








УДК 633.111.1:58.02:581.14:581.111
ОСОБЕННОСТИ РОСТА И ПОТРЕБЛЕНИЯ ВОДЫ РАСТЕНИЯМИ СОРТОВ ЯРОВОЙ МЯГКОЙ ПШЕНИЦЫ С КОНТРАСТНОЙ ОТЗЫВЧИВОСТЬЮ НА ЛОКАЛЬНОЕ ВНЕСЕНИЕ УДОБРЕНИЙ
И.И. ИВАНОВ, В.К. ТРАПЕЗНИКОВ, Н.Г. ТАЛЬВИНСКАЯ
В модельных экспериментах (водная культура) изучали влияние локального высоко-солевого воздействия на ростовую функцию и потребление воды растениями двух сортов яровой мягкой пшеницы с контрастной отзывчивостью на гетерогенное распределение удобрения в почве. Показано, что генотипические различия в ответных реакциях на такое воздействие достаточно четко проявляются на самых ранних этапах онтогенеза растений.
Ключевые слова: яровая пшеница, сорт, рост, потребление воды, гетерогенность питательной среды.
Показано, что технология локального внесения минеральных удобрений по сравнению с разбросным способом позволяет повысить (на 10-20 % и более) урожай культурных растений без снижения (а чаще с повышением) биохимических и технологических показателей его качества. При локальном внесении ту же прибавку урожая можно получить при дозах удобрений, которые на 30-50 % ниже. Не менее важно, что локализация минеральных удобрений повышает устойчивость растений к засухе. Столь значимые эффекты достигаются при фактическом равенстве ресурсов внешней среды (запасы элементов минерального питания, воды, энергия солнечного излучения) и генетического потенциала растений. При неравномерном распределении удобрения создается зона с экстремально высоким (на порядок и более) содержанием элементов минерального питания, поэтому с начальных этапов онтогенеза часть корневой системы растения или даже часть отдельно взятого корня функционирует в условиях солевого стресса.
В то же время вопрос об отзывчивости сортов культурных растений на подобное воздействие остается малоизученным. Работы, выполненные на ограниченном наборе генотипов ячменя, яровой мягкой и твердой пшеницы, сои, шпината и редиса (1-4), не отражают, как нам представляется, всего разнообразия ответных реакций растений на особенности минерального питания, создаваемые гетерогенным распределением удобрения в почве. В исследованиях на 15 сортах яровой мягкой и твердой пшеницы различного эколого-географического происхождения выявлен широкий спектр изменчивости показателей ростовой функции, структуры урожая, потребления элементов питания и продуктивности растений в ответ на локальное внесение удобрений. Так, урожай зерна по сравнению с разбросным способом колебался от -5 до +25 % (5). Причины такой неодинаковой генотипической отзывчивости неясны. Менее всего исследован ответ растений на локальное высокосолевое воздействие на начальных этапах онтогенеза.
Наиболее выраженной реакцией на гетерогенность корнеобитаемой среды, отмечавшейся многими исследователями, служит активация ветвления корней в зоне повышенного содержания элементов минерального питания (2, 5-7). Однако наблюдения за ростом корней, особенно при выращивании растений в почве, сопряжены с методическими трудностями, поэтому целесообразными представляются модельные эксперименты в условиях водной культуры.
В задачу нашей работы входило изучение реакции растений двух сортов яровой мягкой пшеницы на гомогенное и гетерогенное распределение элементов минерального питания в среде (гетерогенность питательной среды рассматривали как аналог локального применения минеральных удобрений в полевых условиях).
Методика. Объектами исследования были два сорта яровой мягкой пшеницы с контрастной отзывчивостью на локальное внесение комплексного минерального удобрения (NPK) — Воронежская 12 и Саратовская 62. На выщелоченном черноземе лесостепной зоны Республики Башкортостан при локальном внесении удобрений по сравнению с разбросным способом урожай зерна у первого сорта возрастал на 17 %, у второго — не изменялся (8).
