








УДК 634.74:581.3
ДИНАМИКА ПРОРАСТАНИЯ ПЫЛЬЦЫ И РОСТА ПЫЛЬЦЕВЫХ ТРУБОК in vitro У ГИБРИДОВ ОБЛЕПИХИ КРУШИНОВИДНОЙ РАЗНОГО ЭКОЛОГО-ГЕОГРАФИЧЕСКОГО ПРОИСХОЖДЕНИЯ
Д.А. ЛАПШИН, Т.Н. КУЗНЕЦОВА
Используя питательную среду, изучали качественные показатели пыльцы у гибридов Hippophaё rhamnoides L. разного эколого-географического происхождения селекции Нижегородской государственной сельскохозяйственной академии. Уточнена методика определения жизнеспособности пыльцы и установлено время прорастания всей жизнеспособной пыльцы. Показано, что гибриды H. rhamnoides по динамике прорастания пыльцы и роста пыльцевых трубок подразделяются на две группы с разной продолжительностью этих процессов (соответственно 2-4 и 16-22 ч).
Ключевые слова: облепиха крушиновидная, гибриды, пыльцевое зерно, пыльцевая трубка, жизнеспособность.
Облепиха крушиновидная (Hippopha rhamnoides L.) (1) в отличие от большинства плодово-ягодных культур — двудомное растение, поэтому в формировании высокого урожая плодов у H. rhamnoides важная роль принадлежит мужским особям (2, 3). Их селекция велась по хозяйственно ценным признакам — зимостойкости, габитусу, вилтоустойчивости (4-6), однако в последнее время уделяется внимание пыльцепродуктивности и качественным показателям пыльцы, представляющим как теоретический, так и практический интерес. Вопросы качества пыльцевых зерен (ПЗ) H. rhamnoides освещены сравнительно мало (7-9). Основными показателями качества ПЗ служат прорастаемость (жизнеспособность) и длина пыльцевой трубки (ПТ) как фактор, характеризующий количество собственных питательных веществ ПЗ. В доступной нам литературе данных по динамике качественных показателей ПЗ гибридов H. rhamnoides мы не обнаружили.
В связи с этим целью нашей работы была характеристика гибридов облепихи кушиновидной разного эколого-географического происхождения селекции Нижегородской государственной сельскохозяйственной академии (НГСХА) по динамике прорастания ПЗ и росту ПТ in vitro.
Методика. Исследования проводили в течение весенних периодов 2005-2007 годов. Объектом изучения служили мужские гаметофиты (пыльца) гибридов H. rhamnoides разного эколого-географического происхождения селекции В.А. Фефелова (НГСХА): Геракл, 31/89 и 5/93 — гибриды, полученные от опыления сортов Ботанического сада Московского государственного университета им. М.В. Ломоносова (далее сорта МГУ, прибалтийский экотип) пыльцой мутантной мужской формы катунской облепихи; Дебют — гибрид катунского экотипа; 5/87, 1/89 и 1/91 — гибриды, полученные от опыления сортов МГУ опылителями катунского экотипа; 1/90 — гибрид, полученный от опыления сортов МГУ опылителями саянского экотипа. По комплексу морфофизиологических признаков все изученные мужские гибриды можно условно разделить на две группы: с преобладанием морфофизиологических признаков прибалтийского (Геракл, 31/89, 1/90 и 5/93) и катунского (Дебют, 5/87, 1/89 и 1/91) экотипа.
Зрелую пыльцу отбирали по описанной методике (10). Жизнеспособность пыльцы in vitro оценивали методом Транковского (11, 12) (3 повторности по 1000 ПЗ в каждой; проращивание в чашках Петри при температуре 20-22 °C). Динамику прорастания ПЗ прослеживали темпорально — через 2, 4, 8, 12, 16, 22 и 24 ч после посева на питательную среду. При просмотре препаратов проросшими считали ПЗ, имевшие длину ПТ более диаметра ПЗ, наклюнувшимися — с длиной ПТ не менее 1/2 диаметра ПЗ. Длину ПТ измеряли у 100 проросших ПЗ каждого образца через 2, 4, 8, 12, 16, 22 и 24 ч, используя микрометр МОВ-1-15 (Россия) при увеличении x375. Все препараты просматривали в световом микроскопе Laboval-4 («Carl Zeiss», Германия).
