








УДК 633.88:581.144:615.32
НАКОПЛЕНИЕ И ИЗМЕНЧИВОСТЬ СОДЕРЖАНИЯ ЭКДИСТЕРОИДОВ В ЛЕКАРСТВЕННОМ СЫРЬЕ ЛЕВЗЕИ САФЛОРОВИДНОЙ
Н.П. ТИМОФЕЕВ
В условиях европейского Севера в течение 17-летнего жизненного цикла агропопуляции изучали продуктивное долголетие в связи с накоплением действующих веществ и изменением качества лекарственного сырья левзеи сафлоровидной (Rhaponticum carthamoides). Установлено, что основой потенциала продуктивного долголетия вида служит система вегетативных полициклических побегов. Накопление экдистероидов в онтогенезе положительно коррелирует с величиной продуктивности надземных органов, минимальные значения во время вегетации совпадают с наступлением засушливых условий. Динамика содержания экдистероидов в отдельных элементах и фракциях фитомассы согласуется с процессом формирования побегов, при котором происходит перераспределение биологически активных веществ от старых в пользу растущих и развивающихся органов.
Ключевые слова: левзея сафлоровидная, развитие в онтогенезе, продуктивность, качество лекарственного сырья, содержание экдистероидов, 20-гидроксиэкдизон.
Rhaponticum carthamoides (Willd.) Iljin (синонимы: Leuzea carthamoides DC., левзея сафлоровидная, рапонтикум, большеголовник альпийский, маралий корень) — многолетний, быстрорастущий вид сем. Asteraceae (Compositae). R. carthamoides и препараты на его основе включены в Государственную фармакопею СССР, Российской Федерации, а также в Государственный реестр лекарственных средств России (1, 2). Вид интродуцирован из высокогорной зоны субальпийского пояса (1200-2700 м над уровнем моря); характеризуется длительностью жизненного цикла, нетребователен к почвенным условиям (3), относится к реликтам плейстоценового флористического комплекса, обладающим специфической формой вторичного метаболизма, который сопровождается биосинтезом фитоэкдистероидов, обусловливающих биологическую активность (4, 5).
Согласно санитарно-эпидемиологическим правилам и нормативам Российской Федерации (СанПиН 2.3.2.2351-08), с 1 мая 2008 года разрешено использовать все части R. carthamoides (корни, корневища, стебли, листья и соцветия), их экстракты и продукты переработки в составе фармпрепаратов, одно- и многокомпонентных биологически активных добавок к пище (без ограничений, установленных для других 16 видов растительных адаптогенов). Из действующих веществ в лекарственном сырье содержание мажорного (основного на 95-98 % по массовой доле) экдистероида 20-гидроксиэкдизона (20Е) должно быть не менее 0,1 % в расчете на сухое вещество (6).
Недостатки использования корневищ — невозобновляемость источника сырья после заготовки, сложность и трудоемкость операций по уборке, очистке, сушке и хранению. Кроме того, подземные органы характеризуются низким содержанием действующих веществ по сравнению с листовой частью (7) и их большими потерями при хранении и переработке (8). В листьях может накапливаться до 40 % экстрактивных веществ по сравнению с 13 % в подземных органах (6).
Проблема технологии производства растительного сырья из надземной фитомассы экдистероидсодержащих видов заключается в неравномерности распределения действующих веществ по органам (9, 10), поскольку синтез органического вещества и накопление вторичных метаболитов в лекарственных растениях весьма сложно скоррелированы с развитием в онтогенезе и ростом в течение вегетационного периода (11). Целью работы было изучение морфофизиологических и биохимических характеристик различных элементов в структуре фитомассы лекарственного сырья и их фракций.
Методика.Исследования выполняли на юго-востоке Архангельской области, в подзоне средней тайги (г. Котлас-Коряжма; географические координаты — 61°20'' с.ш., 47° в.д.) в 1989-2006 годах на двух разновозрастных (12 и 17 лет) агропопуляциях R. carthamoides (площадь соответственно 1 и 3 га,единый земельный массив). Агропопуляции представлены моноэдификаторными ценозами семенного происхождения. Первая заложена в 1989 году семенами, полученными из Коми НЦ УрО РАН, вторая — в 1994 году семенами собственного производства. Густота всходов — 96-121, плотность с 3-го по 10-й год — 27-23, с 13-го по 17-й — 16-14 тыс. экз/га.
Предшественниками в севообороте были картофель, однолетние и зерновые культуры. Предпосевная обработка почвы включала вспашку на глубину 22-25 см, дискование и 2-кратную культивацию с одновременным боронованием. Перед посевом участок прикатывали. Посев подзимний с междурядьями 70 см, в середине октября, четырехрядной овощной сеялкой СОН-2.8 (Россия). Норма высева семян — 2,2-2,7 кг/га при полевой всхожести 58-66 %, глубина заделки — 2-3 см. Минеральные удобрения (NPK60-90) вносили только в первые три года после посева; далее удобрения, химические средства защиты и регуляторы роста растений не применяли. Для отбора модельных растений ежегодно в начале фазы отрастания закладывали учетные площадки. Делянки (60-80 м2 каждая) размещали по двум диагоналям поля в 5-6-кратной повторности по трансектам. Изъятие образцов надземной биомассы проводили в фазы бутонизации и цветения (I и III декада июня), массовое отчуждение (силосная масса) — начиная с 4-го года однократно после фазы плодоношения (III декада июля—I декада августа), выкопку корневищ — осенью, после завершения вегетации (октябрь).