Семена проращивали на плотиках на дистиллированной воде в течение 5 сут. За 1 сут до постановки эксперимента воду заменяли питательным раствором Хогланда-Арнона 1 (9) (5 % от нормы) с микроэлементами. В фазу выхода из колеоптиля 1-го листа проростки переводили на питательную среду с различной концентрацией элементов питания. На гомогенной высокосолевой (ВС — 300 % содержание солей) и низкосолевой (НС — 5 %) средах размещали все пять зародышевых корней. В первом варианте создавали условия солевого стресса, во втором — дефицита элементов минерального питания для всей корневой системы. На гетерогенной среде (300 и 5 %) — корни распределяли в сочетании ВС:НС = 2:3, которое наиболее благоприятно при выращивании растений пшеницы в условиях изолированного питания. Растения находились в градуированных пробирках, вмещающих 60 мл питательного раствора, на светоплощадке при 14-часовом дне и освещенности 16 000 лк. В дневные часы температура воздуха составляла 22-24 °С, в ночные — 18-20 °С. В течение всех экспериментов pH питательных растворов поддерживали в пределах 6,5-7,2 (pH исходных растворов — 4,2-4,4). Потерю объема компенсировали ежесуточным добавлением дистиллированной воды. Смену питательного раствора не проводили. Ростовую функцию оценивали по изменению сухой массы побега, корней и целого растения. Биологическая повторность опыта — 6-кратная, представлены средние значения из двух опытов. Продолжительность выращивания растений — 14 сут. Экспериментальные данные обрабатывали статистически с определением стандартной ошибки.
Результаты. Известно, что рост — интегральный критерий реакции растений на условия произрастания. В наших опытах качественная реакция растений при испытанных режимах минерального питания (масса побега, корней и целого растения) имела сходный характер, а показатели ранжировались по одинаковой схеме: гомогенная высокосолевая среда < гомогенная низкосолевая среда < гетерогенная среда (табл. 1). Однако у растений сорта Воронежская 12 различия, как правило, были более контрастными, чем у неотзывчивого сорта Саратовская 62: у первого при дефиците элементов в питательном растворе и на гетерогенной среде значения сухой массы корней были выше, чем при солевом стрессе, соответственно на 39 и 56 %, у второго — только на 8 и 27 %. Характерно, что по абсолютным значениям сухой биомассы побега, корней и целых растений на гомогенной высокосолевой и низкосолевой средах достоверных сортовых различий мы не выявили, однако они оказались существенными на гетерогенной среде: у растений сорта Воронежская 12 масса побега, корней и целого растения была выше, чем у растений сорта Саратовская 62, соответственно на 37, 14 и 32 %. Кроме того, у сортов различались значения соотношения между массой побега и корней: у первого они изменялись в более широких пределах (2,9-3,6), чем у второго (3,0-3,4). Все это представляется следствием неодинаковых генетически обусловленных возможностей растений изученных сортов адаптироваться к высокосолевому воздействию различной напряженности (в частности, за счет перераспределения ассимилятов между фотосинтезирующим побегом и корнями).
| 1. Сухая масса (X±x) частей и целых растений пшеницы разных сортов в зависимости от концентрации и распределения элементов питания в среде | |||||||
Питательная среда |
Побег |
Корневая система |
Целое растение |
Масса побега/масса корней |
|||
мг |
% |
мг |
% |
мг |
% |
||
В о р о н е ж с к а я 12 |
|||||||
Гомогенная ВС |
46,7±5,7 |
100 |
12,8±1,5 |
100 |
59,5±4,2 |
100 |
3,6 |
Гомогенная НС |
51,9±2,0 |
111 |
17,8±0,8 |
139 |
69,7±2,7 |
117 |
2,9 |
Гетерогенная |
72,7±5,2 |
156 |
20,0±1,1 |
156 |
92,7±6,2 |
156 |
3,6 |
С а р а т о в с к а я 62 |
|||||||
Гомогенная ВС |
43,0±2,5 |
100 |
13,7±0,8 |
100 |
56,7±2,9 |
100 |
3,1 |
Гомогенная НС |
50,5±2,7 |
117 |
14,8±1,0 |
108 |
65,3±3,6 |
115 |
3,4 |
Гетерогенная |
52,9±4,9 |
123 |
17,5±1,3 |
127 |
70,4±6,2 |
124 |
3,0 |
П р и м е ч а н и е. ВС и НС — соответственно высоко- и низкосолевая среда. |
|||||||
Известно, что линейные размеры корневой системы играют существенную роль в поиске и обеспечении растения элементами минерального питания и водой. Естественно, степень проявления этого признака у растений сортов с неодинаковой отзывчивостью на локальное внесение удобрений представляет большой интерес. По нашим данным, суммарная длина зародышевых корней у обоих сортов возрастала в ряду гомогенная высокосолевая среда < гомогенная низкосолевая среда < гетерогенная среда (рис. 1). Наиболее значимые различия между сортами проявились при