Данные обработаны методами прикладной статистики (13, 14) с использованием пакета программ Microsoft Excel и Statistica 6.0. Жизнеспособность и динамику прорастания ПЗ оценивали при помощи критерия Пирсона (χ2), длину ПТ — критерия Крускала-Уоллиса, представляющего собой непараметрический аналог показателя, который дает однофакторный дисперсионный анализ, а также множественного попарного сравнения по критерию Крускала-Уоллиса с поправкой Бонферрони (14). Для выявления сходства между гибридами применяли кластерный анализ (мера удаленности — Eвклидово расстояние, иерархическое объединение в кластеры по методу Уорда) (14).
Результаты. Известно, что трудности при использовании метода проращивания in vitro для определения жизнеспособности ПЗ и длины ПТ у растений любых видов обусловлены необходимостью подбора условий культивирования. Мы оптимизировали состав среды, предложенной другими авторами для проращивания ПЗ облепихи (15). Разработанная модификация представляет собой 1 % агаризованную среду Транковского с содержанием сахарозы 15 % при добавлении CaCl2 и H3BО3 (по 3x10-5 %).
У всех изученных гибридов единичные проросшие ПЗ отмечались уже через 1 ч после посева свежесобранной пыльцы на модифицированную среду Транковского; время прорастания всех потенциально жизнеспособных ПЗ составляло 16-22 ч (рис. 1, А). ПТ H. rhamnoides начинали лизироваться через 22 ч после посева ПЗ (см. рис. 1, Б), что соответствует периоду прорастания всех потенциально жизнеспособных ПЗ. Поэтому оценку жизнеспособности пыльцы и длины ПТ H. rhamnoides целесообразно проводить не ранее, чем через 22 ч после посева на питательную среду.
Анализ данных по жизнеспособности ПЗ с использованием критерия χ2 выявил статистически значимые различия как по годам наблюдений (χ2 = 304,15; p < 0,001), так и между изученными гибридами на пыльце, полученной в один год (χ2 = 65063,90; p < 0,001).
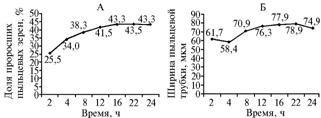 |
Рис. 1. Динамика прорастания пыльцы (А) и роста пыльцевых трубок (Б) у гибридов Hippopha rhamnoides при высеве на модифицированную среду Транковского (усредненные данные за 2005-2007 годы исследований по всем изученным гибридам; описание гибридов см. в разделе «Методика»). |
В 2005 году более активно прорастали ПЗ у гибридов Дебют, 1/91 и 5/93 (жизнеспособность соответственно 61,0; 69,7 и 66,4 %), в 2006 — у гибридов 1/89 и 31/89 (50,5 и 53,5 %), в 2007 — у гибрида 5/87 (37,7 %).
Наименьший показатель прорастаемости пыльцы каждый год отмечали у разных образцов: в 2005 — у гибрида 31/89 (6,8 %), в 2006 — у гибрида Дебют (21,1 %), в 2007 — у гибрида Геракл (11,0 %); в том же году у гибридов 1/89 и 1/91 пыльца имела прорастаемость менее 1 %, то есть была охарактеризована как нежизнеспособная.
