПШЕНИЦЫ И ГОРОХА
Н.Е. ПАВЛОВСКАЯ, А.И. ГРИНБЛАТ
Исследовали функции ферментов, обезвреживающих активные формы кислорода (супероксиддисмутаза, каталаза и пероксидаза), при апоптозе колеоптилей у пшеницы, а также некрозе 1-го настоящего листа у пшеницы и гороха под влиянием ионов Cd2+ и заражения грибом Fusarium oxysporum. Показано, что в норме и при патогенезе изученные ферменты функционируют как синергисты. При апоптозе и действии ионов Cd2+ каталазная активность возрастает (как в норме, так и при воздействии биотических факторов). Активность супероксиддисмутазы резко падает и при апоптозе, происходящем в клетках колеоптиля пшеницы, и под влиянием ионов Cd2+ в листьях пшеницы и гороха. Следовательно, ионы Cd2+ оказывают апоптозный эффект, а заражение возбудителем фузариозного увядания индуцирует некроз.
Ключевые слова: апоптоз, некроз, активные формы кислорода. супероксиддисмутаза, пероксидаза, каталаза.
Известно, что у живых организмов существуют две формы клеточной гибели: апоптоз, или программируемая смерть, и некроз — патологическая гибель клеток, названная сверхчувствительностью. Сверхчувствительность (некроз) представляет собой одну из форм несовместимой реакции взаимодействия растения с патогеном. Гибель клеток в очаге инфекции можно расценивать как самоубийство части для сохранения целого. В отличие от некроза апоптоз — обязательная функция организма, имеющая большое значение для эмбриогенеза, морфогенеза, дифференцировки тканей, опадения листьев, созревания плодов, а также как инструмент иммунного ответа (1).
У прокариотов апоптоз возник в качестве механизма защиты клеток от активных форм кислорода (АФК). Защита от АФК также относится к функции апоптоза у эукариотических организмов. Повышенное содержание активных форм кислорода отмечается в клетках животных и человека под воздействием радиации, пестицидов, промышленных отходов, при канцерогенезе, инфекционных заболеваниях. А.А. Аверьянов обнаружил аналогичную функцию АФК у растений (2). Предполагается, что свободнорадикальные процессы играют важную роль в механизмах старения и апоптоза. Ферменты, осуществляющие каталитическое обезвреживание или разрушение потенциально вредных для клетки веществ, прежде всего свободных радикалов, хорошо исследованы (3). Это супероксиддисмутаза (СОД), каталаза и пероксидаза. СОД катализирует превращение супероксидного радикала (О2-) в перекись водорода (Н2О2) и молекулярный кислород (О2), каталаза — превращение Н2О2 в Н2О и О2, пероксидаза восстанавливает Н2О2 до воды с использованием различных восстановителей. Поскольку активация кислорода относится к ранним ответам растительной клетки на неблагоприятные воздействия, АФК может принадлежать важная роль в подавлении развития патогенов.
Целью работы было выяснение роли супероксиддисмутазы (СОД), каталазы и пероксидазы — системы ферментов, обезвреживающих активные формы кислорода, в процессах апоптоза колеоптилей пшеницы, а также в реакции сверхчувствительности в 1-м настоящем листе пшеницы и гороха под влиянием ионов Cd2+ и заражения патогенным грибом Fusariumoxysporum.
Методика. Для подготовки проростков семена яровой пшеницы сорта Колумбия и гороха сорта Норд (последние предварительно замачивали в дистиллированной воде на 1 сут) этиолировали на фильтровальной бумаге, смоченной дистиллированной водой, при температуре 25 °С в течение 2 сут в программируемой климатической камере. Проросшие семена каждой культуры отделяли от непроросших и рассаживали в три кюветы с фильтровальной бумагой. В качестве питательной среды в первые (контрольные) кюветы добавляли дистиллированную воду, во вторые — раствор Cd2+ (5·10-3 М); растения из третьих кювет сначала погружали на 2 ч в водную суспензию спор F. oxysporum, затем высаживали, используя водную суспензию спор в качестве питательной среды. Кюветы размещали в программируемой климатической камере и продолжали этиолирование проростков при температуре 25 °С.