Рельеф местности слабоволнистый повышенный, почвы супесчаные, дерново-среднеподзолистые, сформированы на двучленных отложениях. Верхний горизонт (0-28 см) — частицы песка, с глубины 70-85 см преобладает среднесуглинистая фракция. По комплексу агрохимических показателей почва участка относится к высокоокультуренным минеральным (почвенные пробы исследовали общепринятыми методами в ФГУ «Агрохимцентр Кировский», г. Киров). Содержание гумуса в пахотном слое 3,6; органического вещества — 3,1 %. Кислотность корнеобитаемого слоя оптимальная (pHKCl обменная 6,4-6,5; pH гидролитическая 0,7 мг-экв); насыщенность основаниями высокая (12,4 мг-экв, или 93,5 %). По элементам питания обеспеченность фосфором высокая (P2O5 подвижный, 31,2 мг/100 г почвы), калием средняя (K2O подвижный, 9,6 мг/100 г); содержание Ca — 6,4, Mg — 1,0 мг-экв/100 г.
Особенности климата, средняя температура воздуха и почвы описаны в литературе (12, 13). Продолжительность светлого времени суток в начале отрастания R. carthamoides (I декада мая) — 16-17 ч, во время цветения (во II-III декаде июня) — 20 ч. Влажность почвы определяли в корнеобитаемом слое популяций в течение четырех вегетационных периодов (2000-2003 годы). Образцы отбирали по диагонали исследуемого участка в 6-10 точках на глубину пахотного слоя 22-25 см, формировали среднюю пробу, в течение 7-10 сут доводили до воздушно-сухого состояния при комнатной температуре, затем досушивали при 130 °С в течение 30 мин для удаления гигроскопической влаги.
Изучение онтогенезарастений и его периодизацию проводили по Т.А. Работнову (14) и Л.Б. Заугольновой с соавт. (15). Возрастные состояния учитывали по доминирующей группе особей. Выделяли виргинильный (прегенеративный), генеративный и сенильный (постгенеративный) периоды онтогенеза. В виргинильный период выделяли следующие возрастные состояния: проростки (p), ювенильные (j), имматурные (im) и взрослые вегетативные растения (v), в генеративном — молодые (g1), средневозрастные (g2) и старые генеративные растения (g3), в сенильном — субсенильное возрастное состояние (ss). Календарный (абсолютный) возраст популяций отсчитывался со времени появления всходов.
В прегенеративный период индикаторными признаками возрастных состояний служили: для ювенильного — формирование зародышевого розеточного побега со стеблекорнем, для имматурного — кущение вегетативных побегов и формирование корневища, для виргинильного — развитие системы вегетативных побегов, отсутствие плодоношения. Критерии молодого генеративного состояния: формирование репродуктивных побегов, слабое плодоношение, отсутствие признаков отмирания корневища; взрослого — относительный максимум числа репродуктивных побегов, высокая интенсивность роста и плодоношения, уравновешенность процессов новообразования и отмирания; старого — резкое снижение доли репродуктивных побегов, ослабленный рост, неполноценность и периодичность плодоношения, преобладание процессов отмирания корневища. В постгенеративный период субсенильное возрастное состояние различали по отсутствию генеративных побегов у большинства особей, резкому снижению качества плодоношения и числа почек возобновления, а также партикуляции корневища (распаду материнской особи на дочерние).
Фенологические наблюдения проводили в соответствии с методическими указаниями (16). Отмечали даты наступления и завершения следующих фаз: в цикле вегетативных (розеточных) побегов — отрастания, вегетации, отмирания и покоя; в цикле репродуктивных (генеративных) побегов — начала отрастания, бутонизации, начала цветения, массового цветения, плодоношения, отмирания. Периодичность наблюдений составляла: до фазы бутонизации — 3-4, цветения и плодоношения — 2 раза в неделю. Оценку динамики среднесуточного роста побегов выполняли с интервалом 5-6 сут на 15-20 типичных, случайно выбранных на каждую дату особях. Динамику роста учитывали, исходя из высоты наиболее развитых побегов растения, от уровня почвы до верхушки побега в выпрямленном виде. Ширину листовой пластинки измеряли в самом широком месте, распрямив лист. В развитии побегов отмечали развертывание, взросление, старение и отмирание листовых органов, исходя из характера листосложения, активности функционирования и степени разрушенности листовой пластинки. Для определения биологического возраста в онтогенезе и величины фитомассы в агропопуляции на 1-2-й год жизни с учетных площадок отбирали 18-20 случайно выбранных особей с корневой системой, на 3-4-й год — 8-10; на 6-9-й — 6; на 10-13-й — 5; на 14-17-й — 4. Продуктивность рассчитывали на основе первичного анализа морфологической структуры надземных и подземных органов. Для изучения содержания экдистероидов в различных органах растений, элементах и фракциях фитомассы на 6-8-м году жизни (взрослое генеративное состояние) из агроценоза отбирали по 6-10 типичных особей, надземную часть которых срезали на уровне почвы. В надземной фитомассе выделяли морфологически разнородные органы (17) — укороченные вегетативные (розеточные) и стеблевые генеративные (репродуктивные) побеги; среди розеточных и стеблевых листьев различали разновозрастные фракции (18). В каждой выборке исследовали 275-300 вегетативных побегов, содержащих 1100-1500 розеточных листьев, и 30-35 генеративных побегов со 750-1000 стеблевых листьев. Листовые органы относили к отдельным фракциям, исходя из физиологического возраста и состояния (размеры, опушенность, окраска, цельность листовой пластинки). В составе генеративных побегов идентифицировали: стебли, стеблевые листья разного физиологического возраста (верхние молодые, средние взрослые и нижние старые) и соцветие, вегетативных — фракции молодых, взрослых и старых листьев. У стеблей дополнительно выделяли апикальную (прицветную), верхнюю, среднюю и нижнюю (базипетальную) часть; у соцветий — цветоложе и семена.