В 2005 году гибриды сформировали пыльцу, наиболее разнокачественную по жизнеспособности. Так, у растений 1/91 этот показатель составлял 69,7 %, у 31/89 — только 6,8 %, что стало наименьшим значением для всех образцов за три года исследований. Отмеченный максимум можно объяснить благоприятными погодными условиями в зимне-весенний период формирования пыльцы.
|
Рис. 2. Жизнеспособность пыльцы (А) и динамика роста пыльцевых трубок (Б) у гибридов Hippopha rhamnoides разного эколого-географического происхождения по годам наблюдения при высеве на модифицированную среду Транковского: 1 — Геракл, 2 — 31/89, 3 — 5/93, 4 — Дебют, 5 — 5/87, 6 — 1/89, 7 — 1/91, 8 — 1/90 (описание гибридов см. в разделе «Методика»). |
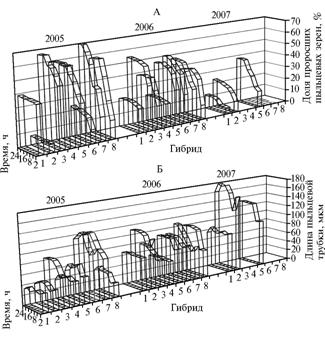
Снижение доли прорастающих ПЗ в 2007 году зарегистрировали практически у всех гибридов, кроме 31/89, при минимальном разбросе показателей жизнеспособности между гибридами (от 11,0 до 37,7 % соответственно у гибридов Геракл и 5/87) (рис. 2). Резкое снижение жизнеспособности пыльцы, вероятно, стало следствием заморозков в период развития мужского гаметофита у растений. Действительно, в 2005 и 2007 годах, когда заморозки были отмечены соответственно 6 и 20 раз, жизнеспособность пыльцы варьировала от наибольшей до резко снизившейся у части образцов (вплоть до гибели пыльцы у гибридов 1/89 и 1/91).
Выявленные различия в динамике прорастания ПЗ гибридов H. rhamnoides (см. рис. 2) позволили разделить их на 2 группы. В 1-й (гибриды Геракл и Дебют) для пыльцы было характерно дружное прорастание через 2-4 ч после посева, во 2-й — несинхронизированное, продолжительное по времени прорастание (в течение 22 ч после посева).
При кластерном анализе изученные гибриды по степени сходства динамики (характера) прорастания также разделились на 2 группы: в 1-ю вошли образцы 1/91, 1/90, 5/93, 31/89, 1/89 и 5/87, во 2-ю — Дебют и Геракл с формированием подгруппы внутри каждой группы. Наиболее сходными были следующие пары, имеющие как одинаковое, так и разное эколого-географическое происхождение: 1/91 и 1/90; 5/93 и 31/89; 1/89 и 5/87; Дебют и Геракл (рис. 3).
Поскольку при анализе значений длины ПТ по критерию согласия Колмогорова-Смирнова (d = 0,15; p < 0,001) и Лиллиефорса (l = 0,15; p = 0,001) обнаруживались достоверные отклонения от нормального распределения, для оценки показателя применяли методы непараметрической статистики. Используя критерий Крускала-Уоллиса, выявили статистически значимые различия в разные годы наблюдений (χ2 = 225,16; p < 0,001), поэтому далее выполняли межгибридный анализ по годам исследований. Статистически значимые различия между гибридами по длине ПТ отмечали во все годы (2005: χ2 = 242,89; p < 0,001; 2006: χ2 = 166,13; p < 0,001; 2007: χ2 = 86,35; p < 0,001). Выявленное достоверное влияние генотипических особенностей на проявление признака указывало на необходимость проведения попарных межгибридных сравнений. Такое сравнение по критерию Крускала-Уоллиса с поправкой Бонферрони по годам (табл.) показало, что статистически значимые различия отсутствуют только между следующими гибридами: Геракл и 1/90; 31/89 и 5/87, 1/89, 1/91; 5/93 и 1/90; 5/87 и 1/89; 1/89 и 1/91. Гибриды, которые между собой статистически значимо не различались по длине ПТ (см. табл.), неодинаково реагировали на изменение внешних условий по годам наблюдений (см. рис. 2). В других парных комбинациях выявлены достоверные различия гибридов, следовательно, более объективную оценку дает анализ показателей у отдельных гибридов разного эколого-географического происхождения по годам. Полученные результаты указывают на целесообразность изучения динамики роста ПТ в качестве критерия качества мужских гибридов.