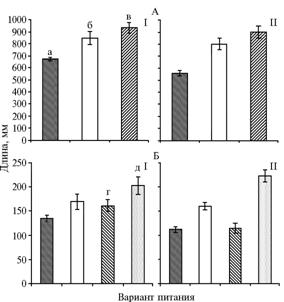 |
Рис. 1. Суммарная длина зародышевых корней (А) и средняя длина корня (Б) у растений пшеницы сортов Воронежская 12 (I) и Саратовская 62 (II) с контрастной отзывчивостью на локальное внесение удобрений: а, б — соответственно высокосолевая и низкосолевая среда при гомогенном питании, в — гетерогенное питание; г, д — соответственно высокосолевая и низкосолевая среда при гетерогенном питании. |
выращивании на гомогенной высокосолевой среде: у растений сорта Воронежская 12 длина зародышевых корней была на 20 % больше, чем у растений сорта Саратовская 62. Различия по суммарной длине корней между сортами в двух других вариантах были менее выраженными, но с некоторым преимуществом у отзывчивого сорта.
Поскольку на гетерогенной питательной среде ко-рни распределяли по схеме ВС:НС = 2:3, представлял интерес вопрос о том, как фактор гетерогенности корнеобитаемой среды отражается на средней длине одного корня. Гетерогенность питательной среды активировала линейный рост корней низкосолевого статуса (относительно соответствующего показателя при дефиците элементов питания) у растений обоих сортов, а также снижала ингибирующий эффект высокой концентрации ионов в среде у растений сорта Воронежская 12. У растений сорта Саратовская 62 подобного эффекта не наблюдали: средняя длина высокосолевого корня оставалась такой же, как у растений, произраставших на гомогенной высокосолевой среде. В целом, при локальном и тотальном высокосолевом воздействии средняя длина одного корня высокосолевого статуса у растений сорта Саратовская 62 была достоверно меньше, чем у растений сорта Воронежская 12.
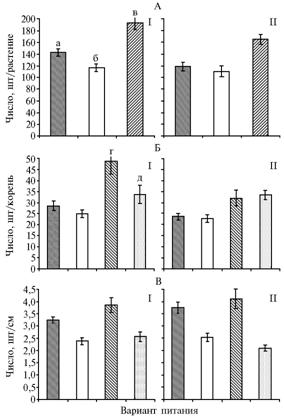 |
Рис. 2. Показатели, характеризующие ветвление корней при различной напряженности солевого воздействия у растений пшеницы сортов Воронежская 12 (I) и Саратовская 62 (II): А — среднее число боковых корней на растении, Б — среднее число боковых корней на один зародышевый корень, В — плотность распределения боковых корней в зоне ветвления; а, б — соответственно высокосолевая и низкосолевая среда при гомогенном питании, в — гетерогенное питание; г, д — соответственно высокосолевая и низкосолевая среда при гетерогенном питании. |
Вероятно, наиболее наглядное проявление эффекта локального внесения азотных и фосфорных минеральных удобрений — активация ветвления корней в соответствующей почвенной зоне. Генотипические особенности этой ответной реакции растений остаются неизученными. В наших экспериментах, как и следовало ожидать, наименее выраженное ветвление отмечали для обоих сортов у растений, все корни которых находились в низкосолевой среде (рис. 2), при практически одинаковом числе боковых корней. Иное наблюдали при высокосолевом воздействии на всю корневую систему или ее часть. В последнем варианте у отзывчивого сорта Воронежская 12 сформировалось 192 боковых корня, у неотзывчивого Саратовская 62 — 165. Подобные различия, особенно на ранних этапах онтогенеза, могут играть важную роль в поиске и поглощении элементов минерального питания и воды. При действии повышенной концентрации солей на всю корневую систему у растений первого сорта эффект ветвления зародышевых корней проявлялся достоверно сильнее, чем у второго, то есть имела место та же закономерность, что и по суммарной длине корней. Однако общее число сформировавшихся боковых корней в расчете на растение не дает представления о влиянии локального высокосолевого воздействия на процесс ветвления материнских корней различного солевого статуса. Поэтому мы сравнили число боковых корней в расчете на один зародышевый корень на гетерогенной и гомогенной питательных средах. Оказалось, что у растений обоих сортов на гетерогенной питательной среде проявлялась четко выраженная стимуляция ветвления как высокосолевых, так и низкосолевых корней (см. рис. 2). Например, у растений сорта Воронежская 12 на один высокосолевой и низкосолевой корень при гетерогенном питании приходилось соответственно в 1,75 и 1,36 раза больше боковых корней, чем на корень аналогичного солевого статуса на гомогенных средах. Следовательно, на гетерогенной питательной среде активируется ветвления корней как непосредственно подвергающихся высокосолевому воздействию, так и функционирующих в низкосолевой среде. Механизм передачи сигнала от высокосолевой пряди к низкосолевой и его природа остаются неясными. Возможно, он имеет электрическую, гормональную или трофическую природу (не исключается и их совокупное влияние).