Растительный материал (органы, элементы и фракции) сушили при переменной температуре от 23-25 до 35-40 °С и относительной влажности воздуха 25-40 % в соответствии с правилами по заготовке и сушке лекарственного сырья (19). Остаточная влажность воздушно-сухого сырья, определенная методом ускоренной сушки при 130 °С (20), составляла 10-12 %. Образцы из воздушно-сухого сырья для определения концентрации экдистероидов формировали методом квартования. До анализа их хранили в полиэтиленовых пакетах при комнатной температуре 3-5 мес.
Изменчивость содержания экдистероидов в различных элементах лекарственного сырья отслеживали в фазы бутонизации и цветения, в онтогенезе агропопуляции — по возрастным периодам и состояниям, в сезонном цикле развития — со времени весеннего отрастания до фазы покоя осенью. Объектами служили элементы — концентраторы экдистероидов: листовые пластинки вегетативных побегов иапикальные части генеративных побегов, представленные цветоложем и семенами. Образцы вегетативных побегов отбирали в разные фазы развития от 20-25 растений (по 2 розеточных листа), генеративных (семена) — во время фазы плодоношения (от урожая соцветий массового сбора).
Содержание экдистероидов в сухих образцах определяли методом обращенно-фазовой высокоэффективной жидкостной хроматографии (ОФ-ВЭЖХ) с компьютерной обработкой данных по методу внутреннего стандарта (21); анализы выполнены в лаборатории биохимии и биотехнологии растений (1992-2000 годы) и биохимической лаборатории Ботанического сада (2001-2006 годы) Института биологии Коми НЦ УрО РАН (г. Сыктывкар). Использовали жидкостный микроколоночный хроматограф «Милихром-5-3» (колонка 80x2 мм, сорбент Nucleosil C18 с размером частиц 5 мкм) (ООО «Медикант», Россия); элюент — смесь вода:этанол:бутанол (75:24,2:0,8), скорость элюции — 100 мкл/мин, УФ-детектор (l = 242 нм). Учитывали средние значения 2 биологических и 3 аналитических повторностей (% от воздушно-сухого вещества).
Математическую обработку показателей проводили стандартными методами вариационной и корреляционной статистики с помощью модуля Statistica программы Excel 2003. Использовали параметры генеральной (по результатам сплошного учета) и выборочной совокупности. Для исключения систематических погрешностей с краевых участков агроценоза образцы не отбирали, поверхностные слои отбираемых проб не анализировали; данные по единичным экземплярам, по виду отличающимся от нормальных, при обработке не учитывали; к остаточному материалу и типичным особям применяли метод случайной выборки.
Результаты. Р а з в и т и е п о б е г о в в о н т о г е н е з е. В е г е т а т и в н ы е п о б е г и. Продолжительность жизненного цикла агропопуляции R. carthamoides на супесчаных почвах составляет более 17 лет (прегенеративный период — с 1-го по 3-й год жизни). При подзимних сроках посева первые проростки появлялись через 5-7 сут после схода снежного покрова. Основная масса семян всходит в течение 2 нед в I декаде мая. В состоянии проростков особи находились 25-30 сут (средняя масса 2 семядолей — 13,3 мг, что примерно соответствует массе высеянных семян). После перехода в ювенильное возрастное состояние в течение 30-40 сутформировался розеточный побег из 5-7 настоящих листьев, в зоне гипокотиля которого закладывались почки возобновления.
После отмирания первых листьев из их основания начинался рост пазушных почек (их число примерно соответствует числу отмерших за вегетацию листьев). Развития пазушных почек в боковые побеги на 1-м году жизни не происходило из-за короткого вегетационного периода (у единичных растений 1-2 почки успевают дать самостоятельные побеги обогащения).
Фаза кущения у имматурных растений начиналась весной следующего года (в конце апреля—начале мая). При этом зимующая терминальная почка укороченного побега трогалась в рост и формировала новую, вторичную розетку. Через 7-10 сут начиналось развертывание наиболее зрелых пазушных почек, которые образуют побеги 2-го порядка. Процесс развития боковых побегов полностью повторял развитие материнского побега и длился до поздней осени, а из пазух отмирающих листьев начинали расти почки побегов следующего, 3-го порядка. Все пазушные почки прошлогодней закладки (4,1 шт/растение) на 2-м году постепенно достигали стадии зрелости и формировали вегетативные побеги.