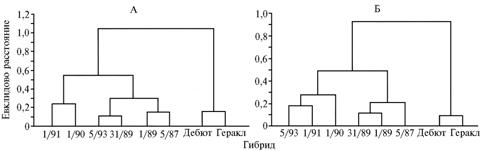 |
Рис. 3. Дендрограмма сходства гибридов Hippopha rhamnoides разного эколого-географического происхождения по динамике прорастания пыльцы (А) и роста пыльцевых трубок (Б). Описание гибридов см. в разделе «Методика». |
В 2005 году наиболее длинные ПТ имела пыльца гибридов 1/89 (119,6 мкм) и 1/91 (123,9 мкм); в 2006 — 31/89 (111,3 мкм), 1/90 (100,4 мкм) и 1/91 (105,6 мкм); в 2007 — гибрида 31/89 (179,7 мкм) (см. рис. 2).
В целом, наибольшую длину ПТ отмечали в 2007 году, что, на наш взгляд, связано с уменьшением общего количества пыльцы вследствие повреждения мужских растений из-за аномально сложившихся погодных условий в осенне-зимний и зимне-весенний периоды 2006-2007 годов. Однако только для образцов пыльцы гибридов 5/87 и 31/89 достоверно показано, что, несмотря на снижение числа проросших ПЗ, они имели более длинные ПТ, чем в предыдущие годы. Самыми короткими ПТ в 2005 и 2007 годах характеризовалась пыльца гибрида Геракл, в 2006 — гибрида Дебют (см. рис. 2).
За три года исследований наибольшую и наименьшую вариабельность по этому признаку выявили в 2006 году соответственно у гибридов Дебют (Cv= 81,9 %) и 1/89 (Cv = 26,8 %).
Таким образом, относительно стабильной длиной ПТ во все годы исследований характеризовался гибрид Дебют (48,6 мкм — в 2005, 49,6 мкм — в 2006 году) (см. рис. 2). У гибрида 31/89 длина ПТ значительно варьировала (см. рис. 2): в 2005 году — составила 44,0, в 2007 — 179,7 мкм; аналогичную, но менее выраженную тенденцию наблюдали у гибрида 5/87.
Следует отметить, что как максимальные, так и минимальные значения длины ПТ и коэффициентов вариации зарегистрировали в разные годы исследований у разных гибридов.
| Показатели попарного сравнения гибридов Hippopha rhamnoides разного эколого-географического происхождения по годам (по критерию Крускала-Уоллиса с учетом поправки Бонферрони) | ||||||||||
Гибрид |
Гибрид |
|||||||||
Геракл |
31/89 |
5/93 |
Дебют |
5/87 |
1/89 |
1/91 |
1/90 |
|||
2005 г о д |
||||||||||
Геракл |
1 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
0,0095 |
||
31/89 |
< 0,0010 |
1 |
< 0,0010 |
< 0,0010 |
0,2109 |
0,8807 |
0,0631 |
< 0,0010 |
||
5/93 |
< 0,0010 |
< 0,0010 |
1 |
< 0,0010 |
0,0016 |
< 0,0010 |
< 0,0010 |
0,0822 |
||
Дебют |
< 0,0010 |
< 0,0010 |
< 0,0010 |
1 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
||
5/87 |
< 0,0010 |
0,2109 |
0,0016 |
< 0,0010 |
1 |
0,1230 |
< 0,0010 |
< 0,0010 |
||
1/89 |
< 0,0010 |
0,8807 |
< 0,0010 |
< 0,0010 |
0,1230 |
1 |
0,0120 |
< 0,0010 |
||
1/91 |
< 0,0010 |
0,0631 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
0,0120 |
1 |
< 0,0010 |
||
1/90 |
0,0095 |
< 0,0010 |
0,0822 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
1 |
||
2006 г о д |
||||||||||
Геракл |
1 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
0,0095 |
||
31/89 |
< 0,0010 |
1 |
< 0,0010 |
< 0,0010 |
0,2109 |
0,8807 |
0,06310 |
< 0,0010 |
||
5/93 |
< 0,0010 |
< 0,0010 |
1 |
< 0,0010 |
0,0015 |
< 0,0010 |
< 0,0010 |
0,0822 |