C 3-х по 11-е сут от начала проращивания ежесуточно отделяли навески: у пшеницы — колеоптилей контрольных растений, поскольку известно (4), что этот орган пшеницы подвергается запрограммированной клеточной гибели (апоптозу) естественным образом, то есть без введения в питательную смесь каких-либо дополнительных индукторов апоптоза, настоящих листьев контрольных растений, настоящих листьев растений, обработанных Cd2+, настоящих листьев растений, зараженных F. oxysporum; у гороха — листьев контрольных растений, листьев растений, обработанных Cd2+, листьев растений, обработанных F. oxysporum.
В пробах определяли активность группы ферментов (СОД—катала-за—пероксидаза).
Активность СОД оценивали по модифицированной методике (5, 6). Растительный материал измельчали в ступке с 25 мл фосфатного буферного раствора (pH 7,8), центрифугировали (7000 об/мин в течение 15 мин), отделяли надосадочную жидкость (в дальнейшем — экстракт СОД). Анализ проводили методом фиксированного времени. Реакционная среда (общий объем — 2,5 мл) содержала фосфатный буфер (50 мМ, pH 7,8), L-метионин (13 мМ), нитросиний тетразолий (75 мкМ), рибофлавин (2 мкМ), ЭДТА (0,1 мМ) и экстракт СОД (0,5 мл). Рибофлавин добавляли в последнюю очередь и помещали пробирки в фотореактор. Через 15 мин облучение прекращали, выключая лампу фотореактора, затем измеряли оптическую плотность растворов на спектрофотометре Spekol-11 («VЕP Karl Zeis», ГДР) при l = 560 нм. Контролем служила необлученная реакционная смесь, которая не окрашивалась. В отсутствие фермента реакционная смесь приобретала интенсивное пурпурное окрашивание вследствие восстановления нитросинего тетразолия; интенсивность окрашивания смеси, содержащей СОД, была невысока и зависела от активности фермента.
Активность СОД (AСОД) рассчитывали по формуле:
АСОД = |
Dконтр. - Dх |
x |
Vреакц. смеси x Vэкстракта |
, |
Dконтр. |
Vдля реакции x mнавески |
где Dконтр.— среднее арифметическое оптических плотностей контрольных растворов; Dх— среднее арифметическое оптических плотностей растворов с экстрактами СОД; Vреакц. смеси— общий объем реакционной смеси;
Vэкстракта— общий объем экстракта СОД; Vдля реакции— объем экстракта СОД, взятого для реакции; mнавески— масса навески биологического материала.
Для определения активности каталазы использовали модифицированную методику (7), основанную на измерении объема выделившегося О2 после добавления перекиси водорода к водному экстракту каталазы. Растительный материал измельчали в ступке с молотым стеклом в 20 мл фосфатного буфера (рН 7,8). Объем полученной массы доводили до 100 см3 буферным раствором, используя мерную колбу. Затем гомогенат переносили в коническую колбу, которую помещали на механический встряхиватель на 30 мин. Далее суспензию переносили в реакционный сосуд, добавляли 3-5 капель толуола, туда же помещали маленький стаканчик с 3 см3 3 % Н2О2. Сосуд соединяли с остальной частью прибора, используемого для измерения объема выделившегося О2, и запускали реакцию, включая магнитную мешалку (600 об/мин), которая опрокидывала стаканчик. С 3-й по 15-ю мин после начала реакции с интервалом 3 мин регистрировали выделяемый объем О2. Кинетические кривые зависимости этого показателя от времени аппроксимировали функцией вида y = a×ln(x) + b. Исходным показателем активности каталазы служил предлогарифмический коэффициент a, рассчитанный методом дифференциального анализа кинетических кривых, окончательным — приведенный предлогарифмический коэффициент, полученный как отношение коэффициента a к массе навески. Для обработки данных волюмометрического анализа активности каталазы разработали компьютерную программу «Каталазоинформер», позволяющую осуществлять администрирование биохимических данных — групповой ввод, аппроксимацию, спрямление и множественный анализ кинетических кривых (8).