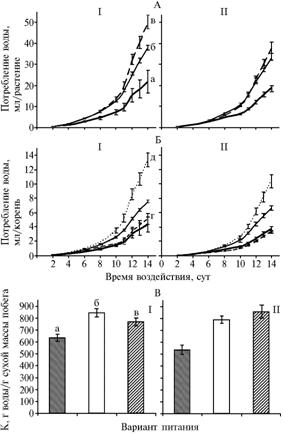 |
Рис. 3. Динамика потребления воды (А — в расчете на растение, Б — на корень) и коэффициент транспирации К (В) при различной напряженности солевого воздействия у растений пшеницы сортов Воронежская 12 (I) и Саратовская 62 (II): а, б — соответственно высокосолевая и низкосолевая среда при гомогенном питании, в — гетерогенное питание; г, д — соответственно высокосолевая и низкосолевая среда при гетерогенном питании. |
В настоящей работе не изучали влияние напряженности высокосолевого воздействия и характера распределения солей в среде на структуру корня и его митотическую активность, однако представляется, что оно весьма существенно как на клеточном, так и на молекулярном уровне. Основанием для такого предположения служат полученные данные по абсолютной и относительной величине базальной зоны без боковых корней, зоны ветвления и апикальной части корня, включающей в себя зоны растяжения клеток и меристему. Реакция растений изученных сортов по перечисленным показателям была практически одинаковой (табл. 2). В обоих вариантах на апикальную часть корня без боковых корней на гомогенной высокосолевой среде приходилась 1/5 общей длины, на низкосолевой — почти 1/3 составляла апикальная зона; на зону ветвления приходилось около 60 %. При гетерогенном питании у растений обоих сортов заметно изменялось соотношение размера зон зародышевых корней: у корней как высоко-, так и низкосолевого статуса повышалась доля зоны ветвления и снижалась относительная величина апикальной части без боковых корней по сравнению с этими показателями на гомогенной среде. Вероятно, подобная реакция связана с иным функциональным состоянием материнских корней, затрагивающим процесс закладки примордиев и их развитие в боковые корни. Во многих исследованиях показана роль фитогормонов (ауксинов и цитокининов) в регуляции ветвления корней, однако также высказывается предположение, что этот процесс имеет более сложную природу и регулируется на уровне целого растения (10). Регуляция ветвления зародышевых корней на гетерогенной питательной среде, по-видимому, происходит по еще более сложной схеме, чем на гомогенной, поскольку локальное высокосолевое воздействие на зародышевые корни активирует не только линейный рост корней низкосолевого статуса, но и их ветвление, что наиболее ярко проявлялось у растений отзывчивого сорта Воронежская 12.