||
Дебют |
< 0,0010 |
< 0,0010 |
< 0,0010 |
1 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
||
5/87 |
< 0,0010 |
0,2109 |
0,0015 |
< 0,0010 |
1 |
0,1231 |
< 0,0010 |
< 0,0010 |
||
1/89 |
< 0,0010 |
0,8807 |
< 0,0010 |
< 0,0010 |
0,1231 |
1 |
0,0123 |
< 0,0010 |
||
1/91 |
< 0,0010 |
0,0631 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
0,0123 |
1 |
< 0,0010 |
||
1/90 |
0,0095 |
< 0,0010 |
0,0822 |
< 0,0010 |
< 0,0010 |
< 0,0010 |
|
1 |
||
2007 г о д |
||||||||||
Геракл |
1 |
< 0,0010 |
|
|
< 0,001 |
|
|
|
||
31/89 |
< 0,0010 |
1 |
|
|
< 0,001 |
|
|
|
||
5/87 |
< 0,0010 |
< 0,0010 |
|
|
1 |
|
|
|
||
П р и м е ч а н и е. В таблице представлены только p-значения. Описание гибридов см. в разделе «Методика». В 2005-2006 годах достоверными считали различия при уровне значимости p < 0,0018, в 2007 — при p < 0,0167. |
||||||||||
Анализ динамики роста ПТ (см. рис. 2) позволил разделить гибриды на 2 группы: 1-я — с дружным и интенсивным ростом ПТ в первые 2 ч и последующим резким замедлением (гибриды Геракл и Дебют); 2-я — с несинхронизированным, продолжительным периодом роста (формы 31/89, 5/93, 5/87, 1/89, 1/91 и 1/90). По результатам кластерного анализа динамики роста ПТ изученные гибриды также разделились на 2 группы (и далее на подгруппы) (см. рис. 3): в 1-ю вошли гибриды 5/93, 1/91, 1/90, 31/89, 1/89 и 5/87, во 2-ю — Дебют и Геракл. Наиболее сходными по этому показателю были пары гибридов 5/93 и 1/91, 31/89 и 1/89, Дебют и Геракл.
Следует обратить внимание, что как по динамике прорастания ПЗ, так и по динамике роста ПТ (см. рис. 2) в одну группу вошли гибриды разного эколого-географического происхождения. Этот факт позволяет утверждать, что при работе со сложными гибридами их разделение на группы по подобному принципу и дальнейший анализ таких групп малоэффективны. Требуется индивидуальная и детальная оценка каждого гибрида, хотя, несомненно, между гибридами из одной группы имеется определенное сходство.
Несмотря на гибель части мужских гибридов H. rhamnoides из-за критических погодных условий зимы 2007 года, резкого угнетения роста ПТ полученных ПЗ мы не отмечали. На наш взгляд, доля проросших ПЗ отражает потенциальную фертильность пыльцы, а длину ПТ, которая характеризует количество запасных питательных веществ в ПЗ, обеспечивающих рост ПТ, можно рассматривать как более объективный показатель ее фертильности.
Итак, пыльцевые зерна гибридов Hippopha rhamnoides на модифицированной среде Транковского при 20-22 °С начинают прорастать через 1 ч после посева, время прорастания всех потенциально жизнеспособных пыльцевых зерен H. rhamnoides составляет не менее 22 ч. По динамике прорастания пыльцы и роста пыльцевых трубок изученные гибриды H. rhamnoides можно разделить на 2 группы. К 1-й относятся образцы, у которых эти процессы в основном завершаются через 2-4 ч после посева (Геракл и Дебют), ко 2-й — те, у которых они происходят менее синхронно и в течение более длительного периода — 16-22 ч (5/87, 1/89, 31/89, 1/90, 1/91 и 5/93).