|
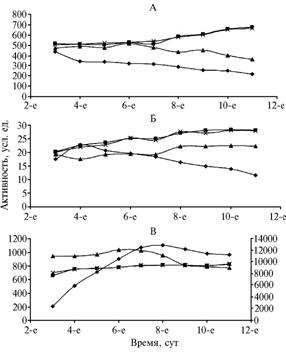
Рис. 1. Активность супероксиддисмутазы (А), каталазы (Б) и пероксидазы (В) в тканях проростков яровой пшеницы сорта Колумбия под влиянием биотических и абиотических факторов: квадраты, ромбы, треугольники и звездочки — соответственно контроль (настоящий лист) и значения показателей в пробах тканей колеоптиля листа проростка, обработанного Cd2+ и Fusarium oxysporum. Значения пероксидазной активности в тканях колеоптиля соответствуют правой ординате. |
Для определения активности пероксидазы использовали методику (7), основанную на окислении бензидина перекисью водорода под действием пероксидазы. Растительный материал измельчали в ступке с молотым стеклом в 20 мл ацетатного буфера (рН 4,8), центрифугировали (4000 об/мин, 15 мин) и далее использовали надосадочную жидкость. С помощью секундомера регистрировали время после добавления бензидина к реакционной смеси, в течение которого ее оптическая плотность при l = 490 нм достигала 0,15. Активность пероксидазы (Ап) рассчитывали по формуле:
АП = |
D·а·б·в |
, |
с·t |
где а — степень разбавления навески в экстракте, мл/г; б — степень разбавления экстракта, взятого для реакции (x40); в — степень разбавления экстракта в реакционной смеси (x80); c — длина оптического пути в кювете (1 см); D — оптическая плотность реакционной смеси (в опыте D = 0,15); t — время, в течение которого эта оптическая плотность достигается, с.
Результаты. В течение 9 сут наблюдений в колеоптилях пшеницы активность СОД резко падала (с 436 усл. ед. у 3-суточных до 211 усл. ед. — у 11-суточных). В тканях листа активность этого фермента, напротив, возрастала (с 516 усл. ед. на 3-и сут до 670 усл. ед. — на 11-е). Под влиянием ионов Cd2+ в листе пшеницы показатель снижался, как и при апоптозном эффекте в колеоптилях. Заражение проростков возбудителем фузариозного увядания F. oxysporum приводило к повышению активности СОД по сравнению с контролем. Активность каталазы листьев в контроле и при заражении F. oxysporum во всех вариантах возрастала (рис. 1), что связано с функцией фермента, обеспечивающего нейтрализацию ядовитых перекисей, которые образуются и при патогенезе, и в норме. Апоптоз в колеоптилях пшеницы приводил к снижению активности каталазы. Пероксидазная система клеток колеоптилей пшеницы (см. рис. 1) при апоптозе интенсивно функционировала до 8-9-х сут, после чего соответствующие показатели начинали снижаться (вплоть до гибели клеток). Подобным образом пероксидаза действовала и в тканях настоящего листа под влиянием ионов Cd2+. При заражении F. oxysporum показатель возрастал по сравнению с контролем в 1-е сут после заражения, со 2-х сут динамика активности фермента в опытных и контрольном образцах была практически одинаковой.