| 2. Относительная длина зон корней различного солевого статуса (от общей длины корня, %) у растений пшеницы разных сортов в зависимости от концентрации и распределения элементов питания в среде | ||||||
Питательная среда |
Воронежская 12 |
Саратовская 62 |
||||
1 |
2 |
3 |
1 |
2 |
3 |
|
Гомогенная ВС |
15 |
65 |
20 |
19 |
60 |
21 |
Гомогенная НС |
16 |
53 |
31 |
10 |
60 |
30 |
Гетерогенная: |
|
|
|
|
|
|
ВС корни |
13 |
75 |
12 |
15 |
74 |
11 |
НС корни |
10 |
67 |
23 |
10 |
74 |
16 |
П р и м е ч а н и е. То же, что в таблице 1; 1 — базальная часть корня без боковых корней; 2 — зона ветвления; 3 — апикальная часть корня без боковых корней. |
||||||
Физиология продукционного процесса в условиях гетерогенного распределения элементов минерального питания в почве остается малоизученной, что в полной мере относится и к проблеме водного обмена растений. Воспроизвести в условиях водной культуры все возможное разнообразие солевого воздействия на корневую систему, возникающее при локальном внесении удобрения в почву, невозможно. В почве даже по длине корня солевой статус его частей может существенно различаться и изменяться от стрессовых до фоновых значений. Несмотря на это, как показали наблюдения, в условиях водной культуры все же удается выявить определенные различия в потреблении воды у растений сортов с контрастной отзывчивостью на локальное внесение минеральных удобрений. Состав питательных сред и характер распределения в них элементов питания в целом оказывали сходный эффект на потребление воды растениями обоих сортов (рис. 3). Общее количество поглощенной воды за период эксперимента ранжировалось в ряду гетерогенная среда > дефицит элементов питания > солевой стресс. Высокосолевое воздействие на все зародышевые корни приводило к явному ингибированию их водонагнетающей активности. Последнее, вероятно, связано с подавлением осмотической составляющей корневого давления (11) и величиной потерь воды на испарение через открытие устьиц. Суммарное количество поглощенной растениями воды определяется не только эндогенными механизмами регуляции ее поглощения и транспорта, но и массой побега. Прирост сухой массы побега за период эксперимента у растений сорта Воронежская 12 при всех режимах питания (и особенно на гетерогенной среде) был выше, чем у растений сорта Саратовская 62, поэтому растения отзывчивого сорта Воронежская 12 потребляли за весь период эксперимента большее количество воды, чем неотзывчивого Саратовская 62.
Учет суммарного потребления воды целым растением не позволяет выявить роль гетерогенитета питательной среды в изменении этой функции у корней различного солевого статуса, поскольку высокосолевому воздействию (в отличие от общего солевого стресса) подвергается лишь часть корневой системы. Этот вопрос решается сравнением поглощения воды одним корнем. Из наших данных следует, что локальное высокосолевое воздействие в большей степени влияло на поглощающую активность низкосолевых корней, чем высокосолевых. У растений сорта Воронежская 12 один низкосолевой корень на гетерогенной среде поглотил в 1,76 раза больше воды, чем на гомогенной, у растений сорта Саратовская 62 — в 1,55 раза больше (см. рис. 3). Неоднозначное влияние гетерогенной питательной среды проявилось в отношении поглощения воды корнями высокосолевого статуса: если у растений сорта Воронежская 12 один высокосолевой корень поглощал на 18 % больше воды, чем при воздействии солей на всю корневую систему, то у растений сорта Саратовская 62 этот показатель был ниже на 10 %.
Таким образом, гетерогенность корнеобитаемой среды вызывает четко выраженную специализацию корней различного солевого статуса: основной вклад в обеспечение побега водой вносят низкосолевые корни. Сходные результаты были получены нами ранее по транспорту ионов, аминокислот и фитогормонов с ксилемным экссудатом (2, 3). Условия питания влияли и на такой показатель водного обмена растений, как коэффициент транспирации, то есть расход количества воды на синтез единицы сухой массы: в условиях высокосолевого воздействия на всю корневую систему он был наименьшим при наибольшем коэффициенте эффективности использования воды на синтез биомассы побега, чем при двух других режимах питания (см. рис. 3). Реакция растений обоих сортов на дефицит элементов питания и гетерогенитет питательной среды была сходной.