Каждый розеточный лист проходил пять фаз развития: выдвижение из почки за счет интеркалярного роста черешков с белесо опушенной, свернутой в трубочку листовой пластинкой; разворачивание листовой пластинки, приобретение светло-зеленой окраски, интенсивный рост; достижение зрелости с прекращением роста и приобретением темно-зеленой окраски; постепенное старение листовой пластинки, приобретающей желто-зеленый оттенок, и засыхание верхушечной части; побурение и отмирание листа с засыханием, скручиванием листовой пластинки к средней жилке и основанию черешков.
Таким образом, в имматурном возрастном состоянии наблюдается ветвление главного розеточного побега на боковые (процесс кущения), развитие которых происходит одновременно с осевым, но со сдвигом календарного срока развертывания из почки. С начала сентября отрастание новых листьев и их рост замедляются, а после начала постоянных заморозков надземные побеги отмирают и все растительные остатки переходят в опад.
Взрослые вегетативные растения (виргинильное возрастное состояние)в начале 3-го года жизни характеризовались наличием 3-порядковой системы побегов. Побеги 1-го зародышевого порядка (у 43 % особей) в третий раз формируют вегетативную розетку. Цикл моноподиального нарастания осей 1-го порядка может повторяться и на 4-5-й сезон вегетации, пока побеги не достигнут генеративного состояния, то есть для них характерна полицикличность. Число органов растений последовательно увеличивалось по годам жизни, составляя соответственно 1,0; 2,8 и 5,7 для вегетативных побегов, 9,6; 12,3 и 23,7 — для розеточных листьев. Высота побегов увеличивается с 21,1 до 58,3 см, надземная фитомасса — с 0,42 до 16,4 г.
C 4-го по 12-й год жизни у большинства особей длится генеративный период (4-5-й годы жизни — молодое, 6-8-й — зрелое, 9-12-й — старое генеративное возрастное состояние). Надземная часть растений при этом представлена сложной системой побегов. По способу нарастания в ней одновременно присутствовали моно-, ди- и полициклические побеги; по функциональной роли — укороченные вегетативные (розеточные) и удлиненные генеративные (стеблевые). По долевому участию преобладают моноподиально нарастающие полициклические побеги, разветвленные на 2-3 порядка и несущие до 20-25 листьев по сравнению с 9-12 у дициклических.
В зоне сближенных междоузлий вегетативных побегов непрерывно закладывались пазушные почки, из которых в последующий год развивались побеги младшего порядка. Число побегов на 4-й год по сравнению с 3-м утроилось, надземная масса — возросла в 3,5 раза (56,8 против 16,4 г). На 5-м году вегетативная сфера состояла из 35,4 побегов и 198,6 розеточных листьев (общая фитомасса — 210,7 г), высота побегов достигала 89,8 см (вегетативные) и 114,0 см (генеративные).
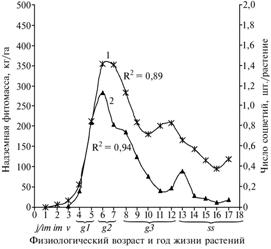 |
Рис. 1. Продуктивность фитомассы и формирова ние развитых репродуктивных органов у растений Rhaponticumcarthamoides в онтогенезе: 1 — сухая масса надземной части, 2 — число соцветий на растении; R2 — соответствующая величина достоверности (Архангельская обл., подзона средней тайги; 1990-2006 годы). Обозначение возраста растений см. в разделе «Методика». |
На 6-7-м году жизни среднестатистические особи до-стигали максимума количественных показателей развития в онтогенезе (рис. 1) (надземная фитомасса — 354,0 г, число побегов — 60,8, при ширине листовой пластинки 28-38 см средняя высота у вегетативных побегов 119,1, генеративных — 143,1 см). Переход от зрелого к старому генеративному возрастному состоянию (8-й год жизни) сопровождался снижением фитомассы до 282,3 г (на 20 %), числа побегов до 52,0 шт. У старых генеративных растений на 9-12-м году наблюдали начало дезинтеграционных процессов: в первичном центре и боковых ветвях корневища образовывались зоны отмерших тканей. Как следствие, возрастало число побегов, отмерших за вегетационный период, и перестало увеличиваться число пазушных почек. Среднее число розеточных листьев на 1 побег в период с 6-го по 9-й год снижалось с 6,5 до 3,9 шт., величина фитомассы — на 41,2 % и составляла 208,7 г.
Сенильный период в целом по агропопуляции с 13-го по 17-й год жизни растения не наблюдали — большинство особей находилось в субсенильном возрастном состоянии. У стареющих растений продолжались дезинтеграционные процессы — общая численность побегов уменьшалась до 24,3-26,0 шт., высота вегетативных побегов — до 71,3-78,6 см, генеративных — до 119,6-120,8 см. Фитомасса составляла 95,1-118,0 г на 1 особь и была близка к показателям молодых генеративных растений.
Г е н е р а т и в н ы е п о б е г и. У небольшой части дициклических побегов терминальная почка ранней весной 2-го года развития трогалась в рост и формировала розетку, а затем репродуктивные органы, развивающиеся в удлиненный ортотропный стебель, который, однако, не достигал фазы плодоношения. Если генеративная часть побега не сформирована с осени в достаточной для инициации цветения степени, в следующем году весь незаконченный цикл повторяется снова до тех пор, пока не сложится благоприятное сочетание условий двух следующих друг за другом вегетационных периодов, позволяющих образовавшемуся полициклическому побегу завершить развитие. Для природных ценозов показано, что растения остаются в прегенеративном возрасте в течение 5-8 лет (3).