Л И Т Е Р А Т У Р А
1. Т а х т а д ж я н А.Л. Систематика и филогения цветковых растений. М.-Л., 1966.2. Ж а м с р а н Ц. Биология облепихи крушиновидной Северной Монголии. Автореф. канд. дис. Иркутск, 1971.
3. Б у к ш т ы н о в А.Д., Т р о ф и м о в Т.Т., Е р м а к о в Б.С. и др. Облепиха. М., 1985.
4. К о н д р а ш о в В.Т., С о к о л о в а Е.П. О новых вилтоустойчивых формах облепихи. Бюл. Моск. об-ва испытателей природы, Отд. Биол. М., 1991, 96(I): 146-153.
5. Ф е ф е л о в В.А., С е л е х о в В.В., К о р о в и н а М.А. и др. Материалы к разработке программы селекции мужских сортов-опылителей облепихи. В сб.: Мат. III. Междунар. симп. по облепихе. Новосибирск, 1998: 32-35.
6. С е л е х о в В.В. Селекционно-биологическая оценка мужских форм облепихи: Автореф. кад. дис. Брянск, 2000.
7. К о н д о р с к а я В.Р. Морфология тычиночного цветка и развитие мужского гаметофита облепихи. Науч. докл. высшей школы. Биол. науки. М., 1967, 4: 69-75.
8. И л ь и н а Н.А. Особенности опыления облепихи в Челябинской области. Бюл. Всес. ин-та растениеводства им. Н.И. Вавилова. Л., 1982, вып. 118: 70-71.
9. К а м е л и н а О.П., П р о с к у р и н а О.Б. Семейство Elaeagnaceae. В кн.:Сравнительная эмбриология цветковых растений. Davidiaceae—Asteraceae. Л., 1987: 105-110.
10. С о к о л о в а Е.П., К о н д р а ш о в В.Т. Биологические особенности опылителей облепихи, перспективных для Центральной Черноземной зоны. В сб. науч. тр. Горьковского с.-х. ин-та: Биология, селекция и агротехника облепихи. Горький, 1988: 64-70.
11. П а у ш е в а З.П. Практикум по цитологии растений. М., 1974.
12. Б а р ы к и н а Р.П., В е с е л о в а Т.Д., Д е в я т о в А.Г. и др. Справочник по ботанической микротехнике. Основы и методы. М., 2004.
13. Д о с п е х о в Б.А. Методика полевого опыта. М., 1973.
14. Г л а н ц С. Медико-биологическая статистика. Пер. с англ. М., 1998.
15. Методические рекомендации по применению цитологических методов в плодоводстве. М., 1988.
DYNAMICS OF POLLEN GERMINATION AND GROWTH OF POLLEN TUBES in vitro IN Hippopha rhamnoides L. HYBRIDS OF DIFFERENT ECOLOGO-GEOGRAPHIC ORIGIN
D.A. Lapshin, T.N. Kuznetsova
By in vitro cultivation the qualitative parameters of pollen were investigated in hybrids of Hippopha rhamnoides L. of different ecologo-geographic origin of selection of Nizhni Novgorod State Agricultural Academy. The technique for determination of pollen viability was perfected and the time of germination all viable pollen was determined. It was established that all H. rhamnoides hybrids on dynamics of pollen germination and growth of pollen tubes subdivide into two groups with different duration of these processes (2-4 and 16-22 hours, respectively).
Key words: sea buckthorn, hybrids, pollen-grain, pollen-tube, viability.
ФГОУ ВПО Нижегородская государственная |
Поступила в редакцию |
Оформление электронного оттиска
![]()
![]()