|
Рис. 2. Активность супероксиддисмутазы (А), каталазы (Б) и пероксидазы (В) в тканях проростков гороха сорта Норд под влиянием биотических и абиотических факторов: квадраты, ромбы и треугольники — соответственно контроль и значения показателей в пробах тканей листа проростка, обработанного Cd2+ и Fusarium oxysporum. |
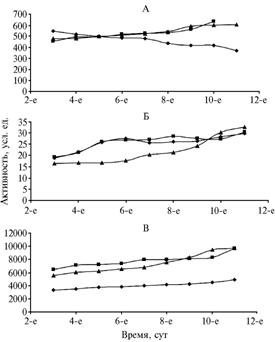
В листьях гороха обработка ионами Cd2+ вызвала апоптозный эффект в отношении СОД (рис. 2); заражение F. oxysporum почти не сказалось на активности фермента. Величина каталазной активности в тканях листьев у гороха (см. рис. 2) под влиянием биотических и абиотических факторов практически ра-внялась контролю, а пероксидазная активность (см. рис. 2) под действием ионов Cd2+ снижалась почти в 2 раза по сра-внению с контролем (соответственно 3271 и 6419 усл. ед.).
Таким образом, в норме и при патогенезе ферменты супероксиддисмутаза (СОД), каталаза и пероксидаза, защищающие аэробную клетку от «самозагрязнения» О2- и Н2О2 (2), функционируют как синергисты. При апоптозе и действии ионов Cd2+ активность каталазы снижается, а активность пероксидазы возрастает как в норме, так и под влиянием биотических факторов. Активность СОД резко снижается при апоптозе (в клетках колеоптиля пшеницы), а также в присутствии ионов Cd2+ (в тканях листьев пшеницы и гороха). Следовательно, ионы Cd2+ оказывают апоптозный эффект на клетки настоящих листьев пшеницы и гороха. Заражение растений возбудителем фузариозного увядания подобных проявлений не вызывает, однако отмечается реакция сверхчувствительности (некроз).
Л И Т Е Р А Т У Р А
1. Д ь я к о в Ю.М., О з е р е ц к о в с к а я О.Л., Д ж а в а х и я В.Г. и др. Общая и молекулярная фитопатология. Уч. пос. М., 2001.
2. А в е р ь я н о в А.А. Активные формы кислорода и иммунитет растений. Усп. совр. биол., 1991, III: 722-737.
3. Х о ч а ч к а П., С о м е р о Дж. Стратегия биохимической адаптации. М., 1977.
4. B a k e e v a L.E., Z a m y a t i n a V.A., S h o r i n g B.Yu. e.a. Apoptosis in the initial leaf of etiolated wheat seedlings: influence of the antioxidant ionol (BHT) and peroxides. Biochemistry, 2001, 66(8): 850-859.
5. G i a n n o p o l i t i e s C.N., R i e s S.K. Superoxid dismutase. I. Occurrence in higher. Plant Physiol., 1977, 59: 309-314.
6. Г р и н б л а т А.И. Способ определения активности фермента. Патент на изобретение № 2293969Б. Опубл. Бюл. изобретений, № 5 от 20.02.2007. М., 2007.
7. Методы биохимического исследования растений /Под ред. Е.А. Ермакова Л., 1987: 38-112.
8. Г р и н б л а т А.И. Разработка биоинформационной модели апоптоза и некроза у растений. Автореф. канд. дис. Орел, 2007.
ACTIVE FORMS OF OXYGEN AND APOPTOSIS IN WHEAT AND PEA
N.E. Pavlovskaya, A.I. Grinblat
The authors investigated the functions of enzymes neutralizing the active forms of oxygen (superoxide dismutase, catalase and peroxydase) during of coleoptiles apoptosis in wheat, necrosis of first true leaf in wheat and pea under the influence of Cd2+ ions and infection by Fusarium oxysporum. It was shown, that in normal and pathological conditions the studied enzymes act as synergists. During apoptosis and Cd2+ ions action the catalase activity increases (in normal and under the influence of biotic factors). The superoxide dismutase activity drops sharply during apoptosis in cells of wheat coleoptiles and under the influence of Cd2+ ions in leaves of wheat and pea. Consequently, the Cd2+ ions have an effect of apoptosis and the infection by fusariose pathogen induces the necrosis.
Key words: apoptosis, necrosis, active oxygen, superoxide dismutase, peroxidation, catalase.
ФГОУ ВПО Орловский государственный |
Поступила в редакцию |