В условиях агропопуляции на 3-м году жизни у 17 % особей сформировались зачаточные соцветия, основная часть которых отмирала в начале отрастания, а зацветающие не завязывали семян. Завершить фазу репродукции были способны менее 0,1 % особей агропопуляции. На 4-й год 80 % растений имели в среднем по 1,2 генеративных побега, из которых каждый седьмой достигал фазы плодоношения, на 5-й год жизни число генеративных побегов увеличивалось до 4,2 шт. (плодоносящие — 0,84 шт.). Максимальное число генеративных побегов, тронувшихся в рост, отмечали в начале 6-й вегетации (9,8 шт.), в фазе цветения находились 7,0; число побегов с полным циклом развития до плодоношения равнялось 1,1 шт. В дальнейшем число растений с развитыми соцветиями уменьшалось по годам (см. рис. 1).
Видимый рост генеративных побегов начинался через 5-7 сут после начала отрастания вегетативных побегов из укрупненной флоральной почки, дифференцированной с осени прошлого года на составные элементы (зачаточный стебель со стеблевыми листьями и соцветием). Скорость роста побегов обоих типов уравнивалась примерно с 35-х по 38-е сут вегетации, после чего преимущество в развитии получали генеративные побеги. Прирост побегов обоих типов после 62-68-х сут вегетации полностью прекращался, а с началом плодоношения, сопровождающегося засыханием верхней части стебля, его величина приобретала отрицательные значения. Такая корреляция сохранялась во все годы при значительной разнице в сроках отрастания (17 апреля, 1 и 3 мая) (рис. 2).
|
Рис. 2. Динамика роста побегов у растений Rhaponticum carthamoides в генеративный период (Архангельская обл., подзона средней тайги, 1995-1997, 2000-2002 годы): А, Б и В — соответственно 6-й, 7-й и 8-й год жизни; 1 — генеративные, 2 — вегетативные побеги. |
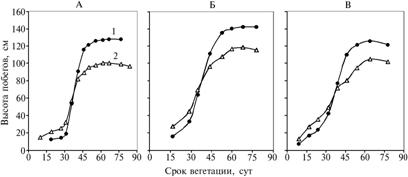
Сроки цветения развитых побегов (независимо от складывающихся погодных условий) приходились на 14-26 июня. В конце июня зацветали менее 1 % побегов. В июле—сентябре появление новых генеративных побегов, их рост и цветение не наблюдали. В целом развитие R. carthamoides до фазы бутонизации занимает 15-23, цветения — 44-56, плодоношения — 72-77 сут. Все побеги после формирования удлиненных генеративных побегов полностью отмирали. Также отмирали почки возобновления, располагающиеся на участке прошлогоднего годового прироста корневища, а в некоторых случаях и боковые вегетативные побеги. Розеточные листья продолжали вегетировать до перехода среднесуточной температуры через 0 °С во II декаде октября с постепенным уменьшением размера и числа.
С о д е р ж а н и е э к д и с т е р о и д о в в о ф р а к ц и я х и э л е м е н т а х ф и т о м а с с ы. В е г е т а т и в н ы е п о б е г и. Как было показано выше, появление новых листьев, их взросление и отмирание не приурочено к определенным фазам развития. Длительность вегетации и масса листьев возрастает до фазы цветения и убывает после плодоношения.
Розеточные листья у взрослых генеративных растений крупные черешковые, более или менее глубоко перисто-рассеченные на 15-22 (0-27) доли; по окраске светло-, темно- или желто-зеленые; образуют розетку диаметром 55-90 (37-112) см. В молодом возрасте размеры у неразвернутых листьев — 3-23 см по длине и 0,3-1,0 см по ширине, у полуразвернутых — 18-68 и 2-8 см, взрослых в фазу цветения — 60-80 (100-120) и 10-25 (35-43) см.
| 1. Содержание фитоэкдистероидов (20Е) во фракциях листьев разного возраста у вегетативных побегов Rhaponticumcarthamoides6-8-го года жизни (Архангельская обл., подзона средней тайги, 1995-1997, 2000-2002 годы, фаза бутонизации) | ||
Розеточные листья (сухая масса одного листа, г) |
Содержание 20Е, % |
|
от сухой массы |
относительное |
|
Молодые: |
|
|
неразвернутые белесые (0,07-0,18) |
0,43 |
100 |
полуразвернутые светло-зеленые (0,35-0,57) |
0,35 |
81,4 |
полуразвернутые темно-зеленые (0,37-1,10) |
0,31 |
72,1 |
Взрослые: |
|
|
темно-зеленые (1,35-2,12) |
0,28 |
65,1 |
зеленые (2,43-2,51) |
0,23 |
53,5 |
светло-желто-зеленые (2,10-2,56) |
0,19 |
44,2 |
Старые: |
|
|
желто-зеленые (1,80-3,76) |
0,16 |
37,2 |
пожелтевшие (1,00-2,11) |
0,11 |
25,6 |
Отмершие: |
|
|
желто-бурые (0,58-1,63) |
0,05 |
11,6 |
почерневшие (0,26-0,69) |
0,02 |
4,6 |
Старые и отмершие листья вегетативных побегов на-блюдали в течение все-го периода вегетации, они сохраняли связь с побегом и составляли фракцию ветоши, переходящей в опад в конце вегетационного сезона. Разновременное развитие листовых зачатков от разных побегов во взрослые органы приводит к одновременному накоплению в надземной сфере фракций отрастающих, молодых растущих, взрослых, старых и отмерших листьев с переходными признаками. При морфологическом анализе структуры фитомассы отдельные фракции розеточных листьев различались по окраске в соответствии с их физиологическим возрастом.