Для выявления генотипических особенностей реакции растений на гетерогенность корнеобитаемой среды представляется важным сравнение функциональной активности пряди корней, непосредственно подвергающейся высокосолевому воздействию. В наших экспериментах в высокосолевую среду помещали два корня (или 40 % от общего числа). Как оказалось, их вклад в ростовые процессы и поглощение воды целым растением не всегда адекватен числу (табл. 3), причем между отзывчивым и неотзывчивым сортом проявились достаточно четкие различия: у первого вклад высокосолевых корней был выше, чем у второго (в наибольшей степени — по суммарной длине зародышевых корней и числу образовавшихся боковых) (см. табл. 3). Вероятно, испытуемые сорта характеризуются неодинаковыми адаптивным потенциалом. На гетерогенной среде высокосолевое воздействие ингибировало поглощение воды высокосолевыми корнями у растений обоих сортов практически в одинаковой степени. На их долю приходилась лишь 1/5 часть воды, поставляемой в побег, то есть столько, сколько в среднем на один корень низкосолевого статуса.
| 3. Вклад высокосолевой пряди корней в формирование корневой системы и поглощение воды (от суммарных значений для целого растения, %) на гетерогенной питательной среде у разных сортов пшеницы | ||||
Сорт |
Сухая масса корней |
Длина |
Число боковых корней |
Поглощение |
Воронежская 12 |
47 |
35 |
50 |
20 |
Саратовская 62 |
40 |
26 |
39 |
18 |
Итак, гетерогенное распределение элементов минерального питания в условиях водной культуры позволяет выявить ряд различий в ответных реакциях растений у сортов яровой мягкой пшеницы с контрастной отзывчивостью на локальное внесение удобрений. Поскольку эти различия проявляются на начальных этапах онтогенеза, с их помощью можно с минимальными затратами диагностировать отзывчивость сортов на локальное внесение удобрений, что особенно важно при подборе родительских пар для скрещивания, а также отборе перспективных форм из гибридных популяций при целенаправленной селекции сортов, адаптированных к локальному высокосолевому воздействию на корневую систему.
Л И Т Е Р А Т У Р А
1. К а л и к и н с к и й А.А. Пути повышения эффективности применения минеральных удобрений под зерновые культуры (на примере Белорусской ССР). Автореф. докт. дис. Рига, 1977.
2. Т р а п е з н и к о в В.К. Физиологические основы локального применения минеральных удобрений. М., 1983.
3. Т р а п е з н и к о в В.К., И в а н о в И.И., Т а л ь в и н с к а я Н.Г. Локальное питание растений. Уфа, 1999.
4. С о к о л о в О.А., С е м е н о в В.М. Теория и практика рационального применения азотных удобрений. М., 1992.
5. D r e w M.C. Comparison of the effects of a localized supply of phosphate, nitrate, ammonium and potassium on the growth of the seminal root system, and the shoot in barley. New Phytologist, 1975, 75: 479-490.
6. H o d g e A. The plastic plant: root responses to heterogeneous supplies of nutrients. New Phytologist, 2004, 162: 9-24.
7. T r a p e z n i k o v V.K., I v a n o v I.I., K u d o y a r o v a G.R. Effect of heterogeneous distribution of nutrients on root growth, ABA content and drought resistance of wheat plants. Plant and Soil, 2003, 252(2): 207-214.
8. Т р а п е з н и к о в В.К., И в а н о в И.И., Т а л ь в и н с к а я Н.Г. и др. Особенности минерального питания сортов и видов яровой пшеницы при различных способах внесения сложного удобрения. Агрохимия, 2004, 1: 51-59.
9. Г р о д з и н с к и й А.М., Г р о д з и н с к и й Д.М. Краткий справочник по физиологии растений. Киев, 1973.
10. П л о щ и н с к а я М.Е., И в а н о в В.Б., С а л м и н С.А. и др. Анализ возможных механизмов регуляции ветвления корня. Журн. общ. биол., 2002, 63(1): 68-74.
11. Ж о л к е в и ч В.Н. Транспорт воды в растении и его эндогенная регуляция. 66-е Тимирязевское чтение. М., 2001.
FEATURES OF THE GROWTH AND THE WATER CONSUMPTION BY PLANTS OF SPRING SOFT WHEAT WITH CONTRASTING TENDERNESS ON LOCAL APPLICATION OF FERTILIZERS
I.I. Ivanov, V.K. Trapeznikov, N.G. Tal’vinskaya
In model experiments (water culture) the authors studied the local highly saline effect on a growth and water consumption by two varieties of spring soft wheat with contrasting tenderness on heterogenic distribution of fertilizers in soil. It was shown, that genotypic differences in the responses on such exposure become apparent early in the development of plants.
Key words: spring wheat, variety, growth, water using, nutrient heterogeneity.
Институт биологии Уфимского научного центра РАН, |
Поступила в редакцию |