2. Содержание фитоэкдистероидов (20Е, %) в элементах генеративных побегов у растений Rhaponticumcarthamoides 6-го года жизни в разные фазы развития (Архангельская обл., подзона средней тайги, 1995, 2000 годы) |
||
Орган растения |
Фаза развития |
|
бутонизация |
цветение |
|
Соцветие: |
|
|
цветоложе |
0,72 |
1,17 |
семена |
– |
0,57 |
Стебель: |
|
|
апикальная часть |
0,58 |
0,11 |
верхняя часть |
0,25 |
0,03 |
средняя часть |
0,16 |
0,02 |
нижняя часть |
0,10 |
0,01 |
Листья стеблевые: |
|
|
апикальные прицветные |
0,55 |
– |
верхние молодые |
0,28 |
– |
средние взрослые |
0,28 |
0,03 |
нижние старые |
0,28 |
0,02 |
П р и м е ч а н и е. Прочерк означает, что в указанную фазу элемент не формируется. |
||
Листья разных возрастов дифференцированы по содержанию действующих веществ (табл. 1). В фазу бутонизации максимальное содержание 20Е (0,38-0,43 %) в составе вегетативных побегов обнаружили в наиболее молодых по физиологическому возрасту фракциях без выраженной функции фотосинтеза — в неразвернутых белесых листьях с сильной опушенностью. В молодых полуразвернутых листьях содержание фитоэкдистероидов (20Е) снижалось, но незначительно (0,31-0,35 %) при увеличении массы в 3-8 раз.
Во взрослых листьях содержание 20Е было значительно меньше, чем у молодых (см. табл. 1), в старых — еще ниже, в отмерших составляло только 0,05-0,02 %. Масса 1 листа в последних фракциях примерно соответствовала массе неразвернутых и полуразвернутых листьев, поэтому отмеченное уменьшение количества 20Е на порядок может быть связано с оттоком экдистероидов из старых в более молодые развивающиеся органы.
Г е н е р а т и в н ы е п о б е г и. В структуре генеративных побегов различались следующие элементы: стебель (цветонос), стеблевые листья разных ярусов, соцветия. Стебель нарастал за счет вставочного роста междоузлий, на котором спиралеобразно расположены 28-55 стеблевых листьев различной сложности. На верхушке полого неразветвленного стебля формировалось одиночное соцветие — крупная шаровидная корзинка диаметром 4-6 (3-8) см с обоеполыми фиолетово-лиловыми цветками. Развитые цветоносные побеги имели высоту 110-140 (180), диаметр у основания — 1,8-2,0, у верхушки — 0,7-1,2 см.
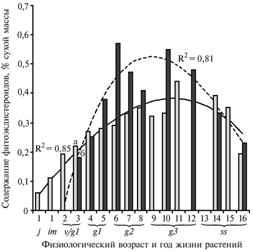 |
Рис. 3. Динамика накопления фитоэкдистероидов в разных органах растений Rhaponticum carthamoides в течение онтогенеза (Архангельская обл., подзона средней тайги, 1990-2005 годы): а — вегетативные побеги (листья розеточные), б — семена. Пунктирная и сплошная линии — сглаженные (приближенные) кривые аппроксимации экспериментальных данных, R2 — соответствующая величина их достоверности. Обозначение возраста растений см. в разделе «Методика». |
Стебель условно разделялся на 4 зоны — нижнюю базипетальную, среднюю, верхнюю и апикальную прицветную. В составе стеблевых листьев также выделялись фракции разного физиологического возраста, а в соцветиях — цветоложе и семена. Листья нижнего яруса физиологически более старые, апикальные прицветные — самые молодые. Стеблевые листья различаются по окраске: апикальные и верхние — темно- и светло-зеле-ные, средние — зеленые или желто-зеленые, нижние — желтые, бурые или почерневшие.
От нижних метамеров к апикальным содержание экдистероидов у растений 6-го года жизни по разным фазам развития варьировало от 0,02 до 0,40, 11-15-го — от 0,01 до 0,58; в том числе во время фазы бутонизации — от 0,10 до 0,58 % (табл. 2). В фазу цветения в нижних, средних и верхних частях побега содержание 20Е снижалось до 0,01-0,03 %, в апикальных (цветоложе и семена) — наоборот, возрастало с 0,28 до 0,57-1,17 %.
Динамика накопления экдистероидов по возрастным периодам онтогенеза R. carthamoides. Во взрослых листьях вегетативных побегов содержание 20Е оказалось минимальным в 1-й год жизни, увеличивалось с возрастом растения и после наступления генеративного периода относительно стабилизировалось (рис. 3).
В ювенильном возрасте этот показатель был равен 0,06-0,11, имматурном — 0,17-0,19 (1-2-й годы жизни), виргинильном — 0,22 % (3-й год), в начале генеративного — 0,27-0,28 (4-5-й годы), во взрослом и старом генеративном (до 12-го года) — 0,32-0,44 %. У субсенильных растений содержание 20Е несколько снизилось и составило 0,39-0,25 % (13-15 год), а на 16-й год достигало значений для виргинильных растений — 0,19 %.
В семенах в онтогенезе наблюдали аналогичную динамику. Максимум содержания экдистероидов в семенах (0,57 %) совпадал с пиком продуктивности в онтогенезе на 6-й год жизни (см. рис. 1).
У зрелых генеративных растений на 6-8-й год массовое плодоношение и последующее снижение продуктивности сочеталось с одновременным уменьшением содержания 20Е в семенах (с 0,57 до 0,47-0,41 %) (см. рис. 3). На 9-10-й год после перерыва в плодоношении у значительной части особей, содержание экдистероидов достигало исходного уровня (0,55 % на 10-й год), что свидетельствует о напряженности донорно-акцепторных отношений между вегетативными и генеративными побегами у старовозрастных растений. Еще более значительное (7-кратное) подавление репродукционных процессов на 11-й год приводило к возрастанию содержания экдистероидов в вегетативных побегах с 0,32-0,33 до 0,44 %.
Имелась сильная положительная зависимость между содержанием экдистероидов в семенах и интенсивностью репродукционных процессов у молодых и взрослых генеративных растений (r = 0,99-0,96 при p = 0,999). В свою очередь, формирование генеративных побегов и плодоношение зависят от продуктивности фитомассы (см. рис. 1). В вегетативных побегах содержание экдистероидов коррелировало с продуктивностью надземной массы (r = 0,78-0,81, p = 0,99). Отсутствие статистически достоверной зависимости содержания экдистероидов в листьях от семенной продуктивности связано с незначительной долей генеративных побегов в структуре фитомассы.
Д и н а м и к а с о д е р ж а н и я ф и т о э к д и с т е р о и д о в з а в е г е т а ц и о н н ы й п е р и о д. Известна изменчивость абсолютной величины содержания экдистероидов в растительных тканях в течение вегетационного сезона (22, 23). Розеточные листья R. carthamoides вегетируют от ранней весны до поздней осени, поэтому выявление пределов такой изменчивости во фракциях листьев практически значимо.
|
Рис. 4. Динамика содержания фитоэкдистероидов в листьях взрослогенеративных растений Rhaponticum carthamoides во время прохождения вегетационного периода (Архангельская обл., подзона средней тайги, 1990-2005 годы): 1, 3 и 4 — соответственно молодые, взрослые и старые растения, 2 — влажность почвы. Сглаженные (приближенные) кривые аппроксимации экспериментальных данных, R2 — соответствующая величина их достоверности. |
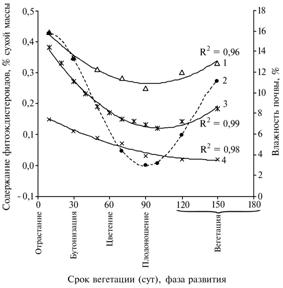
В молодых листьях розеточных полициклических побегов (неразвернутых и полуразвернутых) в начале фазы отрастания содержание 20Едостигало 0,45-0,55 %, через 20 сут вегетации — составляло 0,35-0,40 % (рис. 4). В этот период влажность корнеобитаемого слоя почвы колебалась в пределах 15-18 %, что соответствует оптимальным условиям роста и развития вида (24). В июле—августе этот показатель снижался до 0,25-0,28 %, что совпадало с наступлением засушливых условий (влажность почвы — около 3,0 %). В сентябре после повышения влажности почвы до 11-14 % содержание 20Е вновь возрастало. В целом пределы колебания показателяу молодых листьев равнялись 0,28-0,55 %.
Во взрослых листьях падение содержания 20Е также приходилось на засушливый период, а разница показателя за период вегетации была 3-кратной — 0,12-0,38 %. В старых листьях наблюдалось практически равномерное снижение содержания 20Е, которое не зависело от фазы или календарных сроков развития растений (от 0,15 до 0,02 %, то есть в 7-8 раз).
Итак, на европейском Севере жизненный цикллевзеи сафлоровидной (Rhaponticum carthamoides) в условиях агропопуляций длится свыше 17 лет. Потенциал продуктивного долголетия закладывается в прегенеративном возрасте и базируется на системе вегетативных полициклических побегов, разветвленных на 2-3 порядка и включающих 20-25 розеточных листьев. Формирование и развитие репродуктивных органов вторично по отношению к вегетативным. Наивысших показателей развития в онтогенезе среднестатистические особи достигают на 6-7-й год жизни. Вегетативные побеги функционируют в течение всего вегетационного периода, генеративные после формирования репродуктивных органов отмирают, часто не достигая фазы плодоношения. Появление новых розеточных листьев, их рост и отмирание не приурочены к определенным фазам развития. Разновременное развитие листовых зачатков от разных побегов во взрослые органы приводит к одновременному накоплению в надземной фитомассе фракций отрастающих, молодых растущих, взрослых, старых и отмерших листьев. Накопление действующих веществ (экдистероидов) в онтогенезе положительно коррелирует с величиной продуктивности надземных органов. Динамика содержания экдистероидов в отдельных элементах и фракциях фитомассы согласуется с процессом перераспределения биологически активных веществ от старых органов к растущим и развивающимся: максимальные показатели выявлены в неразвернутых белесых, паутинисто-опушенных листьях, цветоложе и семенах. Падение содержания экдистероидов до минимальных значений совпадает с наступлением засушливых условий в летний период (июль—август). Относительное восстановление показателей происходит в начале осени в молодых и взрослых листьях при снижении дневной температуры и возрастании относительной влажности почвы. В старых листьях снижение постоянно (от 0,15 до 0,02 %) и не зависит от календарных сроков вегетации.
Л И Т Е Р А Т У Р А
1. Государственная фармакопея СССР. Издание IX, X, XI (в 2-х т.). М., 1961; 1968; 1987; 1990.2. Государственный реестр лекарственных средств. (Лекарственные растения и сырье). М., 1995; 1998.
3. П о с т н и к о в Б.А. Маралий корень и основы введения его в культуру. Новосибирск, 1995.
4. Т и м о ф е е в Н.П. Промышленные источники получения экдистероидов. Часть II. Ecdysterone: Растения рода Rhaponticum (обзор). В сб. науч. тр. Нетрадиционные природные ресурсы, инновационные технологии и продукты. М., 2007, 14: 8-49.
5. Растительные ресурсы СССР. Цветковые растения, их химический состав, использование (сем. Asteraceae). СПб, 1993, 7: 161-163.
6. Корневище с корнями рапонтикума сафлоровидного (левзеи сафлоровидной). Фармакопейная статья ФС 42-2707-90. М., 1990.
7. S t e c h J., C m o l i k o v a R., O p l e t a l L. e.a. Zpusob pripravy extraktu z oddenku s koreny parchy saflorove. Prague, Patent Czechia 279814, July 12, 1995.
8. Т и м о ф е е в Н.П., Л а п и н А.А., З е л е н к о в В.Н. Оценка качества лекарственного сырья левзеи сафлоровидной методом бромной антиокислительной емкости. Бутлеровские сообщения, 2006, 8(2): 35-40.
9. D i n a n L., S a v c h e n k o T., W h i t i n g P. On the distribution of phytoecdysteroids in plants. Cel. Mol. Life Sci., 2001, 58(8): 1121-1132.
10. Фитоэкдистероиды /Под ред. В.В. Володина. СПб, 2003.
11. Ш а и н С.С. Биорегуляция продуктивности растений. М., 2005.
12. Природно-климатический очерк Котласского района Архангельской области. М., 1994.
13. Научно-прикладной справочник по агроклиматическим ресурсам СССР (ежегодные данные). Архангельская область. Архангельск, 1992-1996.
14. Р а б о т н о в Т.А. Фитоценология. М., 1983.
15. З а у г о л ь н о в а Л.Б., Ж у к о в а А.А., К о м а р о в А.С. и др. Ценопопуляции растений (очерки популяционной биологии). М., 1988.
16. Методические указания по изучению коллекции многолетних кормовых трав /Под ред. А.И. Иванова. М., 1979.
17. Методические указания по селекции многолетних трав: фенологические наблюдения. Оценка селекционного материала. М., 1985: 90-103.
18. С е м е н о в а-Т я н-Ш а н с к а я А.М. Накопление и роль подстилки в травяных сообществах. Л., 1977.
19. Справочник по заготовкам лекарственных растений /Под ред. Д.С. Ивашина, З.Ф. Катиной, И.З. Рыбачук и др. Киев, 1989.
20. Р а з у м о в В.А. Справочник лаборанта-химика по анализу кормов. М., 1996.
21. П у н е г о в В.В., С а в и н о в с к а я Н.С. Метод внутреннего стандарта для определения экдистероидов в растительном сырье и лекарственных формах с помощью ВЭЖХ. Растительные ресурсы, 2001, 37(1): 97-102.
22. А л е к с е е в а Л.И., Т е т е р ю к Л.В., В о л о д и н В.В. Динамика содержания экдистероидов у Ajuga reptans L. на северной границе ее ареала. 23. Z i b a r e v a L. Distribution and levels of phytoecdysteroids in plants of the genus Silene during development. Arch. Insect. Biochem. Physiol., 2000, 43(1): 1-8.
24. Т и м о ф е е в Н.П. Возраст и динамика плотности Rhaponticumcarthamoides (Willd.) Iljin и Serratula coronata L. (Asteraceae) на Европейском Севере. Растительные ресурсы, 2005, 41(3): 1-13.
ACCUMULATION AND VARIABILITY OF THE ECDYSTEROIDS CONTENT IN MEDICINAL RAW MATERIAL OF Rhaponticum carthamoides
N.P. Timofeev
In the conditions of European North and during 17-years life cycle of agropopulation the author studied the connatural laws, influencing on productive longevity, accumulation of ecdyster-oids and variability of quality of medicinal raw material of Rhaponticum carthamoides. It was established, that the potential of productive longevity of the species is based on system of vegetative polycyclic shoots. Accumulation of effective substances (ecdysteroids) in an ontogenesis positively corre-lates with productivity of above-ground organs with the minimum registrated under drought condi-tions. Dynamics of the contents ecdysteroids in separate fractions and elements phytomass is combined with process of formation of propagules at which there is redistribution from old in favour of growing and developing organs.
Key words: Rhaponticum carthamoides, development in ontogenesis, productivity, medicinal raw material, contents ecdysteroids, 20-hydroxyecdysone.
Научно-производственное предприятие «КХ БИО», |
Поступила в редакцию |
Оформление электронного оттиска
![]()
![]()




